
倒披针叶虫实种内功能性状的变异特征及对种群稳定的影响
2017-09-07 10:06:55汤景光刘彤周娟崔向阳
石河子大学学报(自然科学版) 2017年4期
汤景光,刘彤*,周娟,崔向阳
(1石河子大学生命科学学院,新疆 石河子 832003;2新疆生产建设兵团第十四师224团,新疆 和田 848116)
倒披针叶虫实种内功能性状的变异特征及对种群稳定的影响
汤景光1,刘彤1*,周娟1,崔向阳2
(1石河子大学生命科学学院,新疆 石河子 832003;2新疆生产建设兵团第十四师224团,新疆 和田 848116)
倒披针叶虫实 (Corispermum lehmannianum)在古尔班通古特沙漠呈优势分布,其重要值仅次于梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)。本研究在古尔班通古特沙漠选择标准样地,通过不同坡位的定点观测,从种内功能性状变异与种群稳定关系的角度,分析倒披针叶虫实种群维持机理。结果表明:(1)该物种种群内存在广泛的性状变异:在时间尺度上,株高、冠幅变异系数随生长发育先增加后降低;在空间尺度上,在同一坡位灌木树冠覆盖区域变异显著高于裸地区域(P<0.05);不同坡位种群对比发现,除萌发期外,坡顶种群性状及变异与其他坡位种群均存在显著差异(P<0.05)。(2)种内变异受不同坡位环境强烈的影响,坡位对种群密度与萌发现象的影响显著:坡顶种群密度最小,萌发数量最少;坡中密度最大,萌发数量最多;萌发率与死亡率均随时间变化降低。综合研究表明,在干旱荒漠区,倒披针叶虫实种群稳定主要受不同坡位、不同树冠覆盖度等微生境的强烈影响,种内功能性状变异表现出对荒漠环境的良好的适应性,对种群维持也起着重要作用。
种内变异;干旱区;功能性状;种群稳定
种群是物种存在的基本单位,又是生态功能的重要单位,近年来众多研究表明种内变异与环境对种群的稳定起关键作用[1-4]。
同一种群的不同个体间存在着丰富的形态[5-6],物候[7],行为及资源利用[8]方式等方面的差异。自然种群内个体表型的变异会影响种群维持[9],种内相互作用强度随着种内性状变异的程度的增大而减小[10]。种群内个体的物候及个体发育差异同样有利于种群的稳定性的维持[11],种内个体物候差异提高了物种在环境变化下的适应能力[12],种内个体发育的异时性降低了种群内个体同时遭遇病菌、天敌等危害的风险[13],同时不同发育期的个体也是维持种群稳定的温备用[14]。种群内个体行为的差异能够影响种内和种间竞争及捕食强度进而决定物种延续[15]。种内功能变异同样能够缓解环境对种群的胁迫甚至能够缓冲物种灭绝的压力[16]。
环境因素在种内性状变异维持种群动态及稳定等生态过程中起着至关重要的作用[13,17],环境过滤作用通过直接作用于功能性状影响种群,在区域尺度环境过滤效应倾向于具有选择相似功能的性状的群体构成功能群[18],而在局部尺度,不同微生境可增加种内性状差异,进而提高了种群稳定性[19]。种内性状变异的相关研究主要集中在森林及草原生态系统[20-21],然而在干旱荒漠区种内性状变异和环境对种群稳定贡献大小和差异研究较少。尤其在干旱荒漠区,降水量少且波动大[22],环境过滤强,因此沙漠植物种群如何在干旱且环境条件波动剧烈的情况下维持种群稳定的机制研究就尤为重要。
倒披针叶虫实属藜科(Chenopodiaceace)虫实属(Corispermum)短命植物,在古尔班通古特沙漠分布重要值仅次于梭梭和白梭梭,其生长季是风沙活动强盛期[23-24],生态功能位居草本植物之首[23],所以该物种具有极高的开发利用价值。
