
3种棕榈藤叶片叶绿素荧光特征分析研究
2017-08-17 09:29杨意宏李利超孙化雨赵韩生高志民
世界竹藤通讯 2017年4期
杨意宏 李利超 孙化雨 赵韩生 高志民
(1 国际竹藤中心竹藤资源基因科学研究所,国家林业局竹藤科学与技术重点开放实验室 北京 100102;2 河北农业大学园艺学院 河北保定 071001)
3种棕榈藤叶片叶绿素荧光特征分析研究
杨意宏1,2李利超1孙化雨1赵韩生1高志民1
(1 国际竹藤中心竹藤资源基因科学研究所,国家林业局竹藤科学与技术重点开放实验室 北京 100102;2 河北农业大学园艺学院 河北保定 071001)
棕榈藤作为热带、亚热带植物中重要的森林资源,优质的藤材是重要的加工利用材料,具有重要的经济价值。叶片光合能力对藤材的形成具有重要影响,利用叶绿素荧光仪测定黄藤(Daemonoropsjenkinsiana)、大白藤(Calamusfaberii)、小白藤(C.balansaeanus)的叶绿素荧光参数,为研究逆境条件下棕榈藤的光合能力和选择适宜的栽培条件提供参考。结果表明,在实验室条件下3种棕榈藤的光系统Ⅰ(PSⅠ)实际光量子效率Y(Ⅰ)为小白藤>大白藤>黄藤,光系统Ⅱ(PSⅡ)的Y(Ⅱ)为大白藤>黄藤>小白藤;非光化学猝灭系数qN由高到低依次为大白藤、黄藤和小白藤;而光化学猝灭系数qP则是小白藤最高,黄藤次之,大白藤最低;PSⅠ的电子传递效率ETR(Ⅰ)是小白藤>大白藤>黄藤,而PSⅡ的ETR(Ⅱ)值则是小白藤>黄藤>大白藤;PSⅡ最大光量子产量Fv/Fm维持在0.78~0.8范围内,且大白藤显著高于黄藤和小白藤(P<0.05)。由此可见,在实验室条件下小白藤的光合效率在这3种藤中最高,其次为黄藤,大白藤最低;且光保护能力为大白藤最高,黄藤次之,小白藤最低。
棕榈藤;黄藤、大白藤、小白藤;叶绿素荧光;光合作用
棕榈藤为棕榈科(Palmae)植物,是重要的热带雨林植物类群。棕榈藤有13个属600余种,主要分布在亚洲热带地区[1-2]。棕榈藤因具有优良的加工工艺特性在编织工业上得到广泛应用,早在20世纪70年代棕榈藤创造的市场价值就已达到70亿美元[3]。因此,关于棕榈藤的开发利用倍受关注。在棕榈藤的天然分布[4]、引种驯化[5]、离体快繁[6-11]、良种选育与育苗[12-13]、栽培技术[14-16]、种植造林[17-18]以及藤材特性与加工利用[19-22]等方面,已有大量研究报道。然而,由于棕榈藤的攀援、伴生、多刺等特性,使得棕榈藤在资源培育、采收等方面均受到限制,相关的基础研究进展缓慢,尤其是在棕榈藤光合作用研究方面更鲜有报道。
