
氮、磷添加对贝加尔针茅草原凋落物分解的影响
2017-09-03 08:59:08李文亚邱璇白龙杨殿林
草业学报 2017年8期
李文亚,邱璇,白龙,杨殿林*
(1.农业部环境保护科研检测所,天津 300191;2.沈阳农业大学园艺学院,辽宁 沈阳110866)
氮、磷添加对贝加尔针茅草原凋落物分解的影响
李文亚1,2,邱璇1,白龙2,杨殿林1*
(1.农业部环境保护科研检测所,天津 300191;2.沈阳农业大学园艺学院,辽宁 沈阳110866)
凋落物分解是草原生态系统养分循环和能量流动的主要途径。养分是温性草原生产力主要限制因素之一。本文主要研究了养分添加对草原凋落物分解的影响,为草原生态系统养分管理和天然草地的恢复建设提供理论依据。试验以内蒙古贝加尔针茅草原为研究对象,使用网袋分解法,实验材料为贝加尔针茅(Sb)、羊草(Lc)以及冷蒿(Af)3种植物的根、茎、叶组织,设置4个处理:CK、N、P和NP(CK:不施任何肥料;N:硝酸铵;P:重过磷酸钙;NP:硝酸铵和重过磷酸钙)。结果表明,1) N、P、NP添加下,3种植物的残留率显著低于无养分添加,氮磷添加对凋落物分解有显著的促进作用。2)分解过程中,叶和根凋落物N元素前期释放,后期有轻微的富集现象,而茎凋落物呈现富集-释放-富集的模式。P元素和C元素基本呈现持续释放的模式。3)磷元素的残留率在N、P以及NP处理条件下均呈下降的趋势,与植物器官残留率的变化趋势相同。4)羊草各器官凋落物分解95%所用的时间为2.95~3.96年,贝加尔针茅为3.32~3.77年,冷蒿为2.64~4.89年。5)不同植物以及植物的不同器官凋落物分解速率对氮、磷添加的响应不同。
贝加尔针茅草原;凋落物;氮、磷添加;分解速率
凋落物是植物生长发育过程中的产物,是草地生态系统的重要组成部分,推动着土壤有机质的矿化分解和土壤养分的循环与转化,对维持草原生态系统过程和功能具有重要作用[1]。养分是草地生态系统的主要限制因素之一。养分添加是提高土壤肥力,恢复草原植被的有效措施。畜牧业发达国家通常将草原施肥作为草原管理的重要手段[2]。凋落物在分解过程中释放的N、P、K是植物生长发育不可缺少的营养元素,对植物生长发育和草原生产力的提高有很好的促进作用[3]。凋落物的分解也是维持草原生态系统物质循环和能量流动的主要动力,是草原生产力的重要组成部分[4]。因此草地凋落物分解研究已逐渐被重视。
国内外,最早对于凋落物的研究起始于森林生态系统,研究方向大多集中在凋落物对于增加土壤的肥力以及改善物质循环方面所起的作用[5-6]。1876年就已经开始研究凋落物的分解过程中养分的循环过程[7],凋落物分解被视为生态系统功能中较为重要的初级生产力。养分添加对凋落物分解影响的研究结果存在很大的争议[8]:有些研究发现土壤氮素增加能够促进凋落物的分解[9],而有些研究发现没有作用[10],甚至是抑制凋落物的分解[11]。草地凋落物分解作为生物地球化学循环重要组成部分的研究逐渐被重视[12]。但大多研究主要涉及叶片等地上部凋落物,或者单一凋落物的分解动态。对于地下部根系凋落物分解以及混合凋落物分解等关注不够[13-15]。有研究表明,根系凋落物分解比地上部凋落物分解要慢[16];有的研究则发现根系凋落物分解要快于地上部凋落物[17];而另一些研究结果显示,根系凋落物和地上部凋落物分解的差异取决于植物种类,有的种类快,有的种类慢[18]。已有研究大多集中于单一凋落物分解,混合凋落物分解的研究不多[4,19]。王其兵等[20]对草甸草原和典型草原的混合凋落物分解进行过研究。现实环境中,不同种类凋落物通常是混合在一起分解的,很可能发生混合效应(混合凋落物的分解速率与期望分解速率显著不同,或促进或抑制)[4,14],但也可能不发生混合效应[21]。单一凋落物分解特征的研究,很难说明其结果能在多大程度上反映混合凋落物作为一个整体进行分解的实际特征[22]。相关研究表明混合效应产生可能与凋落物化学组成有关[23],也可能与混合后导致的物理化学环境改变有关[24],或者与分解者群落和活性有关[25],还可能与植物多样性有关[26]。
全球变化背景下,混合凋落物分解的混合效应是否普遍存在,混合效应的发生与否及其效应方向到底与哪些因子有关,迄今为止,还不清楚混合效应发生的规律以及引起混合效应的原因,这些问题的存在限制了人们更好地了解草原生态系统碳和养分的循环。贝加尔针茅(Stipabaicalensis)草原是亚洲中部草原区所特有的草原群系,是草甸草原的代表类型之一,是开展温带草原生态系统对养分添加响应研究的理想平台之一[13]。探讨N、P、K及其复合养分添加对贝加尔针茅草原3种主要植物地上部凋落物和地下根系分解的影响以及不同植物种单一凋落物分解动态和混合凋落物分解的混合效应,为制定草原合理的施肥管理措施提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古呼伦贝尔市鄂温克旗境内地势平坦、开阔的地区。地理坐标为北纬48°27′-48°35′,东经119°35′-119°41′,海拔763 m。该地区属于温带草甸草原区,半干旱大陆性季风气候,年均气温-1.6 ℃,年降水量328.7 mm,年蒸发量1478.8 mm,年积温2567.5 ℃,无霜期113 d,土壤类型为暗栗钙土。植被优势物种为羊草(Leymuschinensis)、贝加尔针茅,冷蒿(Artemisiafrigida)、羽茅(Achnatherumsibiricum)、日荫菅(Carexpediformis)、祁州漏芦(Rhaponticumuniflorum)、扁蓿豆(Pocockiaruthenica)、草地麻花头(Serratulayamatsutanna)、变蒿(Artemisiacommutate)等为常见伴生种,共有植物66种,分属21科49属[27]。