因此,本研究以倒披针叶虫实为研究对象,通过野外设置固定样方定期调查功能性状特征变化,分析不同生境下(不同坡位、不同树冠覆盖度)功能性状时间和空间变化,及其种群数量的动态变化,以解决从种内功能性状变异与种群稳定关系的角度分析倒披针叶虫实种群维持机理,为开发利用该植物提供指导。
1 材料与方法
1.1 研究区概况
本研究区位于新疆准噶尔盆地内的古尔班通古特沙漠(44°11′-46°20′N;84°31′-90°00′E),面积约4.88×104km2,是我国第二大沙漠。年降水70-150 mm,且主要集中于冬季。年蒸发量2000-2800 mm,年均温度5-7℃最高温度40℃以上,年积温3000-3500℃,光热资源丰富,属典型温带干旱荒漠气候,该沙漠属典型的固定与半固定沙漠,沙丘顶部有明显的活化带。冬季积雪层>20cm,并于初春融化补给土壤水,并为短命植物种子萌发提供水分条件。
研究区位于古尔班通古特沙漠腹地莫索湾地区,经纬度为 45°06′N,86°19′E,海拔为 350 m。春夏季为风季,最大风速20 m/s,地下水位13 m 左右。沙丘坡底沙面固定,上部和中部呈活化状态,顶部呈流动状态[24]。在研究区内分布有白梭梭、倒披针叶虫实、弯角四齿芥(Tetracme recurvata)、疏齿千里光(Senecio subdentatus)、尖喙牻牛儿苗(Erodium oxyrrhynchum)等藜科、十字花科(Cruciferae)、菊科(Compositae)、豆科(Leguminosae)、牻牛儿苗科(Geraniaceae)等多种植物。不同坡位物种分布情况参见文献[24]。
1.2 方法
1.2.1 样方设置和研究方法
选取典型固定半固定沙丘,垂直于沙丘走向横贯沙丘设置1条长250 m宽25 m的样带,根据沙丘的相对高度差,将沙丘分为阴坡、阳坡和坡顶,将每个坡面又细分为坡底、坡中、坡上,共7个不同分区,每个坡位选择梭梭树冠覆盖的区域和附近没有树冠覆盖的裸露区域各设置3个样方。样方规格为50 cm×50 cm,7个分区×2树冠覆盖程度×3个样方=42个观测样方。
2016年3-6月在倒披针叶虫实生育期,自3月27日(萌发期)在不同坡位分区间隔20 d进行观测,样方内的个体定株标记[25]并记录。根据沙丘所有植株生长发育情况,将观测样方生育期分为:萌发期(3月 27日)、苗期(4月 13日)、营养生长期(5月4日)、花期(5月 23日)、结实期(6月 12日)。重点观测与植物利用资源及竞争能力等相关的功能性状值株高、冠幅[26-27],同时调查植株密度(D)、死亡率(DR)、萌发率(GR)。
由于野外实验无法准确计算土壤种子库,所以本研究中定义萌发率的计算为:
萌发率=本次调查新萌发幼苗数量/上一次调查时植株萌发总数。
1.2.2 数据处理与分析
在自然群落内,种群性状的平均值主要受环境条件的影响,并且反映该物种的主体生态位,而种内性状差异则表示生态位宽度,二者比值则能够解释高斯悖论和种内性状的变异程度[28]。本研究采用种内性状特征的变异系数量化种内性状。
1.2.2.1 种内功能性状变异分析方法
采用一般线性模型对坡位(种群)内功能性状进行方差分析,模型[29]如下:

其中,ytijk为时间t调查第i坡位内第j个调查样方(包括树冠下和裸地)内第k个个体性状的观测值,μt为第t次调查所有个体的性状平均值,Sti为坡位i内所有个体性状平均值,Ptij为调查样方内所有个体性状平均值,εtijk为实验误差。