光是影响植物生长的重要因子,光合作用产物的积累是藤材形成的源泉,因此藤种的光合特性对藤材的形成具有重要影响。研究表明[23-24],不同藤种的光合特性存在着一定的差异,如盈江省藤(Calamusnambariensisvar.yingjiangensis)和高地省藤(C.nambariensisvar.alpinus)的净光合速率(Pn)和蒸腾速率(Tr)日变化均呈“双峰”曲线,存在明显的“午休”现象,而小省藤(C.gracilis)的Pn日变化为“双峰”曲线,Tr日变化为“单峰”曲线,环境因子对棕榈藤的光合速率和蒸腾速率均有显著影响。植物吸收光后除了满足光合反应外,还能以热能和荧光的形式散失,尤其是在逆境条件下更为明显。叶绿素荧光参数能快速、有效地反映植物的光合能力和生长情况[25],但目前尚未有关于棕榈藤叶绿素荧光的研究报道。棕榈藤叶片为羽状复叶,羽片叶渐尖,具刚毛,有数条纵向叶脉,中(主)脉较粗,叶脉和羽片边缘具刺或刚毛[26],属于大型热带雨林的攀援性林木,测定野外环境下棕榈藤的光合参数有难度,因此选用实验室环境下的棕榈藤作为研究对象。