
转基因猪育种中的小鼠模型
2017-08-18 01:54:02肖红卫郑新民
中国猪业 2017年7期
肖红卫 郑新民
(湖北省农业科学院畜牧兽医研究所/动物胚胎工程及分子育种湖北省重点实验室,湖北武汉430064)
转基因猪育种中的小鼠模型
肖红卫 郑新民*
(湖北省农业科学院畜牧兽医研究所/动物胚胎工程及分子育种湖北省重点实验室,湖北武汉430064)
本研究旨在研究转基因猪育种中的小鼠模型,探讨转基因小鼠传代中可能出现的情况,为转基因猪育种工作提供参考。选用昆明小白鼠制作转基因小鼠,然后进行转基因小鼠的传代,并对小鼠进行检测,统计转基因阳性小鼠的数量,研究转基因小鼠传代过程中阳性率的变化以及转基因个体的选择、表型分析、性状分析、育种规划以及育种中的线性模型分析。结果表明,转基因所得的首建阳性小鼠的比例为2.3%,转基因阳性小鼠与野生型交配传代时阳性后代的比例为50%左右;结果提示,原核显微注射法制作的转基因阳性个体在传代过程中,后代的比例不会超过50%。该结果将直接用于指导转基因猪的育种工作,即在转基因猪育种中要加强选择。
转基因猪;育种;小鼠模型
自从1908年Nilsson-Ehle提出微效多基因假说(multiple-factor hypothesis)[1]以及1941年Lush[2]提出遗传力概念后,育种学拥有了合理的内核。20世纪60年代到70年代,随着计算机的普及以及在动物育种上的应用,数量遗传学及其相应的育种学进入高速发展时期。然而正是由于计算机的应用使得育种规模越来越大,限制了其潜力的发挥。
转基因动物技术,开辟了在短期内直接改造生物遗传性的新天地,使以几十年为进化单位的常规育种缩短到几年,能将生物种属间不可逾越的鸿沟彻底填平,并可赋予群体特定的性状,因此在育种上具有巨大的潜能。
育种——原意是培育新品种,但是由于亲本枯竭,新品种培育理论研究陷于停滞,杂种优势的应用和理论研究发展缓慢,育种学家转而研究杂种优势利用,但进展缓慢,于是部分科学家把目光集中到转基因动物育种上来[3-6]。由于转基因猪的制作成本太大,转基因猪的育种工作具有长期性,因此,我们在启动转基因猪育种的同时,还开展以转基因小鼠为育种模型的研究,以期为转基因猪育种工作提供参考。
1 材料和方法
以前期所做9种基因(pOMT-PGH、hDAF、pBHSA、pSHSA、PT-HBV 1.3、Bcl-xL、hCD59、GFP、hCD59+MCP)的转基因小鼠传代数据为研究对象,研究转基因小鼠传代过程中阳性率的变化以及转基因个体的选择、表型分析、性状分析、育种规划以及育种中的线性模型分析。
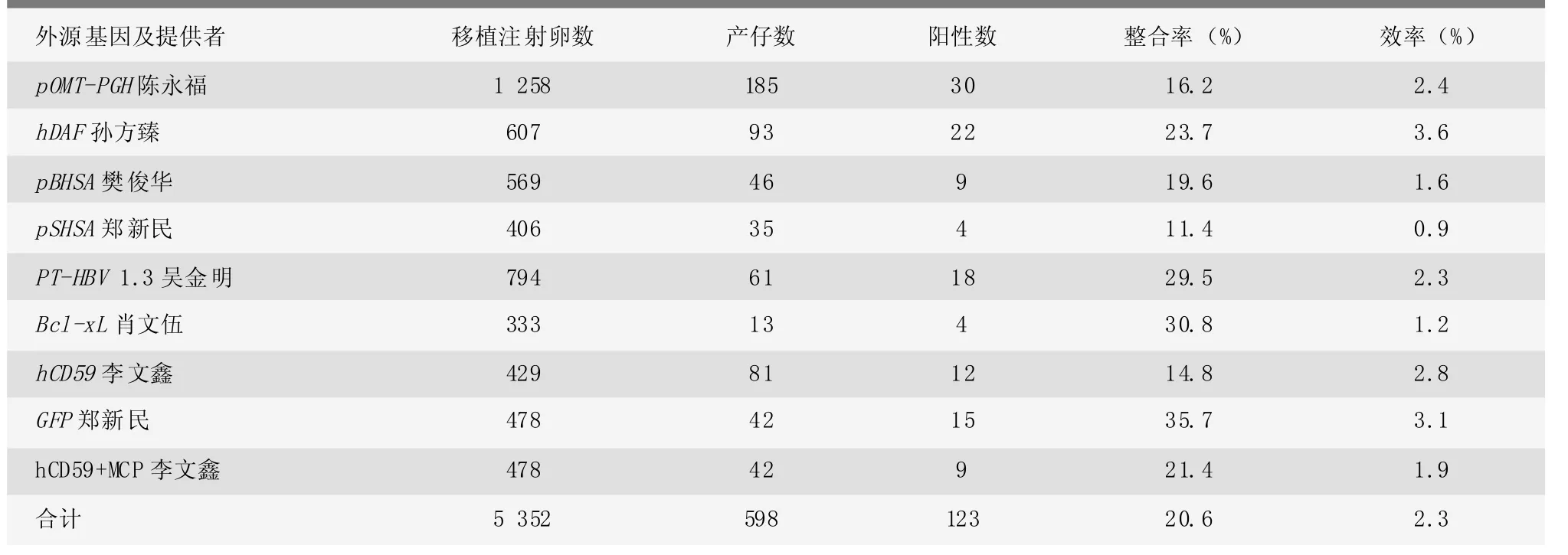
表1 转基因小鼠阳性率
2 结果及分析
2.1 原核显微注射所得后代阳性率的检测结果
结果见表1。
2.2 转基因小鼠的遗传规律
将4只原代转Bcl-xL基因小鼠分别与野生型小鼠交配传代[7],用PCR和Southern杂交确认后代中的转基因小鼠;后代中的阳性小鼠再与野生型小鼠交配传代。依此方式传至第五代。各世代转基因小鼠的比例如表2。