1.2 试验设计
在贝加尔针茅草甸草原选择地势平坦,植被有代表性的地段设置试验样地,试验设CK、P、N、NP 4个处理,N是硝酸铵,P是重过磷酸钙Ca(H2PO4)2·H2O,施肥量按照10 g/m2纯N、P量换算为硝酸铵和重过磷酸钙的施用量6次重复,随机区组排列。小区面积8 m×8 m,重复与小区间分别设5和2 m隔离带。养分添加试验于2010年开始进行,每年分两次进行添加,分别在牧草生长季6月15日、7月15日进行,每次施入全年添加总量的50%。施肥时均匀手撒。
1.3 试验材料与方法
采用凋落物网袋法,依据试验区域的植被物种优势度和物种特性,选择多年生禾本科羊草、多年生丛生草贝加尔针茅和小半灌木冷蒿3种植物为研究对象。在生长季节结束时在未经试验处理的草地上收集新凋落或枯死植株上的组织(叶、茎),同时收集植株根系部分。将收集的凋落物风干后剪成长度为10 cm的条段状,放入孔径为0.2 mm,大小为15 cm×25 cm的尼龙网袋,每袋干重为8.0 g。装袋时保证凋落物均匀平展地充满整个网袋,放入相应的标签。每种组合在每个处理小区放置3袋,地上部分(叶,茎)用铁丝固定在土壤表面,地下部分(根)则埋入以铁锹倾斜45°轻轻掀开的土壤中,并尽量不破坏土壤原来的状态,每网袋间隔0.5 m。样品在2011年8月10日放入样地,分别于2012年6月20日、8月20日和10月20日即分解295、356、417 d后收回样品。取回的样品剔除杂质,将凋落物放入信封在烘箱中70 ℃ 烘48 h,称重并用于元素分析。有机碳用水合热重铬酸钾氧化-比色法,全氮用H2SO4-H2O2消煮,消煮时,称风干样品0.08~0.10 g左右,于消煮管中(加1~3滴水湿润样品),加浓硫酸5 mL,放置过夜,过夜后在消煮炉上先250 ℃消煮(30 min),待H2SO4分解冒出大量白烟后再升高温度至300 ℃,当溶液呈均匀的棕黑色时取下,稍冷后加10 滴 H2O2,摇匀,再加热至微沸,消煮约5 min,取下稍冷后,重复加 H2O25~10滴,再消煮。如此重复3~5次,每次添加的H2O2的量应逐次减少,消煮到溶液呈无色或清亮后(应该为水的颜色),再加热5~10 min,以除尽剩余的H2O2。将消煮液无损地洗入100 mL 容量瓶中,用水定容,摇匀。过滤或放置澄清后供氮、磷的测定[28],之后使用SEALAA3连续流动分析仪(AA3,天津)进行N浓度测定,全磷采用分光光度计(UV-2800H,上海)测定。
1.4 数据分析
凋落物分解速率用k表示,采用负指数衰减模型Xt/X0=ae-kt,其中Xt代表凋落物在分解一段时间t后残留的重量,X0表示凋落物未分解时的初始重量,a为拟合参数,k为凋落物分解系数[g/(g·a)],t为凋落物的分解时间,一般以年表示。元素残留率用R表示,R=[(Ct×Xt)/(C0×X0)]×100%,Ct表示t时刻凋落物元素含量(mg/g),Xt表示t时刻凋落物的干重(g),C0表示初始元素含量(mg/g),X0为初始干重(g)。凋落物元素释放率用NR表示,NR=100-R,其中R表示元素残留率。干重残留率及养分动态变化使用SPSS 17.0进行单因素方差分析,比较各植物器官不同养分添加条件下干重残留率、有机碳、全氮、全磷残留率的变化趋势及显著性水平差异(P<0.05)。
2 结果与分析
2.1 凋落物初始化学成分分析
对贝加尔针茅、羊草以及冷蒿3种植物根、茎以及叶器官凋落物初始化学成分分析表明(表1),凋落物中不同元素的起始浓度为C>N>K>P,不同植物同一器官以及相同植物不同器官间,初始元素化学成分组成存在着一定的差异。羊草、贝加尔针茅和冷蒿3种植物的叶凋落物的C、N含量都高于根茎,羊草和冷蒿叶的全P含量均显著(P<0.05)高于根茎,总体来说地上部分C、N、P含量均高于相应的地下组织。
2.2 氮、磷添加对凋落物重量变化的影响
在凋落物分解过程中,衡量凋落物分解速度快慢的指标之一就是凋落物的重量变化,图1表明,氮磷养分添加条件下(N、P、NP)各凋落物干重残留率均呈持续下降趋势,在分解295、356和417 d时养分添加的残留率均显著低于无任何养分添加处理,养分添加对凋落物分解表现出明显的促进作用,各养分添加处理的残留率之间则无显著差异。分解至417 d时,无养分添加的羊草、冷蒿、贝加尔针茅叶凋落物残留率分别为83%、73%、81%,而养分添加条件下,各叶凋落物残留率则在43%到53%之间。
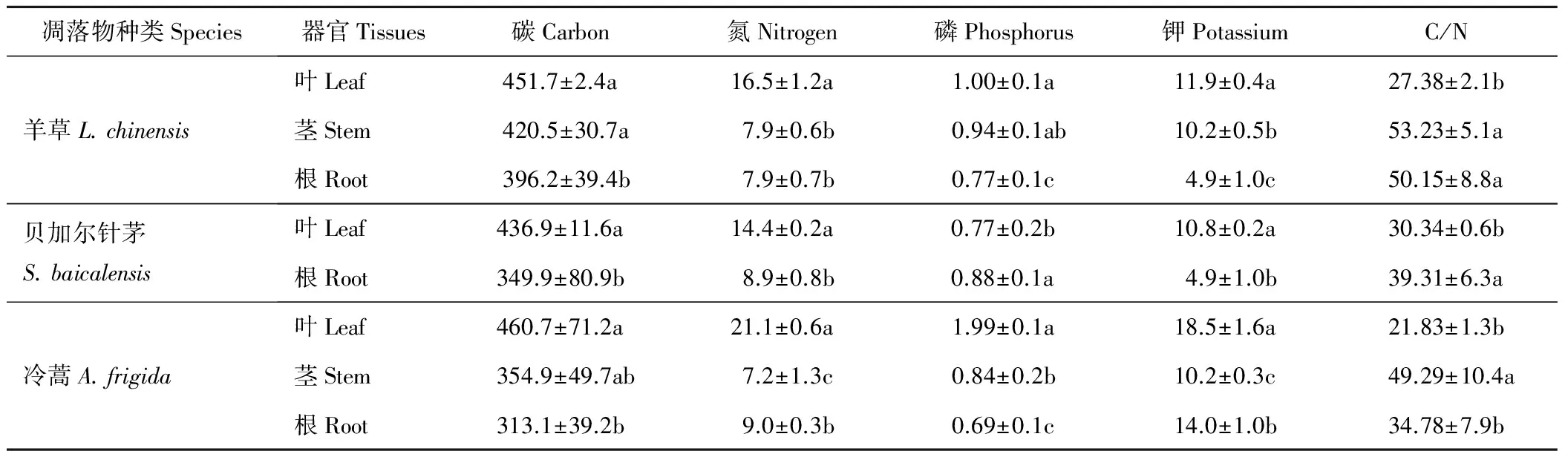
表1 各凋落物初始化学元素组成
注:表中数值为平均值±标准差,同列不同字母表示差异显著(P<0.05)。
Note: The data in the Table represent mean±SD. Different letters in the same colum indicate significant differences atP<0.05.