本研究利用叶绿素荧光仪测定省藤属的大白藤(C.faberii)、小白藤(C.balansaeanus)和黄藤属的黄藤(Daemonoropsjenkinsiana)叶片光系统Ⅰ(PSⅠ)和光系统Ⅱ(PSⅡ)的叶绿素荧光参数,通过比较分析,探讨3种棕榈藤的光合能力的差异,为棕榈藤的栽培条件选择提供参考依据。
1 材料与方法
1.1 实验材料及培养条件
选用2年生省藤属大白藤、小白藤和黄藤属黄藤实生苗为研究对象,在腐殖质土和蛭石(7∶3)基质中进行盆栽,气温为23 ℃,空气相对湿度为65%,光照约为200 μmol/(m2·s),光照周期为光照16 h/黑暗8 h。
1.2 测定方法
1.2.1 叶绿素荧光参数测定
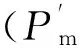
1.2.2 数据分析
使用Microsoft Office Excel 2013软件和SPSS Statistics 19数据处理系统进行数据的统计分析。
2 结果与分析
2.1PSⅠ荧光动力学参数
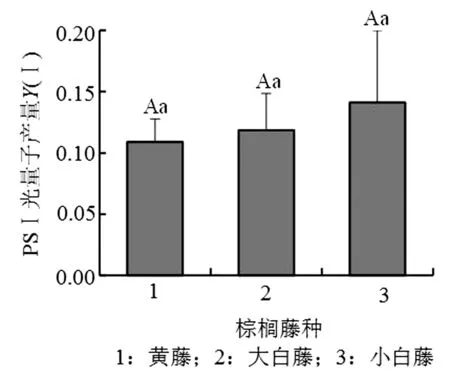
图1 PSⅠ荧光参数Y(Ⅰ)
光照条件下PSⅠ反应中心能吸收700 nm波长的光子并被其激发。利用P700远红光测定3种棕榈藤的PSⅠ实际光合速率Y(Ⅰ)。