根据方差分解结果选择变异程度大且极显著的物候期,对各个坡位(坡位内各个样方)和2种生境种群(树冠下或裸地所有样方)的性状及种内变异进行量化,分析坡位及树冠覆盖度对种内变异的作用。
1.2.2.2 种群数量动态特征
种群大小是物种适应当前环境的直观表现,因此了解种群随时间变化的数量动态有助于种群保护并进一步利用物种资源。本研究通过对不同坡位(种群)调查,利用种群统计学特征萌发率、死亡率、种群密度对整个生长季种群动态变化进行综合描述,并对不同坡位指标进行分析。
1.2.2.3 种群性状变异与种群稳定
为了综合了解种内性状对种群动态稳定的影响,采用多元逐步回归法,研究每个生育期各个坡位所有个体集合(树冠下和裸地)的株高变异系数、冠幅变异系数、种群密度(自变量),与种群死亡率、萌发率(因变量)的关系。数据分析和绘图分别采用Matlab 2015及 Originlab 9.0。
2 结果与分析
2.1 种内性状的时空分布
2.1.1 种内性状的时间分布
通过对倒披针叶虫实不同时间的种群性状值进行方差分析,结果(表1)表明:萌发期,种群内(坡内)性状变异不显著,变异程度低;萌发期后,种内株高及冠幅存在显著的性状变异现象。这主要是由于萌发期个体生长主要利用的子叶提供的营养物质,个体间差异小。随着种群内个体生长发育,进入营养生长期种内性状变异程度迅速增加,说明种群内不同个体具有不同生长速度和物候期。种内性状变异在种群内的变异大于种群间(不同坡位间)的变异。

表1 倒披针叶虫实种群内和种群间功能性状变异分析Tab.1 Variation analysis of functional traits among/within populations inC.lehmannianum
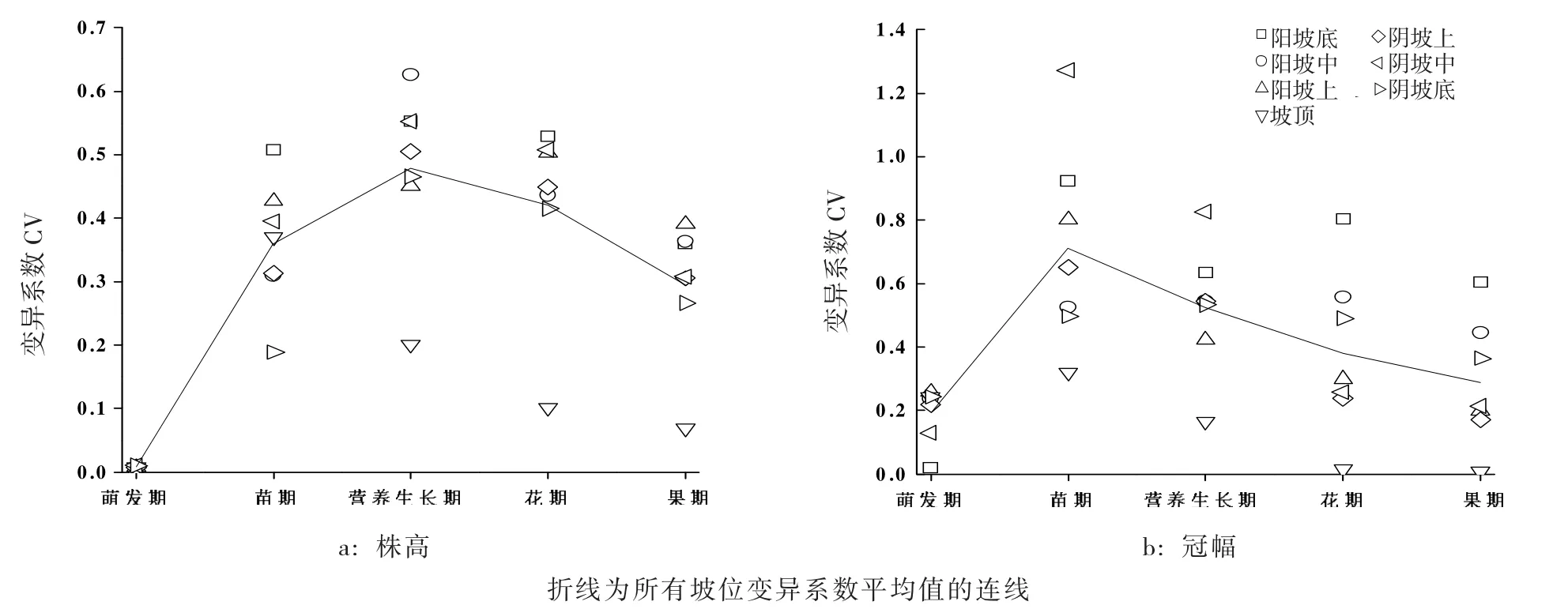
图1 各坡位倒披针叶虫实种群不同物候期株高及冠幅变异系数Fig.1 Coefficient of variation of functional traits in different phenology ofC.lehmannianum
由图1可知:随着生长发育的变化,种内个体差异逐渐增大,但在营养生长期后种内表型相对与性状平均值的离散程度逐渐缩小。
2.1.2 种内性状的空间分布
对营养生长期的调查结果(表2)显示:株高空间分布为坡顶<阴坡<阳坡,变异程度为坡顶<阳坡<阴坡;冠幅大小顺序为坡顶<阳坡<阴坡,变异程度为坡顶<阳坡<阴坡。株高变异程度最高达到63%,冠幅变异程度值最高83%,均出现在坡中位置,说明坡中变异程度较大。
由表2可见:营养生长期阳坡各种群株高差异不显著,阴坡各种群株高差异显著(P<0.01),但均随坡位上升平均高度减小;坡顶株高显著小于阴坡及阳坡各坡位平均值;不同种群间冠幅均呈显著差异;对株高和冠幅的变异系数进行比较发现,2种性状在坡顶的变异程度均最小,坡中部位的种群的变异程度最大。花期阳坡、阴坡各坡位间与坡顶在株高和冠幅2种性状值均差异显著,但各坡位变异系数却有所减小。结实期结果显示,阴坡各坡位间株高性状差异不显著,但每个坡位种群与坡顶及阳坡各坡位种群株高均差异显著,株高及冠幅的变异系数进一步减小。这表明种内变异随物候期变化也存在变化。

表2 不同物候期倒披针叶虫实种内功能性状变异特征Tab.2 Intraspecific functional trait variations in different phenology ofC.lehmannianum