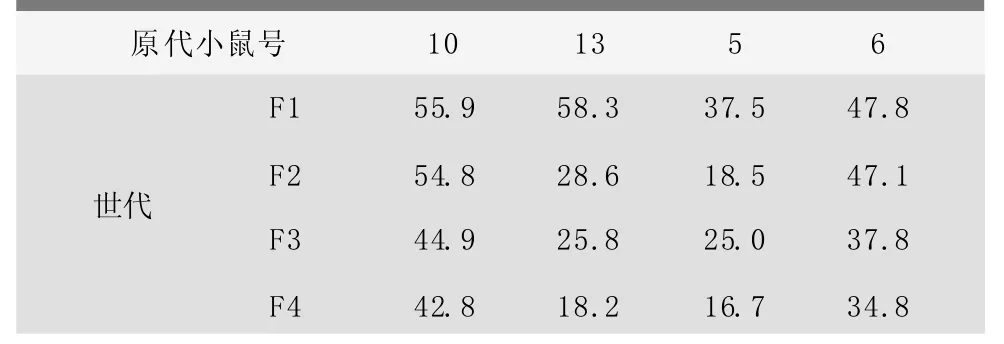
表2 转BcL-xL基因小鼠各世代的比例 (%)
10号阳性鼠经过选择后的后代,阳性率维持在50%左右,与孟德尔遗传的理论值相符,表明外源基因已经整合在小鼠两条染色体中的一条上,且遗传是稳定的;而其他3只小鼠,随着世代的增加,阳性率呈下降趋势,表明外源基因在传代中有丢失现象。
2.3 演化
Fisher曾证明,88.7%的新生基因在无任何压力影响下,只是随机影响,15代以内不可避免地消失。消失的极限概率为1,保留的极限概率为0[8]。因此,在转基因育种上,我们也要进行选择,否则所得到的转基因个体(突变体)也会消失。
演化的模型为:
G0代转基因纯合子个体第n代后代中的阳性个体比例为:

G0代转基因杂合子个体第n代后代中的阳性个体比例为:
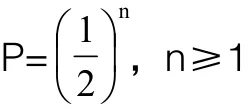
2.4 选择
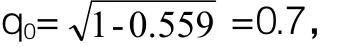
2.4.1 表型选择
采用表型选择的方法育成一个纯种群(p=99.96%即视为成功构建纯种群)需要的代数是:
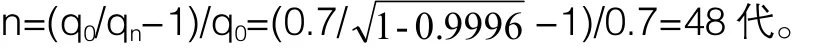
小鼠的世代间隔是9周龄(6周龄性成熟,妊娠期3周),育成一个纯种群需要经过49×(9/52)=8.5年。
2.4.2 测交法选择纯合子种畜留种的方法
采用测交法选择纯合子公畜留种的方法育成一个纯种群(p=99.96%即视为成功构建纯种群)需要的代数是:
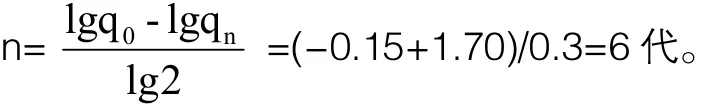
小鼠的世代间隔是9周龄(6周龄性成熟,妊娠期3周),育成一个纯种群至少需要经过6×(9/52)=1年。
2.4.3 染色体荧光原位杂交法选择纯合子种畜留种的方法
DDS产生的信号,经过放大才能在示波器上显示完整而稳定的波形。信号放大模块采用的是NE5532芯片[9]。其引脚图如图7所示。1引脚、7引脚为输出引脚;2引脚、6引脚为反相输入引脚;3引脚、5引脚是同相输入引脚;4引脚、8引脚分别是负电源和正电源引脚。
在获得转基因动物个体之后,用染色体荧光原位杂交法可以在一周内鉴定出所测个体是否为纯合子,节约了测交的时间。
综上,在选择时应该采用染色体荧光原位杂交法选择纯合子的方法,可以节约大量的培育时间。
2.5 表型值剖分
转基因研究中的目的是通过外源基因的表达从而使个体产生一个表型,如该表型为个体基因与外源基因所贡献,则研究人员需要对表型值进行剖分。
上述9种转基因小鼠研究的目的是通过外源基因的表达从而使小鼠产生一个有别于自身现有的表型,该表型全部为外源基因所贡献,因此,研究人员不需要对该表型值进行剖分。
2.5.1 基因型与群体均值的关系
小鼠获得的外源基因就是小鼠的基因型,其特点是表型决定于基因型,偏离群体均值以上的部分就是外源基因的贡献。
2.5.2 基因的平均效应
将外源基因以随机方式与群体的基因(或基因样本)相结合而形成的合子的平均基因型值对群体平均值的离差,就是该基因在特定群体中的平均效应。