结果显示,小白藤最高,大白藤次之,黄藤最低(图1),且三者之间无明显差异;3种棕榈藤PSⅠ的非光化学能量转换的量子产量Y(NA)和光化学能量转换的量子产量Y(ND)值分别维持在0.1~0.2和0.7~0.8范围内,由高到低依次分别为大白藤>黄藤>小白藤、黄藤>大白藤>小白藤(表1)。
黑暗处理的棕榈藤接受饱和脉冲光后,相对电子传递效率ETR能在短时间内快速上升并达到顶
峰,此时大白藤的电子传递效率高于黄藤和小白藤,随后3种棕榈藤的ETR值开始下降,当测定时间延伸到200 ms后,棕榈藤的传递速率变化趋于平稳状态,直至最后3种棕榈藤ETR值由高到低依次为小白藤>大白藤>黄藤(图2)。
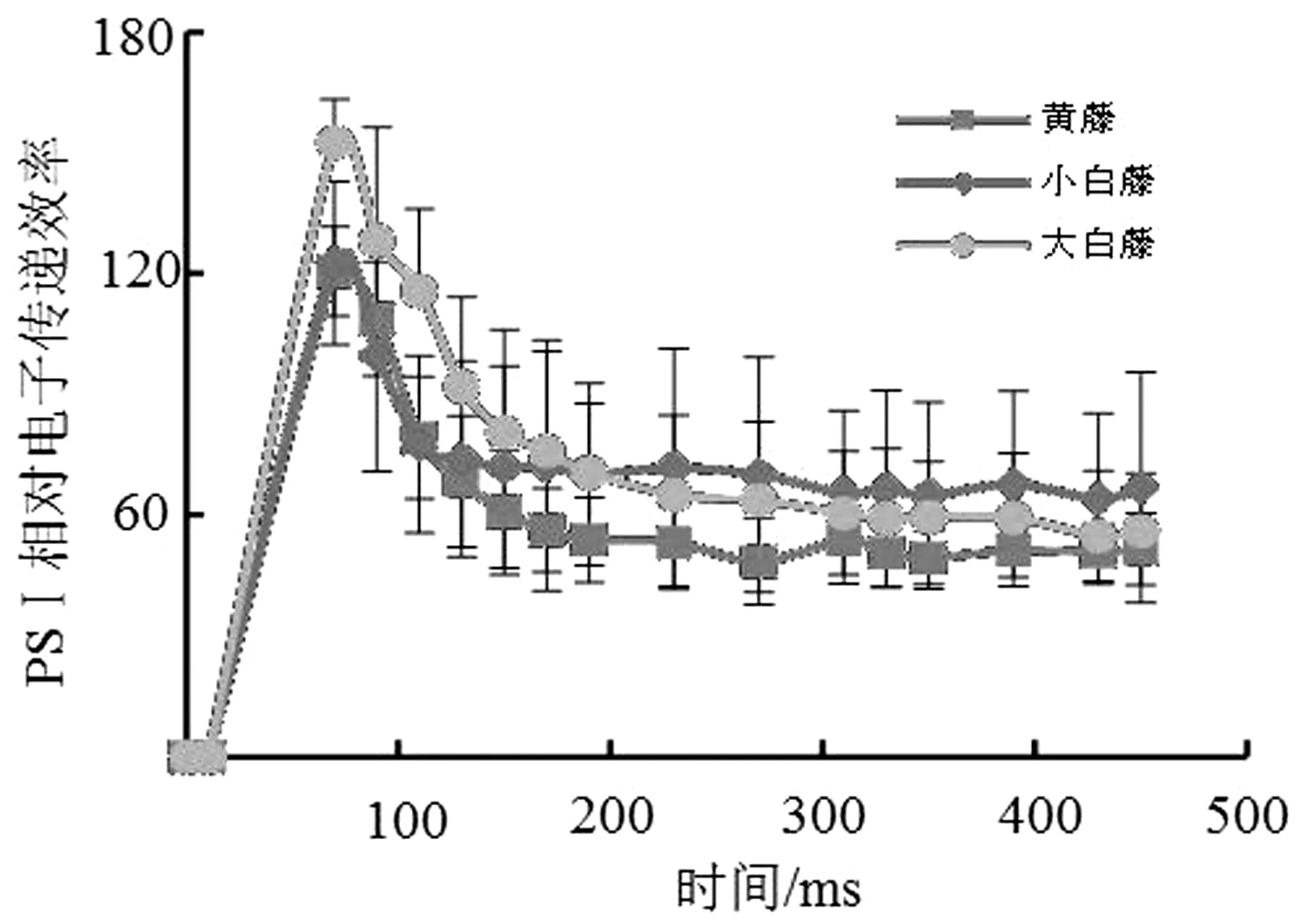
图2 PSⅠ荧光参数ETR变化
藤种Y(ND)Y(NA)FoY(NO)Y(II)黄藤0.755±0.039(Aa)0.136±0.024(Aa)0.297±0.029(Aa)0.278±0.009(Aa)0.039±0.010(Aa)大白藤0.732±0.044(Aa)0.150±0.035(Aa)0.295±0.043(Aa)0.295±0.025(Aa)0.024±0.005(Bb)小白藤0.726±0.109(Aa)0.134±0.056(Aa)0.325±0.038(Aa)0.250±0.033(Bb)0.044±0.013(Aa)