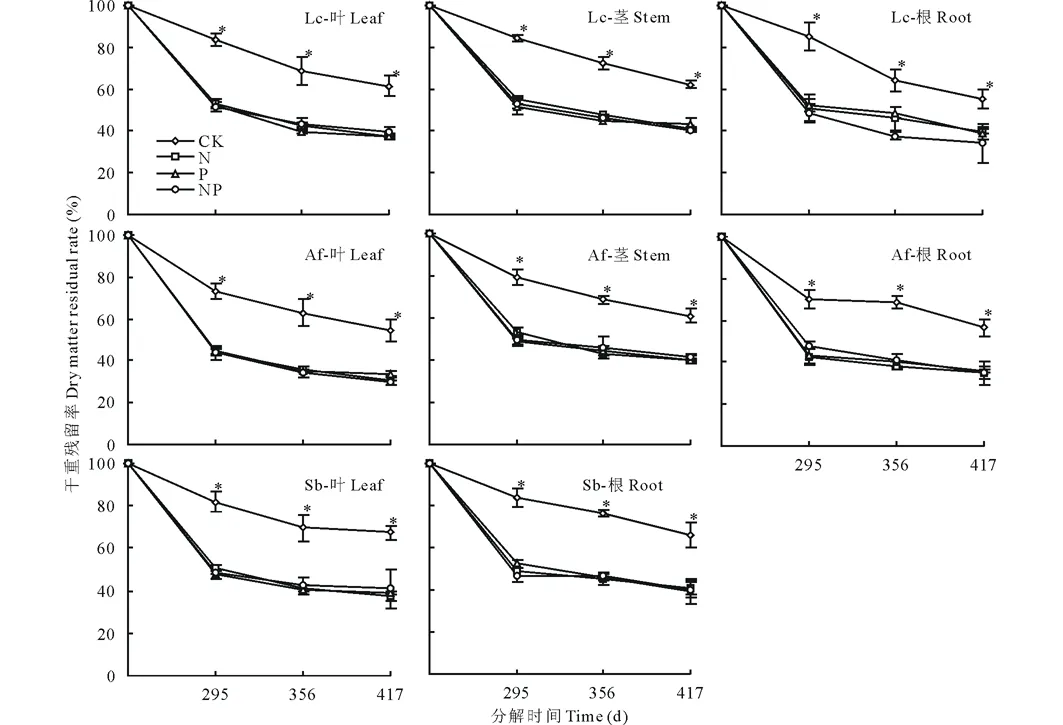
图1 养分添加下各器官凋落物残留率变化Fig.1 Residual rate changes in difference plant organ litters under the nutrient addition *表示不同处理间差异达到显著水平 (P<0.05)。*means the significant variation among different treatments (P<0.05).Lc:羊草Leymus chinensis; Af: 冷蒿Artemisia frigida; Sb:贝加尔针茅 Stipa baicalensis. 下同The same below.
2.3 凋落物分解速率模型
各器官对N、P、NP添加的响应各不相同,3种植物地上地下各器官半分解时间及分解95%时间都有所下降,都表现出前期分解较快,后期分解慢的趋势,且前期各处理分解速率明显快于对照,后期促进效果并不明显(图1)。其中羊草各器官凋落物分解系数在0.794到1.051之间,叶凋落物在N添加下周转期最短,根据Olson分解模型分解95%时间排序为CK>NP>P>N;羊草茎凋落物分解95%的时间顺序为CK>NP>N>P,羊草根凋落物分解95%时间顺序为CK>N>P>NP,在NP添加下周转期最短。茎叶凋落物对N、P添加响应并不明显,CK对羊草根凋落物与茎叶凋落物影响差异不同(表2);贝加尔针茅各器官凋落物在不同养分添加下分解系数变化在0.732到1.270之间,周转期为3.32到3.77年,贝加尔针茅叶对养分添加的响应依据Olson模型的T0.95排序为CK>P>N>NP,根凋落物的T0.95排列为CK>N>P>NP,与羊草根凋落物对养分添加的响应趋势相同;冷蒿各凋落物不同器官的半分解时间较羊草各部分半分解时间短,但分解95%的时间却没有相应的变化,冷蒿叶凋落物对养分添加的响应与贝加尔针茅叶凋落物相同,其周转期为CK>P>N>NP,而冷蒿根凋落物的T0.95与羊草根相同,为CK>N>P>NP。总体来说,其中分解最快的是NP添加下的冷蒿叶凋落物,分解95%的时间为2.64年,分解最慢的是无任何养分添加的冷蒿根凋落物,分解95%的时间为4.89年。
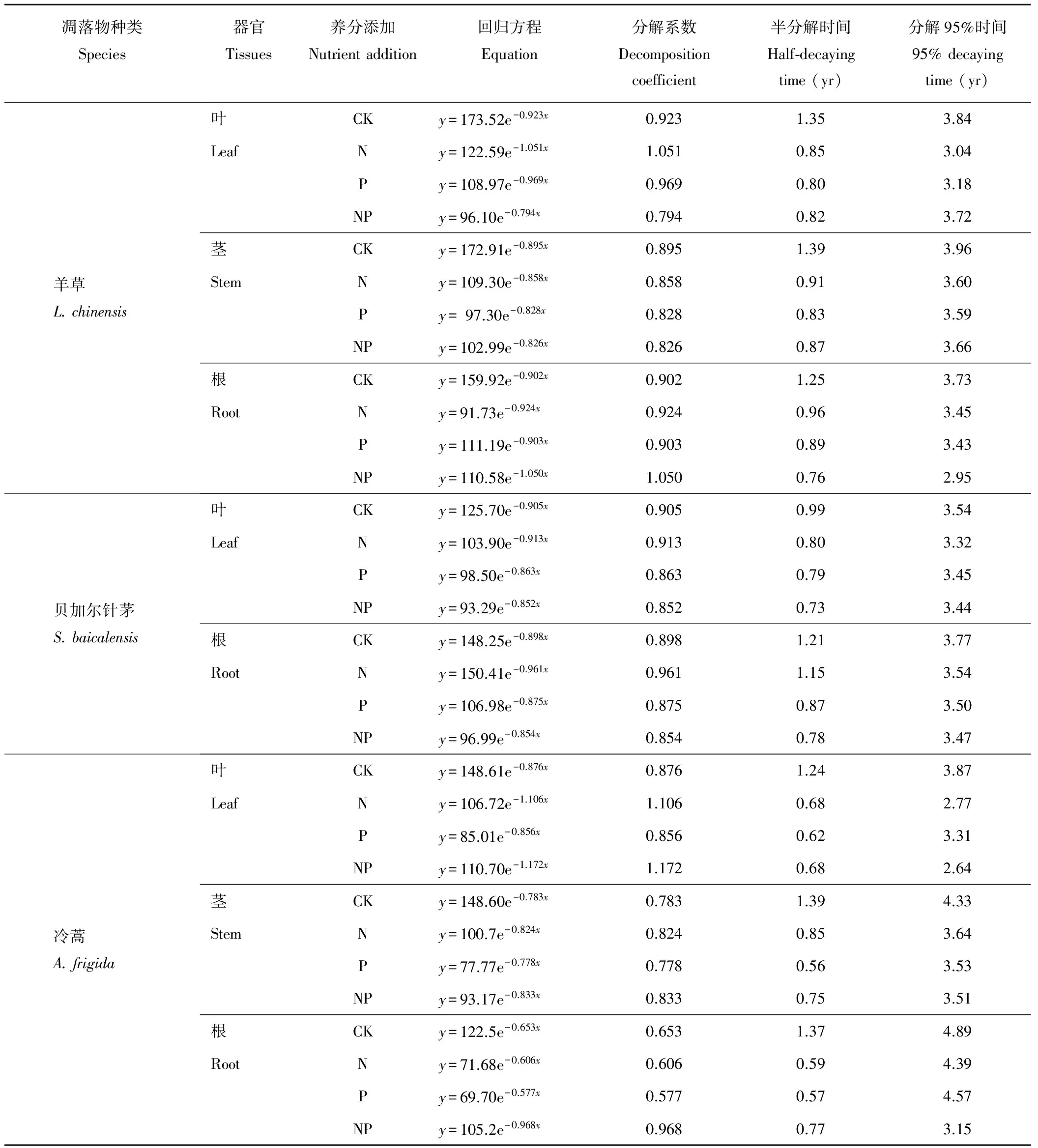
表2 3种植物凋落物不同器官残留率随时间的指数方程
2.4 氮磷添加对凋落物C、N、P释放的影响
有机碳是构成凋落物的重要元素,有机碳的变化情况,可以从侧面反映凋落物的分解情况。图2表明,CK、N、P、NP条件下3种植物的有机碳残留率总体呈持续下降趋势,与凋落物质量残留率变化趋势相近,N、P、NP添加均对有机碳的释放有明显的促进效果,但不同植物不同器官对各养分添加响应不尽相同。其中对照组的羊草和冷蒿茎凋落物在分解初期分解速率较慢,分解初期主要是淋溶作用,茎的C/N较其他器官高,可能造成这一差异。
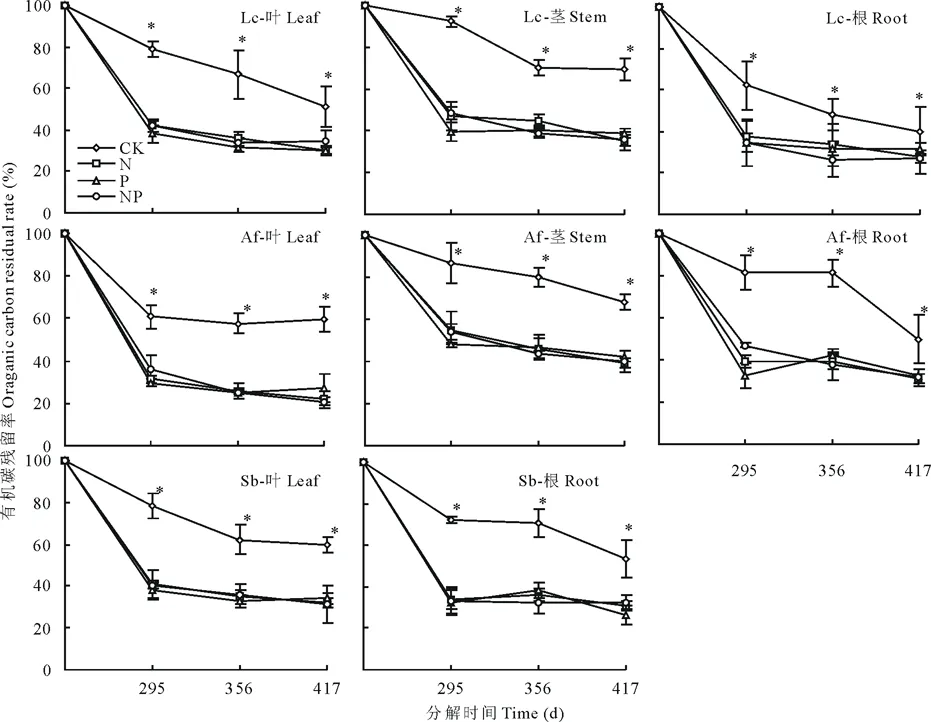
图2 氮、磷添加对有机碳残留率的影响Fig.2 Organic carbon residual rate influenced by N, P addition