2.5.3 基因取代的平均效应
如果以一个等位基因(A1)随机取代另一个等位基因(A2)所产生的基因型值变化,称为基因取代的平均效应。被取代的等位基因来自纯合子和杂合子的概率。
2.5.4 基因取代的平均效应与基因平均效应的直接转换
基因取代的平均效应是取代基因的平均效应与被取代基因频率之比,也是被取代基因平均效应与取代基因频率之比的负值。
2.5.5 关于育种值的群体遗传学分析
个体育种值是个体所有位点全部基因的平均效应之和,对于转基因动物而言,主要是考察外源基因的效应。育种值具备世代连续性:个体传递给后代的是基因而不是基因型,只有基因决定的加性效应即育种值是联系世代的唯一能够稳定遗传的成分。
2.6 来自转基因性状的遗传参数
由于大部分转基因动物中的目的性状是质量性状,因此,其遗传参数是0或者1。如果转基因动物的目的性状是数量性状,并且针对外源基因进行选择,那么其遗传参数还是0或者1。
2.7 育种规划
在转基因阳性动物中并不全部为纯合子,杂合子也表现为阳性,而且在杂合子的后代中会出现野生型的后代,因此在育种开始之前所用的亲本育种素材必须全部是纯合子[10]。
2.8 畜禽育种中的线性模型
常规育种所用的模型多能用数学方程式来表达,数学期望、方差以及协方差可以描述,可以假设及约束条件[11]。而转基因育种时首要选择的是质量性状即表达了外源基因的个体,淘汰没有表达外源基因的个体。在完成外源基因在染色体上的纯合后,可以借鉴常规育种中所用的线性模型。
3 讨论
当今的动物育种科学已经从过去的表型育种、杂交育种进入到分子育种或基因工程育种的新阶段。转基因动物育种与传统的动物育种相比,因其在改良动物遗传特性、提高其经济性状等上面所需要的时间短,就成为动物分子育种学的重要研究内容之一。然而,转基因育种与常规育种存在巨大的分歧,并随着研究的深入逐渐显示出来。
3.1 常规育种
目前,在商品猪生产上利用的是猪的杂交优势,在育种上是培育有特色经济性状的品系,然后筛选配合力高的组合,杂种后代性状突出、体型又符合主流育种界认可就可以推向市场[3]。在经过几个世代之后,所用的品种会逐渐退化。
3.2 转基因育种
转基因猪育种工作既要考虑外源基因在后代的传递,又要考虑不同组合间的配合力。在早期的转基因动物研究中,我们发现外源基因的整合发生在二细胞胚胎后期,会产生嵌合体的现象,导致出现外源基因不能稳定遗传的现象。且G0代与野生型交配传代到F1代时所有个体中阳性个体的比例是50%或100%,F1代阳性个体与野生型交配传代到F2代时所有个体中阳性个体的比例是25%,F2代阳性个体与野生型交配传代到F3代时所有个体中阳性个体的比例是12.5%。即外源基因在G0代阳性个体后代中的比例是递减的,这个发现使转基因动物育种有别于常规育种,即在做转基因育种研究时必须先得到纯合的G0代转基因动物,之后还要考虑不同血统的转基因动物存在。
[1]Nilsson-Ehle H.Einige Ergebnisse von Krenzunger bei Hafer und Weizen[J].Bot.Notiser,1908:257-294.