对比不同生境性状,结果(表3)显示:树冠区域的种群更倾向于产生植株高度高、冠幅面积大的植株个体。相应的,树冠区域的倒披针叶虫实种群内株高、冠幅性状标准差也高于裸露区域功能性状标准差,说明在环境条件优越的情况下,种群倾向于产生更明显的功能性状分化以充分占据生态位利用资源。
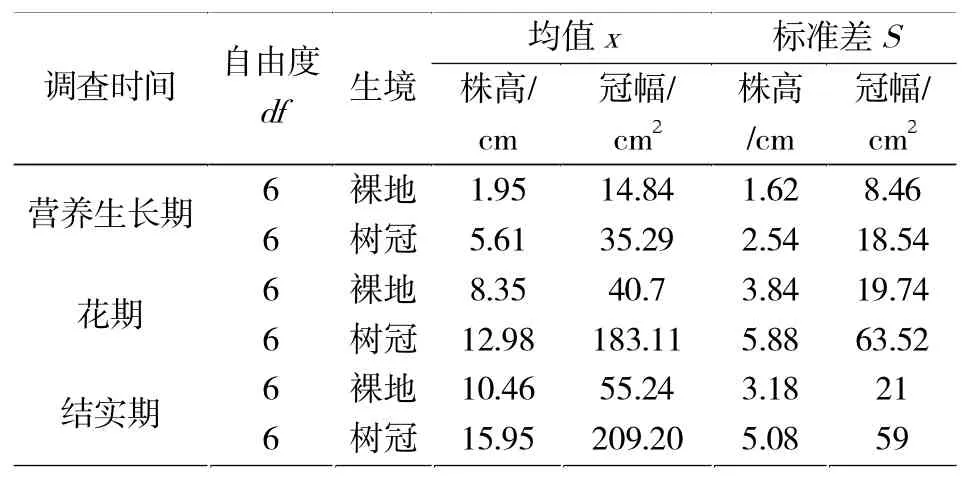
表3 不同生境倒披针叶虫实种内功能性状变异Tab.3 Intraspecific functional traits in different habitats ofC.lehmannianum
2.2 倒披针叶虫实种群动态
由图2可见:倒披针叶虫实种群以坡位为因子进行单因素方差分析表明,种群密度与坡位因素的关系极显著(P=3×10-8<0.01),表明坡位生境是影响种群密度的关键因素。除萌发期外,其余物候期种群密度均差异显著。阴坡阳坡与坡顶的密度比较结果为坡顶<阳坡<阴坡。在时间尺度上,种群密度的变化均成单峰形态,但峰值出现时间不同,表现为坡顶种群在苗期达到最大,其后萌发率急剧下降(图2);阳坡种群与阴坡种群均在营养生长期种群密度达到最大值(图1),说明坡顶的环境波动较大且适宜萌发时间短,导致种群进入苗期后死亡率高于萌发率。

图2 不同坡位倒披针叶虫实种群密度的时间变化Fig.2 Density for 7 partitions of dune ofC.lehmannianumat time scale
综合图1和图2可知,倒披针叶虫实种群萌发率受坡位分区影响显著,苗期坡顶萌发率最高为563%,阴坡中的萌发率最低为59%;后期阴坡各坡位萌发率均值与阳坡各坡位萌发率均值差异并不显著(P=0.44),然而阳坡中与阳坡上种群的萌发率明显高于对应的阴坡中与阴坡上种群,坡顶区域的种群萌发率最低,为18%,其余各坡位萌发率均高于坡顶种群,阴坡种群与阳坡种群萌发率均值依旧差异不显著(P=0.49);花期后的调查显示除坡顶种群外,其余坡位种群均无萌发现象产生(图2)。

图3 整个生育期内不同坡位倒披针叶虫实种群死亡率与萌发率Fig.3 The mortality rate and germination rate at each dune partition during the growing season
由图3可见:倒披针叶虫实不同坡位种群的死亡率在随时间变化明显,总体上呈递减趋势。萌发初期死亡率最高,普遍达到65%左右,花期及果期死亡率较低,低于10%。在整个生长季,并无特定的坡位种群的死亡率一直最高或者最低。
倒披针叶虫实种群的萌发率和死亡率主要集中萌发期与苗期,且这一时期是种群增长的时期(图1)。此时,种群萌发率与种群密度均与生境因素显著相关,表明倒披针叶虫实在不同生境下具有不同的萌发策略:在环境条件恶劣的坡顶位置,萌发数量小,且在整个生长季具有萌发行为;而在其他相对适宜的生境,萌发集中于生长季前期,且萌发量大。
2.3 功能性状变异与种群稳定的关系研究
为量化种内性状变异与种群稳定的关系,用多元逐步回归分析的方法建立回归模型(表4),模型显示:随生长发育影响倒披针叶虫实死亡率的因素在改变,而种群萌发率不受种群密度、种内功能性状(H、C)变异的影响。

表4 种内植株高度变异系数、冠幅变异系数及密度与死亡率回归模型Tab.4 Stepwise regression model betweenCVof functional traits,density and mortality rate,germination rate