注:相同小写字母代表差异不显著,不同小写字母代表差异显著(P<0.05);相同大写字母代表差异不显著,不同大写字母代表差异极显著(P<0.01),下同。
2.2PSⅡ荧光动力学参数
PSⅡ最大光化学量子产量Fv/Fm能够反映植物的潜在光合能力[29-30]。3种棕榈藤Fv/Fm为0.78~0.8,其中以大白藤的数值最高,小白藤和黄藤无显著差异,但大白藤Fv/Fm值显著高于另外2种藤(P<0.05)(图3)。测定3种棕榈藤的初始荧光值Fo维持在0.3左右,小白藤最高,黄藤和大白藤相差无几;衡量PSⅡ量子产量的指标还包括非调节性能量耗散的量子产量Y(NO),其值由高到低为大白藤>黄藤>小白藤;PSⅡ实际光合效率Y(Ⅱ)能作为衡量植物叶片电子传递效率快慢的指标,小白藤和黄藤的Y(Ⅱ)平均值约为0.04,显著高于大白藤(表1)。
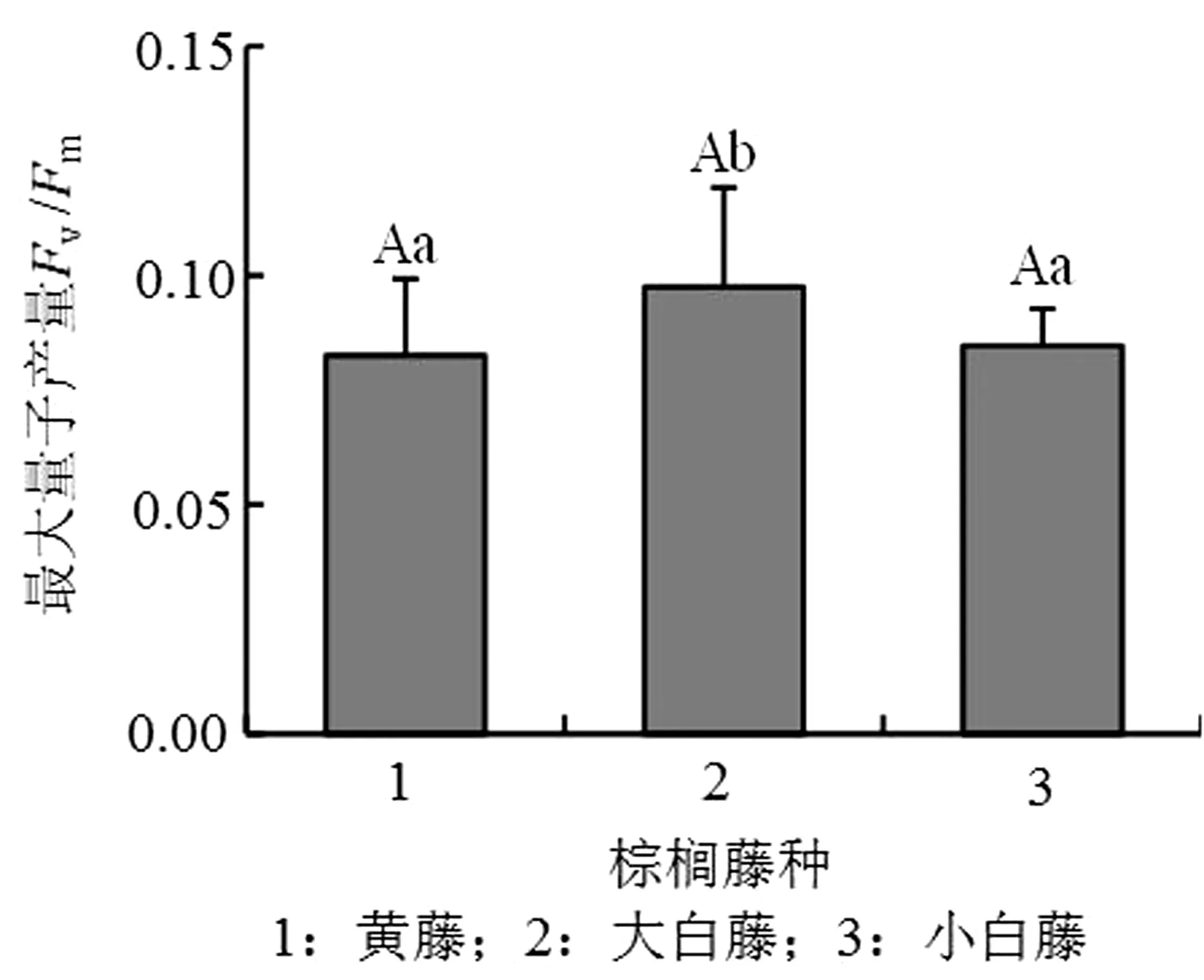
图3 PSⅡ荧光参数Fv / Fm