[2]Lush JL.Intra-sire correlations or regressions of offspring on dam as a method of estimating heritability of characteristics[J]. Proc Am Soc Anim Prod,1940,33:293-301.
[3]刘震乙.家畜育种学[M].北京:中国农业出版社,1981: 66-77.
[4]钟金成,陈智华.分子遗传与动物育种[M].成都:四川大学出版社,2001:288-295.
[5]肖红卫,华文君,张立苹,等,转基因动物技术及其在动物育种中的应用[J].中国猪业,2016(4):57-59.
[6]徐奎,程英,周荣,等,转基因技术,猪品种改良的新希望[J].中国猪业,2016(4):67-68.
[7]王芙蓉,袁慧,肖文伍,等.人bcl2xl转基因小鼠外源基因的复制及传代的稳定性观察[J].华中科技大学学报(医学版),2002,31 (6):615-617.
[8]Fisher RA.The correlation between relatives on the supposition of Mendelian inheritance[J].Trans.Roy.Soc.Edinburgh, 1918,52:399-433.
[9]张沅.家畜育种学[M].北京:中国农业出版社,2001: 111-122.
[10]门正明.动物遗传学[M].兰州:兰州大学出版社,1999: 223.
[11]张沅,张勤.畜禽育种中的线性模型[M].北京:中国农业大学出版社,1993:72-86.
Mice Model in Transgenic Pig Breeding
XIAO HongweiZHENG Xinmin
(Institute of Animal Husbandry and Veterinary/Hubei Key Laboratory of Animal Embryo and Molecular Breeding,Hubei Academy of A-griculture Sciences,Wuhan 430064,China)
This study was focused on the transgenic mouse model for transgenic pig breeding and to explore the possible occurrence of transgenic mice which will provide reference for the transgenic pig breeding.The transgenic mice were made from Kunming mice,and the transgenic mice were passaged.The positive offspring number of transgenic mice,the change of positive rate in the passage of transgenic mice,the selection of individual gene, phenotype analysis,character analysis,breeding program and linear model analysis in breeding were discussed. The results showed that the G0 transgenic mice is 2.3%and the positive offspring by mating G0 transgenic mice with wild type mice were about 50%.The results suggested that the proportion of offspring during passage of transgenic positive individuals by pronuclear microinjection was no more than 50%.The results will be used directly to guide the breeding of transgenic pigs and to strengthen selection in transgenic pig breeding.
Transgenic pig;breeding;mice model
*通讯作者:郑新民(1960-),男,汉族,陕西渭南人,研究员,主要从事动物生物技术研究,E-mail:anbit20@163.com
S813.2
1673-4645(2017)07-0044-04
2017-06-08
转基因生物新品种培育重大专项(2016ZX08010003-006),湖北省农业科技创新中心(2017-620-004-001),湖北省农业科学院青年基金(2016NKYJJ19),湖北省农业科学院竞争性项目(2016jzxjh013),湖北省技术创新专项重大项目(2016ABA117)
肖红卫(1979-),男,汉族,江苏盐城人,硕士,助理研究员,主要从事转基因猪遗传繁育研究,E-mail:xiaohongwei2003@163.com
猜你喜欢
郑州大学学报(医学版)(2023年5期)2023-09-19 01:49:48
舰船科学技术(2022年11期)2022-07-15 07:51:56
中国乳品工业(2022年4期)2022-05-19 07:10:14
福建畜牧兽医(2021年5期)2021-10-03 07:54:44
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
科学与财富(2016年32期)2017-03-04 20:07:39
考试周刊(2015年53期)2015-09-10 20:42:28
中学生物学(2014年1期)2014-05-19 15:10:06