N、P、NP添加条件下,3个物种各器官凋落物氮元素的释放模式未发生明显改变,不同物种、不同器官对各养分添加响应各不相同(图3)。3种植物叶凋落物和根凋落物在356 d之前均呈持续上升趋势,在356 d达到最大释放率后,呈现出一定的下降趋势,叶凋落物在8到10月之间下降并不明显,而3种根凋落物则有比较明显的富集作用;羊草和冷蒿的茎凋落物在295 d之前氮元素呈现富集趋势,在356 d时达到最大释放率,之后趋势与根、叶相同。P添加对3种植物的叶片凋落物N元素的释放有显著(P<0.05)的抑制作用,3种植物N释放率显著低于无养分添加;NP添加对贝加尔针茅叶凋落物在各个时期都有显著的抑制作用。N、P添加对冷蒿茎N释放有显著促进效果。N、NP添加对羊草根凋落物的N释放有抑制作用。

图3 氮素和磷素添加下各器官氮释放率变化Fig.3 Organ nitrogen release rate under the treatment of nitrogen and phosphorus addition
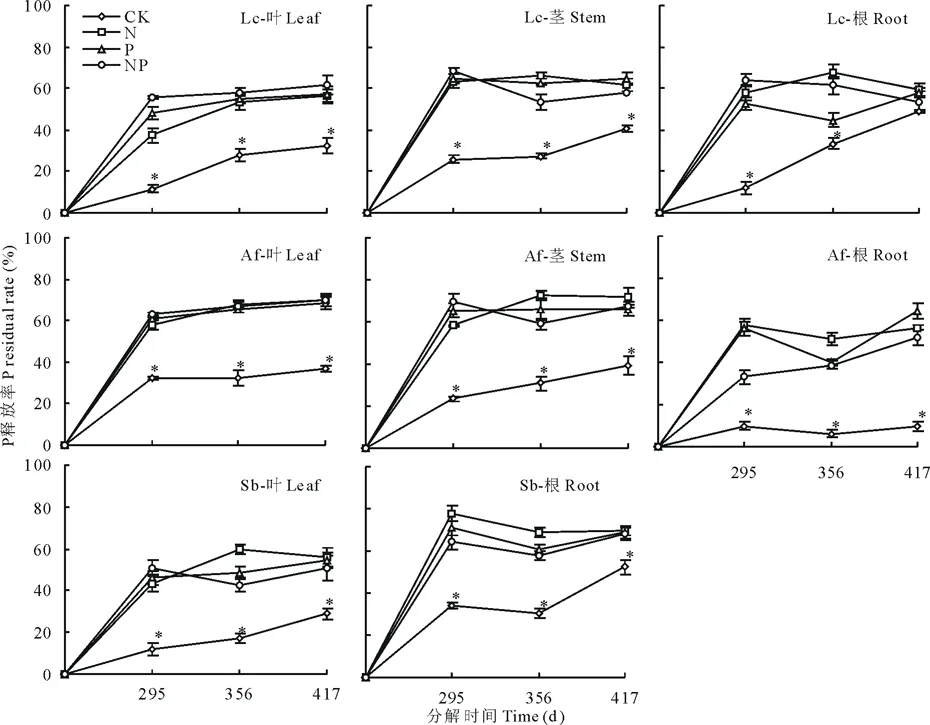
图4 养分添加下3种植物各器官P释放率变化Fig.4 P release rate of the plant organ litters under the nutrient addition
各养分添加条件下,3种植物各个器官的P释放率显著(P<0.05)高于无养分添加(图4),与干重的残留率趋势相近,即养分添加促进了各凋落物P元素释放速率。
3种植物叶凋落物的P释放率呈持续释放模型,且在分解至417 d时各养分添加之间无显著差异,而3种植物根凋落物则在295到356 d之间出现轻微的富集现象,N添加的羊草根凋落物和冷蒿茎凋落物释放率高于NP添加,P添加的冷蒿根凋落物和羊草茎凋落物的磷素释放率显著(P<0.05)高于N、NP添加下的释放率。各养分添加下3种植物叶片凋落物在分解至417 d时P残留率无显著差异。
3 讨论
3.1 养分添加和无养分添加对凋落物分解速率的影响
本试验在贝加尔针茅草原进行了417 d的凋落物野外分解,氮、磷添加对3种植物凋落物影响各不相同,同一植物的不同器官对氮素和磷素添加响应也存在差异。凋落物的分解是一个复杂的物理、生物和化学过程,养分含量主要受这三大因素的影响[3]。分解前期,即从放入凋落物之后12个月之间凋落物都持续分解,此阶段主要是淋溶作用,CK、N、P、NP添加表现出不同程度的抑制或促进作用。295 d时N添加下羊草、贝加尔针茅、冷蒿叶凋落物的瞬时分解速率高于无养分添加,与弓晓静[29]所做氮磷添加对松林凋落叶分解的影响研究结果相同,其研究发现,N添加对松林的叶凋落物前12个月表现为促进效应,但这种促进效应到18个月的时候已经不再明显。无养分添加的3种植物叶凋落物的分解速率均大于茎和根,可能是茎和根中含有较高的木质素与纤维素等不易分解的物质[30];且养分添加会改变土壤微生物的群落结构组成及其对底物的利用方式,因此养分添加的分解速率高于无养分添加[31]。
3.2 N、P以及NP添加对凋落物分解的影响
在自然生态系统中,植物个体的生产能力往往会受到时间和空间等多因素的共同影响,氮是生态系统中对植物生长具有限制性的一个重要因子,虽然磷的作用不如氮突出,但是磷有时也是限制植株生长的一个重要因子[32]。美丽班·马木提[33]在氮磷添加对天童木荷凋落叶分解的影响中发现施肥处理可以改变木荷叶凋落物的部分特征,但是在不同的施肥处理下,凋落物的化学特征受其影响的显著性方面存在着一定的差异,当单独施用N、P肥时,对叶凋落物的分解速率影响不明显,当两者同时施加时,可以明显抑制叶凋落物的分解,N、P添加对凋落物的分解速率还与凋落物中初始的碳氮比有关。