PSⅡ光化学反应过程中的电子传递相对效率ETR在3种棕榈藤间的差异较为明显,黑暗条件下3种棕榈藤的ETR均为0,饱和脉冲光作用下的黄藤和小白藤迅速升高,大白藤在开始阶段缓慢上升,随即上升较快;120 ms左右黄藤达到最大值,随后电子传递效率变化趋于平缓,该阶段小白藤出现缓慢上升的趋势,而大白藤则在100 ms左右达到最大值后变化趋势无明显变化,最后3种棕榈藤PSⅡ的ETR值为小白藤>黄藤>大白藤(图4)。
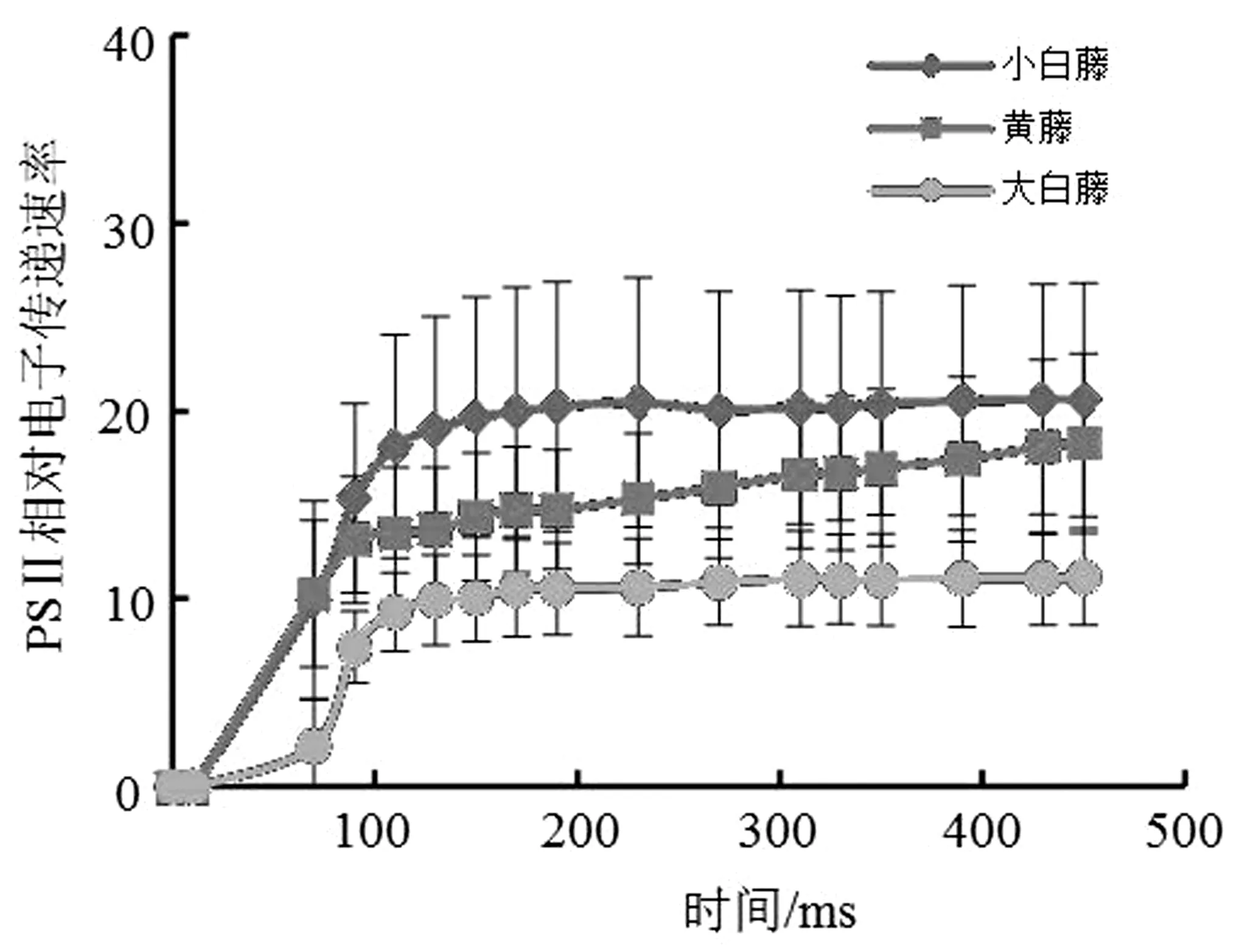
图4 PSⅡ荧光参数ETR变化
光化学系数(qP)反映植物光合活性的高低,非光化学系数(qN)则反映植物耗散过剩光能为热的能力,即光保护能力。3种棕榈藤光化学系数(qP)的数值在0.8左右,小白藤最高,黄藤次之,大白藤最低,黄藤和大白藤之间无明显差异,且均显著低于小白藤(P<0.01)(图5)。非光化学系数qN由高到低依次为小白藤、黄藤、大白藤,黄藤和小白藤无明显差异,与大白藤间呈极显著关系(P<0.01)(图6)。
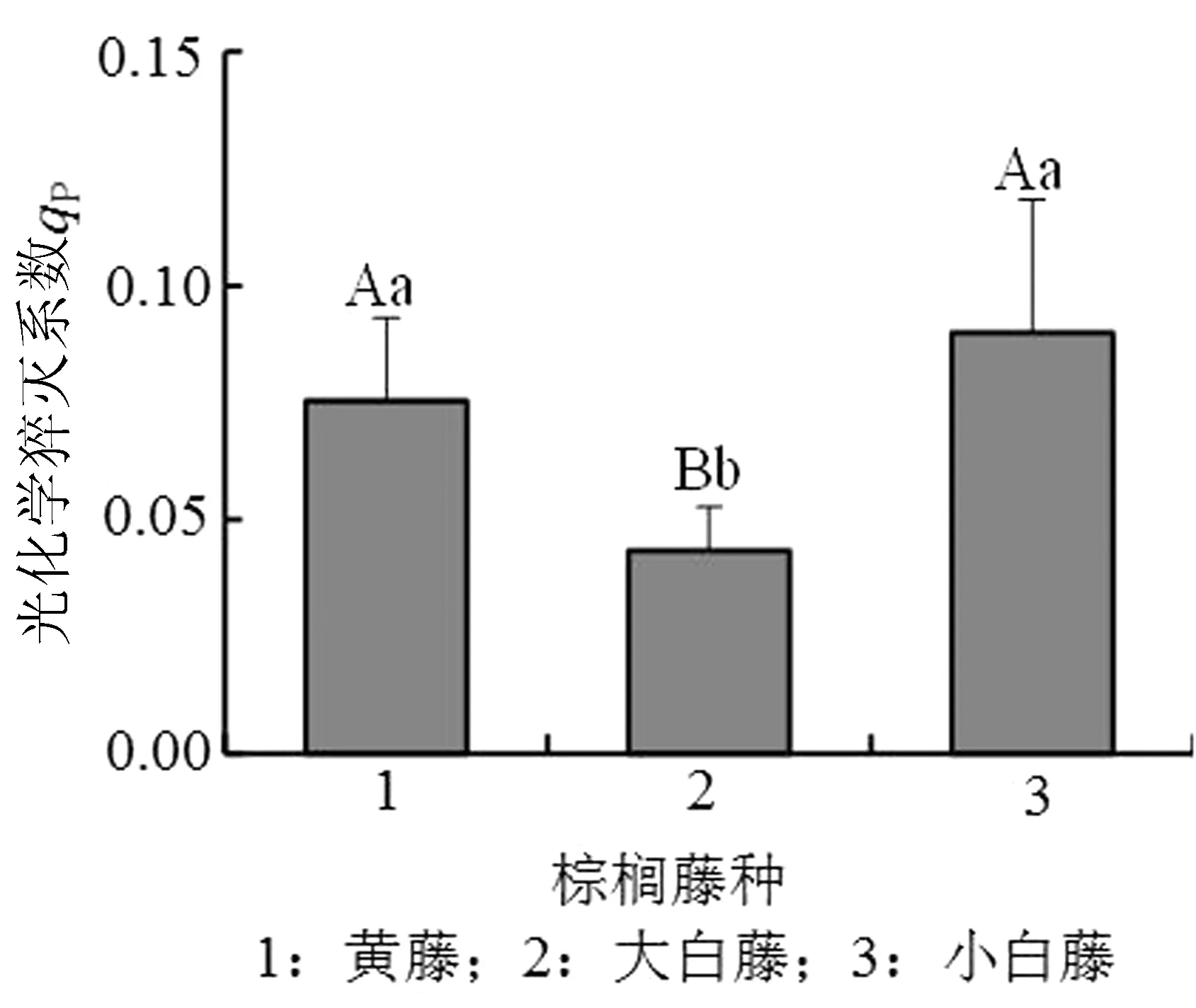
图5 PSⅡ荧光参数qP
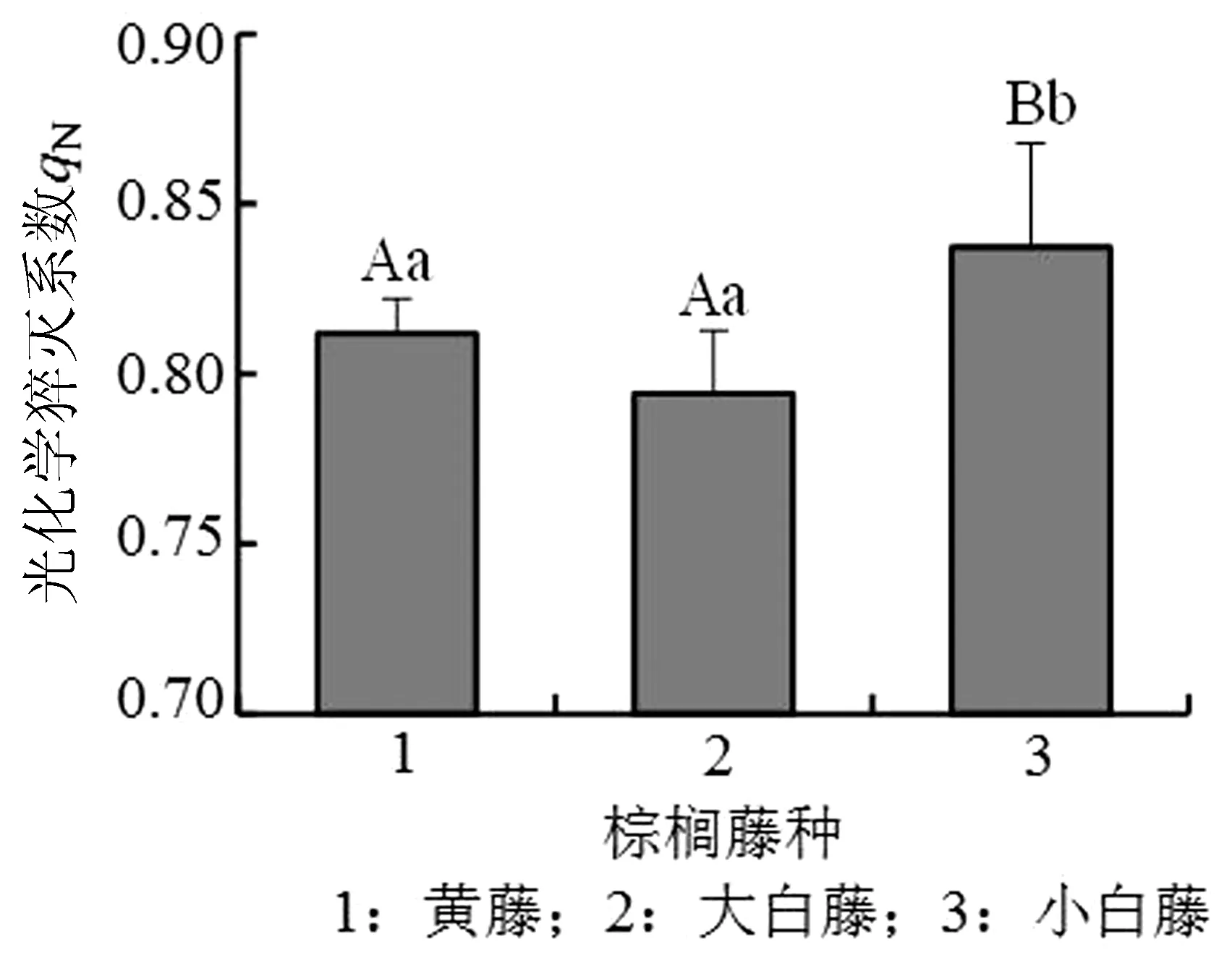
图6 PSⅡ荧光参数qN
3 小结与讨论