由表4可知,倒披针叶虫实种群密度在整个生长季均不影响种群死亡率;苗期种群死亡率与种群密度、种内性状变异无关;营养生长期种群内株高变异系数与种群死亡率呈现极显著的负相关,而与利用光热资源的冠幅变异程度则在模型中被剔除,说明在此时期研究区光热资源不是影响种群存活的限制因素。进入花期后依旧只有种群内株高变异系数影响种群死亡率,然而对种群死亡率的抑制作用减弱,表明株高变异对维持种群稳定作用减弱;对结实期的逐步回归结果显示,种群株高变异系数和冠幅变异系数共同影响种群死亡率。研究表明:在干旱荒漠区倒披针叶虫实种群密度对种群死亡率影响极弱,在萌发期种群死亡率不受功能性状调控,因为萌发期其功能性状并不完善,对个体存活能力的贡献有限;在营养生长期,幼苗建立后性状功能逐步完善,首先采取单一的功能性状差异提升个体存活能力。随生长发育和环境变化,单一性状对个体生存贡献减弱,开始综合利用多个功能性状,冠幅性状与株高性状共同作用于种群死亡率。
3 讨论
3.1 倒披针叶虫实种内性状变异特征
种内变异的生态学作用日益受到关注[21]。本研究表明,倒披针叶虫实种群内存在普遍的功能性状变异,并且种群内大于种群间,说明在干旱区相关研究中种内变异与森林及草原生态系统内的相关研究一样不能被忽略。在时间尺度,种内功能性状随生长发育而不断变化,性状值逐渐上升,而变异程度先上升后下降,在萌发期个体差异较小,营养生长期是个体间功能性状差异上升的重要时期,而花期和果期则是功能性状差异缩小的时期,因为这一时期花期和果期植物开始开花结实,生物量分配不再侧重于功能性状而是繁殖性状,而依旧处于营养期的植株则继续功能性状的生长,直至发育转变到繁殖期。在整个研究区内,种群内功能性状值以及变异随生境的异质变化而变化。与其他种群生境差异较大的坡顶区域种群生态位宽度与变异程度显著低于其余坡位种群,生长在灌木覆盖下的样方内植株性状变异普遍高于裸露区域种群,灌木及裸露区域样方内种群功能特征间差异极显著,表明在荒漠区,灌木对生境的改变作用对于草本植物的生长和分布具有重要影响作用。倒披针叶虫实种群间的差异表明坡位因素影响了功能性状的分布。可见,种内性状变异的来源主要包括遗传因素与环境因素。同时,本研究表明,仅仅调查生长季末时的种内变异程度容易弱化种内变异,因为种内变异在个体营养生长期要大于繁殖期。
3.2 倒披针叶虫实种群动态及其影响因素
每年3月至6月是倒披针叶虫实种群生长季也是北疆风沙发生的主要时期[30]。该物种整个生活史过程几乎均受到风沙的影响并反作用于风沙的活动。在萌发期与苗期种群数量处于增长期,此时死亡率极高,处于种群脆弱期。对比不同生境种群发现,坡中区域是种群数量的主要贡献区,其密度最大。而坡顶沙面活化区域种群规模最小,但冠幅面积最大,且整个生长季均存在萌发的现象,是稳定沙面的重要功能区。可见倒披针叶虫实是活的“草方格”,是生物治沙的重要物种。
生境是影响种群数量稳定的主要因素,而种内性状变异对于生长季中后期物种的存活具有重要影响。然而,目前的研究仍处于对种内功能性状变异的认识及量化阶段[10,19],其对种群稳定贡献的力度仍不明确。本研究结果表明,种内功能性状对种群稳定贡献随生育期变化而变化,并且有贡献作用的种内变异由单一性状变异向着综合性状变异发展。在不同坡位其主体生态位与生态位宽度不同,表明丰富的种内性状变异对于倒披针叶虫实适应不同生境具有重要意义。正是这种性状变异能力减缓了在后期种群数量减少的趋势,是种群稳定性的重要部分。同时本研究发现,在干旱荒漠区密度对于种群稳定作用微弱。