李仁洪等[34]对慈竹不同器官凋落物分解进行模拟氮沉降处理,结果表明,氮沉降可以促进竹子叶、箨以及枝的凋落物分解速率,但是当氮素含量升高时,促进作用逐渐地减慢;陈翔[35]的研究也发现,适当的氮处理对凋落物的分解有着一定的促进作用,但随着时间的增加或者氮素水平的升高,这种促进水平会延缓甚至改为抑制作用。陈昊[36]对亚热带北部常绿阔叶林叶凋落物分解的研究表明,磷元素在不同环境下会出现滞固现象,磷元素的释放主要以复合函数的形式进行。本研究中,羊草、贝加尔针茅、冷蒿3种植物的叶茎凋落物的有机碳含量、氮含量均高于相应的根凋落物。氮、磷添加下,3种植物的分解速率呈先上升后下降的趋势,且在356 d时分解速率达到了最高峰,然后随着时间的推移,慢慢减弱。后期NP处理的分解速率要低于单一氮处理、单一磷处理以及对照,是因为NP处理在后期对凋落物的降解有着明显的抑制作用,这与弓晓静等[37]的研究结果相同;陈瑶等[38]模拟氮磷沉降对华南热带森林尾叶桉木质残体分解和养分动态影响的研究表明,氮添加使得木质残体分解速率常数降低,而磷添加则显著提高了分解常数,在缺少氮素的条件下,磷元素的可利用性限制了木质素残体的分解速率,氮添加使得磷元素的迁移方式由直接释放改变为淋溶—富集—释放,而磷元素的添加使得磷元素的迁移方式改变为富集—释放,氮、磷对木质素残体的分解速率以及养分的使用模式不同。CK、N、P、NP条件下3种植物的有机碳残留率总体呈持续下降趋势,从凋落物质量残留率的变化趋势来看,其与碳元素的残留率趋势相近。这是由于当C/N越低时,凋落物中耐分解的有机化合物就会越少,从而凋落物的分解就会加快[39-40]。
N、P、NP 3种处理条件下,3种植物的残留率显著低于对照组,氮磷添加对凋落物分解有显著的促进作用,这与陈昊[36]的实验结论相符。其原因可能是养分的添加能提高土壤中微生物代谢活性与物种丰富度,且有效促进微生物碳源利用能力,从而提高了凋落物的分解速率。
3.3 植物不同组织元素释放模式
在凋落物分解过程中,叶和根凋落物N元素前期释放,后期有轻微的富集现象,茎凋落物呈现富集-释放-富集的模式,这是因为微生物在占领凋落物的同时,将土壤中的氮转移到凋落物上,从而导致凋落物的氮呈现富集状态[40]。P元素和C元素基本呈现持续释放的模式。另外,凋落物的分解速率不同,与凋落物放置位置的地上生物也有一定的关联,Gabriel等[41]在凋落物分解的生物过程和分解生物群落的交互作用中提到,不同土壤生物群落之间的相互作用和凋落物的初始质量以及类型不同可能导致分解微生物数量和比例的不同,进而影响分解过程。
4 结论
研究发现,4种不同处理条件下,3种植物器官凋落物随着时间的增加,其残留率呈现出了持续下降的趋势,且处理组要明显低于对照组,氮磷添加对凋落物分解有显著的促进作用。在凋落物分解过程中,叶和根凋落物N元素前期释放,后期有轻微的富集现象,而茎凋落物呈现富集-释放-富集的模式。P元素和C元素基本呈现持续释放的模式。N、P、NP添加对混合凋落物的分解速率有显著的促进,但对混合凋落物的加性效应并无显著影响。凋落物分解速率受环境、养分添加和器官组成等各方面因素的影响,氮素和磷素添加能够促进3种凋落物的分解,加快草原养分循环。混合凋落物的组成对凋落物分解影响机制以及养分添加的成分及比例还有待进一步的长期研究。
References:
[1] Carrera A L, Bertiller M B. Combined effects of leaf litter and soil microsite on decomposition process in arid rangeland. Journal of Environmental Management, 2013, 114(2): 505-511.
[2] He X. Effect of Nutrient Additions on Community Biomass and Diversity Varied Among Different Grassland Ecosystems of Inner Mongolia[D]. Hohhot: Inner Mongolia University, 2014. 贺星. 养分添加对内蒙古草原生物量和多样性的影响[D]. 呼和浩特:内蒙古大学, 2014.
[3] Lin S W, Wu C Z, Hong W,etal. Analysis of leaves nutrient dynamics by wave-type time series method. Journal of Fujian College of Forestry, 2009, 29(3): 215-219. 林淑伟, 吴承祯, 洪伟, 等.凋落叶养分动态的起伏型时间序列分析. 福建林学院学报, 2009, 29(3): 215-219.
[4] Chen J, Li Y, Huang J H. Decomposition of mixed litter of four dominant species in an Inner Mongolia steppe. Chinese Journal of Plant Ecology, 2011, 35(1): 9-16. 陈瑾, 李扬, 黄建辉. 内蒙古典型草原4种优势植物凋落物的混合分解研究. 植物生态学报, 2011, 35(1): 9-16.
[5] Zhu J, Liu J, Zhu Q. Hydro-ecological functions of forest litter layers. Journal of Beijing Forestry University, 2002, 24(6): 30-34.
[6] Ogée J, Brunet Y. A forest floor model for heat and moisture including a litter layer. Journal of Hydrology, 2002, 255(1-4): 212-233.
[7] Li X, Sun J, Chang Z,etal. A study on litter and forest floor in Tianshan Mountain. Acta Pedologica Sinica, 1997, 34(4): 406-417.
[8] Keeler B L, Hobbie S E, Kellogg L E. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition. Ecosystems, 2009, 12(1): 1-15.
[9] Hobbie S E, Gough L. Litter decomposition in moist acidic and non-acidic tundra with different glacial histories. Oecologia, 2004, 140(1): 113-124.