4 结论
倒披针叶虫实种群内存在广泛的性状变异。性状变异特征随时间变化先增加后降低的趋势,在空间上种群内性状存在明显的变异现象,性状变异的程度受到环境的制约。时间尺度的差异主要是随着种群内个体生长发育而产生,随生长发育、性状变异程度,其呈现先上升后下降的趋势。在空间尺度、性状及其变异受坡位及微生境影响,优良环境下,物种密度大,物种竞争强,导致种内个体功能特征分化,减少种内竞争。这表明倒披针叶虫实采取不同程度种内性状及性状变异以适应不同生境。
在干旱荒漠区,种内变异受不同坡位环境的强烈影响,坡位对种群密度与萌发现象的影响显著。密度随生育期变化,整体呈先增加后减小,萌发率与死亡率均随时间变化降低。坡顶种群密度最小,萌发数量最少;坡中密度最大,萌发数量最多。不同坡位种群间早期的萌发数量差异明显,而死亡率则差异较小,表明种群的增长率也受到坡位因素的影响。在生态位宽度小的坡顶区域,其种群数量及增长时间显著低于其余坡位种群,但种群内萌发现象持续存在,这可能是倒披针叶虫实适应干旱区独特的萌发策略,即环境适宜大量萌发,环境恶劣则少量多次萌发。
种内功能性状变异能降低种群死亡率,却不能影响种群萌发率。这说明种内性状变异对种群维持具有一定作用,但这种作用未影响到种群数量的变化方向。在干旱荒漠区,环境因素是决定植物种群动态稳定的控制因子,在环境过滤作用的框架内,种群内普遍存在的种内性状变异现象是物种适应异质环境、缓解环境胁迫和灭绝风险的重要手段。
倒披针叶虫实具有重要的生态功能。本研究表明,在开发利用该植物进行稳沙固沙时应注意播种时期和播种地点。在3月上旬应完成播种,并对整个沙丘不同部位均进行播种,通过不同坡位间子代互相传播等方式,达到保证整个种群维持与沙面稳定。
[1]Violle C,Enquist B J,McGill B J,et al.The return of the variance:intraspecific variability in community ecology[J].Trends Ecol Evol,2012,27(4):244-52.
[2]Dochtermann N A,Gienger C.Individual variability in lifehistory traits drives population size stability[J].Current Zoology,2012,58(2):358-362.
[3]Auger S,Shipley B.Inter-specific and intra-specific trait variation along short environmental gradients in an oldgrowth temperate forest[J].Journal of Vegetation Science,2013,24(3):419–428.
[4]陈磊,米湘成,马克平.生态位分化与森林群落物种多样性维持研究展望[J].生命科学,2014,26(2):112-117.Chen L,Mi X C,Ma K P.Niche differentiation and its consequence on biodiversity maintenance in forest communities[J].Chinese Bulletin of Life Sciences,2014,26(2):112-117.
[5]Bolnick D I,Svanb ck R,Fordyce J A,et al.The ecology of individuals:incidence and implications of individual specialization[J].American Naturalist,2003,161(1):1-28.
[6]陈晓翠,陆嘉惠,韩春,等.多裂骆驼蓬种皮微形态及其粘液特征的分析[J].石河子大学学报(自然科学版),2016,34(6):773-778.Chen X C,Lu J H,Han C,et al.Observation on the seed coat micromorphology and the mucus characteristics of thedesert plantPeganum multisectu[J].Journal of Shihezi University(Natural Science),2016,34(6):773-778.
[7]Dupont Y L,Trφjelsgaard K,Olesen J M.Scaling down from species to individuals:a flower-visitation network between individual honeybees and thistle plants[J].Oikos,2011,120(2):170-177.
[8]Tinker M T,Bentall G,Estes J A.Food limitation leads to behavioral diversification and dietary specialization in sea otters[J].Proceedings of the National Academy of Sciences of the United States of America,2008,105(2):560-565.
[9]Gibert J P,Brassil C E.Individual phenotypic variation reduces interaction strengths in a consumer-resource system[J].Ecology and evolution,2014,4(18):3703-3713.
[10]Pruitt J N,Ferrari M C O.Intraspecific trait variants determine the nature of interspecific interactions in a habitatforming species[J].Ecology,2011,92(10):1902-1908.
[11]Yang L H,Rudolf V H.Phenology,ontogeny and the effects of climate change on the timing of species interactions[J].Ecol Lett,2010,13(1):1-10.
[12]Moran E V,Hartig F,Bell D M.Intraspecific trait variation across scales:implications for understanding global change responses[J].Glob Chang Biol,2016,22(1):137-50.
[13]Reed T E,Gienapp P,Visser M E.Density dependence and microevolution interactively determine effects of phenology mismatch on population dynamics[J].Oikos,2015,124(1):81-91.
[14]党承林.植物群落的冗余结构——对生态系统稳定性的一种解释[J].生态学报,1998,18(6):665-672.Dang C L.Redundancy structure in phytocoenosiums as an explanation of ecosystem stability[J].Acta Ecologica Sinica,1998,18(6):665-672.
[15]Lichstein J W,Dushoff J,Levin S A,et al.Intraspecific variation and species coexistence[J].Am Nat,2007,170(6):807-18.
[16]González-Suárez M,Revilla E.Variability in life-history and ecological traits is a buffer against extinction in mammals[J].Ecology Letters,2013,16(2):242-251.
[17]Araujo M S,Costa-Pereira R.Latitudinal gradients in intraspecific ecological diversity[J].Biol Lett,2013,9(6):20130778.
[18]Jamil T,Ozinga W A,Kleyer M,et al.Selecting traits that explain species–environment relationships:a generalized linear mixed model approach[J].Journal of Vegetation Science,2013,24(6):988-1000.
[19]Moreira B,Tavsanoglu,Pausas J G.Local versus regional intraspecific variability in regeneration traits[J].Oecologia,2012,168(3):671-677.
[20]Kang M,Chang S X,Yan E R,et al.Trait variability differs between leaf and wood tissues across ecological scales in subtropical forests[J].Journal of Vegetation Science,2014,25(3):703-714.
[21]Violle C,Enquist B J,McGill B J,et al.The return of the variance:intraspecific variability in community ecology[J].Trends in Ecology&Evolution,2012,27(4):244-252.
[22]Davidowitz G.Does precipitation variability increase from mesic to xeric biomes[J].Global Ecology and Biogeography,2002,11(2):143-154.
[23]张荣,刘彤.古尔班通古特沙漠南部植物多样性及群落分类[J].生态学报,2012,32(19):6056-6066.Zhang R,Liu T.Plant species diversity and community classification in the southern Gurbantunggut Desert[J].Acta Ecologica Sinica,2012,32(19):6056-6066.
[24]陈正霞,刘彤,刘尊驰,等.沙生植物倒披针叶虫实在古尔班通古特沙漠的分布[J].生态学报,2016,36(13):4064-4073.Chen Z X,Liu T,Liu Z C,et al.Spatial and temporal distribution characteristic of ephemeralCorispermum lehmannianumin the Gutbantunggut Desert[J].Acta Ecologica Sinica,2016,36(13):4064-4073.