[10] Dukes J S, Field C B. Diverse mechanisms for CO2effects on grassland litter decomposition. Global Change Biology, 2000, 6(6): 145-154.
[11] Micks P, Aber J D, Boone R D,etal. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology & Management, 2004, 196(1): 57-70.
[12] Stevens C J, Dise N B, Mountford J O,etal. Impact of nitrogen deposition on the species richness of grasslands. Science, 2004, 303: 1876-1879.
[13] Yang D L, Han G D, Hu Y G,etal. Effects of grazing intensity on plant diversity and aboveground biomass ofStipabaicalensisgrassland. Chinese Journal of Ecology, 2006, 25(12): 1470-1475. 杨殿林, 韩国栋, 胡跃高, 等.放牧对贝加尔针茅草原群落植物多样性和生产力的影响. 生态学杂志, 2006, 25(12): 1470-1475.
[14] Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter. Oikos, 2004, 104: 2300-2346.
[15] Zhang C H, Zhang L M, Liu X R,etal. Root tissue and shoot litter decomposition of dominant speciesStipabaicalensisin Hulun Buir meadow steppe of Inner Mongolia, China. Chinese Journal of Plant Ecology, 2011, 35(11): 1156-1166. 张彩虹, 张雷明, 刘杏认, 等. 呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解. 植物生态学报, 2011, 35(11), 1156-1166.
[16] Fujii S, Takeda H. Dominant effects of litter substrate quality on the difference between leaf and root decomposition process above- and belowground. Soil Biology & Biochemistry, 2011, 42: 2224-2230.
[17] Liu Z K, Wang S P, Han J G,etal. Litter decomposition and nutrient dynamics research of typical grassland plant and root in temperate Inner Mongolia. Acta Prataculturae Sinica, 2005, 14(1): 24-30. 刘忠宽, 汪诗平, 韩建国, 等. 内蒙古温带典型草原植物凋落物和根系的分解及养分动态的研究. 草业学报, 2005, 14(1): 24-30.
[18] Berg B, Davey M P, Marco A D,etal. Factors influencing limit values for pine needle litter decomposition: a synthesis for boreal and temperate pine forest systems. Biogeochemistry, 2010, 100(1-3): 57-73.
[19] Bonanomi G, Incerti G, Antignani V,etal. Decomposition and nutrient dynamics in mixed litter of mediterranean species. Plant and Soil, 2010, 331(1): 481-496.
[20] Wang Q B, Li L H, Bai Y F,etal. Effects of simulated climate change on the decomposition of mixed litter in three steppe communities. Acta Phytoecologica Sinica, 2000, 24(6): 674-679. 王其兵, 李凌浩, 白永飞, 等. 模拟气候变化对3种草原植物群落混合凋落物分解的影响. 植物生态学报, 2000, 24(6): 674-679.
[21] Smith C V, Bradford A M. Do non-additive effects on decomposition in litter-mix experiments result from differences in resource quality between litters. Oikos, 2003, 102(2): 235-242.
[22] Hui D, Jackson R B. Assessing interactive responses in litter decomposition in mixed species litter. Plant and Soil, 2009, 314(1-2): 263-271.
[23] Rouifed S, Handa I T, David J F,etal. The importance of biotic factors in predicting global change effects on decomposition of temperate forest leaf litter. Oecologia, 2010, 163(1): 247-256.
[24] Wardle D A, Nilsson M C, Zackrisson O,etal. Determinants of litter mixing effects in a Swedish boreal forest. Soil Biology and Biochemistry, 2003, 35(6): 827-835.
[25] Chen S, Steinberger Y. Microbial functional diversity associated with plant litter decomposition along a climatic gradient. Microbial Ecology, 2012, 64(2): 399-415.
[26] Scherer-Lorenzen M. Functional diversity affects decomposition processes in experimental grasslands. Functional Ecology, 2008, 22(3): 547-555.
[27] Li W J, Liu H M, Yang D L,etal. Effects of nitrogen and water addition on plant species diversity and biomass of common species in theStipabaicalensisSteppe, Inner Mongolia, China. Acta Ecologica Sinica, 2015, 37(19): 6460-6469. 李文娇, 刘红梅, 杨殿林, 等. 氮素和水分添加对贝加尔针茅草原植物多样性及生物量的影响. 生态学报, 2015, 37(19): 6460-6469.
[28] Lu R K. Analytical Method of Soil Agro chemistry[M]. Beijing: Chinese Agricultural Science and Technology Press, 2000. 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社. 2000.
[29] Gong X J. The Response of Litter Decomposition to Carbon, Nitrogen and Phosphorus Addition for Four Typical Tree Species in Mid-subtropical Zone[D]. Nanchang: Nanchang University, 2011. 弓晓静. 中亚热带四种树种凋落叶分解对碳氮磷调控的响应[D]. 南昌: 南昌大学, 2011.
[30] Tu L H, Dai H Z, Hu T X,etal. Effect of simulated nitrogen deposition on litter decomposition in aBambusapervariabilis×Dendrocalamopsiplantation,Rainy Area of West China. Acta Ecologica Sinica, 2011, 31(5): 1547-1557. 涂利华, 戴洪忠, 胡庭兴, 等.模拟氮沉降对华西雨屏区撑绿杂交竹凋落物分解的影响.生态学报, 2011, 31(5): 1547-1557.
[31] He Y T, Qi Y C, Dong Y S,etal. Research review of the impact of exogenous nitrogen input on grassland soil microbial characteristics. Advances in earth science, 2010, 25(8): 877-885. 何亚婷, 齐玉春, 董云社, 等.外源氮输入对草地土壤微生物特性影响的研究进展. 地球科学进展, 2010, 25(8): 877-885.
[32] Yu L, Song X L, Zhao J N,etal. Effects of nutrient addition on plant diversity and productivity in aStipabaicalensisgrassland in Inner Mongolia, China. Acta ecologica sinica, 2015, 35(24): 8165-8173. 于丽, 宋晓龙, 赵建宁, 等.养分添加对贝加尔针茅草原植物多样性及生产力的影响. 生态学报, 2015, 35(24): 8165-8173.