[25]何文琴,刘彤,郝晓冉,等.拟南芥不同萌发时期结实种子的母体效应及其适应意义[J].石河子大学学报(自然科学版),2016,34(6):766-772.He W Q,Liu T,Hao X R,et al.The maternal effect and adaptation significance of Arabidopsis thaliana in different seeds germinate time[J].Journal of Shihezi University(Natural Science),2016,34(6):766-772.
[26]Cornelissen J H C,Lavorel S,Garnier E,et al.et al.Handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J].Australian Journal of Botany,2003,51(4):335-380.
[27]Borgy B,Perronne R,Kohler C,et al.Changes in functional diversity and intraspecific trait variability of weeds in response to crop sequences and climate[J].Weed Research,2016,56(2):102-113.
[28]Macarthur R,Levins R.The limiting similarity,convergence,and divergence of coexisting species[J].American Naturalist,1967,101(921):377-385.
[29]李帅锋,苏建荣,刘万德,等.思茅松天然群体种实表型变异[J].植物生态学报,2013,37(11):998-1009.Li S F,Su J R,Liu W D,et al.Phenotypic variations in cones and seeds of naturalPinus kesiyavar.langbianensis populations in Yunnan Province,China[J].Chinese Journal of Plant Ecology,2013,37(11):998-1009.
[30]王雪芹,蒋进,雷加强,等.古尔班通古特沙漠短命植物分布及其沙面稳定意义[J].地理学报,2003,58(4):598-605.Wang X Q,Jiang J,Lei J Q,et al.The distribution of ephemeral vegetation on the longitudinal dune surface and its stabilization significance in the Gurbantunggut Desert[J].Acta Geographica Sinica,2003,58(4):598-605.
The characteristics of intraspecific functional traits variation
and their effects on population stability ofCorispermum lehmannianum
Tang Jingguang1,Liu Tong1*,ZHOU Juan1,Cui Xiangyang2
(1 College of Life Sciences,Shihezi University,Shihezi 832003,China;2 Agricultural and Animal Husbandry Regimental Farm 224,Agricultural Production Division 14,Xinjiang Production and Construction Corps,Hetian,Xinjiang 848116,China)
Corispermum lehmannianumis the dominant specie in the Gurbantunggut Desert,and its importance was next to theHaloxylon ammodendronandHaloxylon persicum.We studied the relationship between intraspecific functional trait variation and the stability of the population,and analyzed the mechanism of the population persistence through the observation of the permanent quadrats at different partitions in dune.The results showed that there was a common intraspecific trait variation(ITV)in the population.The coefficient of variation (CV)of plant height and canopy area increased first and then decreased during the growth.On the spatial scale,the CV under the shrubs'canopy areas was significantly higher than that in the bare area at the same dune partition(P<0.05).The value as well as the CV of plant height and canopy area at the top of the dune was significantly different from that at other partitions except germination period(P<0.05).The ITV was strongly influenced by different slope positions,there were significantly differences in population density and germination among different dune partitions.The population density and germination quantity were minimum at the top of the dune,but were maximum at the middle of the dune,both germination and mortality decreased with time.The results highlight that the population stability ofC.lehmannianumis strongly affected by microenvironment such as different slope positions and different canopy coverage in arid desert area,and the intraspecific trait variation plays an important role for plants to adapt to the desert environment and contributes to the population persistence.
intraspecific trait variation;functional trait;desert area;population stability
Q948
A
10.13880/j.cnki.65-1174/n.2017.04.018
1007-7383(2017)04-0499-07
2017-02-23
国家自然科学基金项目(31260099)
汤景光(1990-),男,硕士研究生,专业方向为农业生物与生态环境工程,e-mail:tangjingguangruyi@163.com。
*通信作者:刘彤(1968-),男,教授,博士生导师,从事植物生态学研究,e-mail:betula@126.com。
猜你喜欢
林业科学(2020年10期)2020-11-30 00:36:56
作文大王·低年级(2020年2期)2020-03-13 08:10:04
数学大王·低年级(2020年2期)2020-03-13 08:09:51
作文周刊·小学四年级版(2019年8期)2019-04-23 01:04:16
绿色科技(2017年1期)2017-03-01 10:14:33
湖南林业科技(2017年1期)2017-02-06 05:29:18
国际木业(2016年6期)2016-02-28 10:27:03
国际木业(2016年5期)2016-02-28 10:26:34
防护林科技(2014年7期)2014-03-01 02:33:16
城市道桥与防洪(2013年11期)2013-03-11 15:19:29