[33] Meiliban M. The Effect of Nitrogen and Phosphorus Addition on Leaf Litter Decomposition ofSchimasuperbain Tiantong[D]. Shanghai: East China Normal University, 2014. 美丽班·马木提. 氮磷添加对天童木荷凋落叶分解的影响[D].上海: 华东师范大学, 2014.
[34] Li R H, Hu T X, Tu L H,etal. Effects of simulated nitrogen deposition on litter decomposition inNeosinocalamusaffinisstands in rainy area of West China. Chinese Journal of Applied Ecology, 2009, 20(11): 2588-2593. 李仁洪, 胡庭兴, 涂利华, 等.模拟氮沉降对华西雨屏区慈竹林凋落物分解的影响. 应用生态学报, 2009, 20(11): 2588-2593.
[35] Chen X. Effects of Simulated Nitrogen Deposition on Litter Decomposition of Nutrient Release of Dynamic inLarixgemliniiforest Study[D]. Hohhot: Inner Mongolia Agricultural University, 2014. 陈翔. 模拟氮沉降对兴安落叶松凋落物养分释放动态的影响研究[D].呼和浩特: 内蒙古农业大学, 2014.
[36] Chen H. Effect of N and P Addition on Foliar Litter Decomposition and Nutrient Release in a Subtropical Evergreen Broad-leaved Forest[D]. Hefei: Anhui Agricultural University, 2013. 陈昊.氮磷添加对亚热带北部常绿阔叶林落叶分解及其养分释放的影响[D]. 合肥: 安徽农业大学, 2013.
[37] Gong X J, Yu M Q, Hu X F,etal. Effects of nitrogen and phosphorus addition on leaf litter decomposition in a suburban slash pine plantation in red soil region of China. Chinese Journal of Ecology, 2010, 29(12): 2327-2333. 弓晓静, 余明泉, 胡小飞, 等.氮磷添加对红壤区城郊湿地松林凋落叶分解的影响. 生态学杂志, 2010, 29(12): 2327-2333.
[38] Chen Y, Wang F M, Mo Q F,etal. Effects of nitrogen and phosphorus additions on woody debris decomposition in a secondary tropical forest of South China. Chinese Journal of Application and Environmental Biology, 2015, 21(4): 747-753. 陈瑶, 王法明, 莫其锋, 等. 氮磷添加对华南热带森林尾叶桉木质残体分解和养分动态的影响. 应用与环境生物学报, 2015, 21(4): 747-753.
[39] Berg B, Matzer E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest ecosystems. Environmental Reviews, 1997, 5(1): 1-25.
[40] Berg B, Staaf H. Decomposition rate and chemical changes in decomposing needle litter of Scots pine Ⅱ. Influence of chemical composition. Ecological Bulletins-NFR, 1980, 32: 373-390.
[41] Gabriel P, Michaёl A, Thibaud D,etal. Home-field advantage: A matter of interaction between litter biochemistry and decomposer biota. Soil Biology & Biochemistry, 2013, 67: 245-254.
Effects of nitrogen and phosphorus addition on litter decomposition on theStipabaicalensissteppe
LI Wen-Ya1,2, QIU Xuan1, BAI Long2, YANG Dian-Lin1*
1.Agro-EnvironmentalProtectionInstitute,MinistryofAgriculture,Tianjin300191,China; 2.DepartmentofHorticulture,ShenyangAgriculturalUniversity,Shenyang110866,China
Litter decomposition is one of the main pathways of nutrient cycling and energy flow in grassland ecosystems. Nutrient limitation is the major factor limiting production on the warm steppe. In this study, the effects of nutrient addition on litter decomposition were investigated to provide a theoretical reference for nutrient management, restoration, and construction of grassland ecosystems. On theStipabaicalensissteppe in Inner Mongolia, the leaves, stems, and roots ofS.baicalensis(Sb),Leymuschinensis(Lc) andArtemisiafrigida(Af) were placed in mesh bags to measure decomposition in different treatments: CK (no fertilization), fertilization with N (ammonium nitrate), P (triple superphosphate), or NP (mixed ammonium nitrate and triple superphosphate). The residual rates of all three plants were significantly lower in the N, P, and NP treatments than in the CK, indicating that N and P addition promoted decomposition. During the decomposition process, N was released from leaves and roots at an early stage and enriched at later stages, whereas N in stems showed an enrichment-release-enrichment pattern. The leaves, stems, and roots released P and C continuously during decomposition. The residual rate of P decreased as the plant organs decomposed. The 95% decomposition time ofS.baicalensis,L.chinensis, andA.frigidawas 3.32-3.77 years, 2.95-3.96 years, and 2.64-4.89 years, respectively. The decomposition responses to fertilizers differed among plant organs and among plant species.
Stipabaicalensissteppe; litter; N, P addition; decomposition rate
10.11686/cyxb2016383
http://cyxb.lzu.edu.cn
2016-10-12;改回日期:2017-02-14
国家自然科学基金(31170435)资助。
李文亚(1989-),女,河北邯郸人,在读硕士。E-mail:724558548@qq.com*通信作者Corresponding author. E-mail:yangdianlin@caas.cn
李文亚, 邱璇, 白龙, 杨殿林. 氮、磷添加对贝加尔针茅草原凋落物分解的影响. 草业学报, 2017, 26(8): 43-53.
LI Wen-Ya, QIU Xuan, BAI Long, YANG Dian-Lin. Effects of nitrogen and phosphorus addition on litter decomposition on theStipabaicalensissteppe. Acta Prataculturae Sinica, 2017, 26(8): 43-53.
猜你喜欢
今日农业(2022年14期)2022-11-10 00:27:51
现代畜牧科技(2021年7期)2021-07-28 06:40:30
现代畜牧科技(2021年4期)2021-07-21 06:13:04
中外文摘(2018年17期)2018-09-08 02:45:04
环球时报(2018-08-29)2018-08-29 04:10:59
现代畜牧科技(2016年9期)2016-02-21 12:25:04
小哥白尼·趣味科学画报(2014年9期)2015-04-07 21:01:12
湖南农业科学(2014年14期)2014-02-27 14:31:52
环境工程技术学报(2011年4期)2011-09-05 09:54:46
对外经贸(2011年8期)2011-08-09 00:59:44