全球环境变化使得越来越多的植物生长发育均不同程度地受到逆境的影响,其中逆境对植物光合作用的影响尤为明显。叶绿素荧光参数是反映植物PSⅠ和PSⅡ受逆境胁迫状况的有效生理指标,通过它可较为直观快速地了解植物的光合能力,同时又可避免对植物造成伤害,是目前研究植物逆境条件下光合作用的有效手段[31]。Fv/Fm(一般为0.8)与植物PSⅡ的捕光色素复合体传递光能的效率相关,Fv/Fm越高,植物PSⅡ传递光能的潜在效率越高[32]。本研究叶绿素荧光参数测定结果表明,3种棕榈藤的Fv/Fm均在0.8以下,其中大白藤的最高,说明3种棕榈藤在实验室环境下均处于逆境,其中大白藤的胁迫程度最低,而黄藤和小白藤受胁迫程度类似;且在该条件下的大白藤PSⅡ传递效率和能量转化效率最高。
已有研究表明,植物的光合效率越高,Y(Ⅱ)和qP的数值越高,分析可知3种棕榈藤的光合效率能力依次为:小白藤>黄藤>大白藤,且反映植物PSⅡ的电子传递速率ETR值由高到低依次为小白藤>黄藤>大白藤。PSⅠ光量子产量Y(Ⅰ)依次为小白藤>大白藤>黄藤,PSⅠ的电子传递速率ETR(Ⅰ)值由高到低依次为小白藤>大白藤>黄藤,说明小白藤PSⅠ的光合效率大于黄藤和大白藤。反映植物光保护能力的非光化学猝灭系数qN由高到低为大白藤、黄藤和小白藤,Y(NO)和Y(NA)能反映植物在逆境下的受胁迫程度,分析得知大白藤的光保护能力最强,黄藤次之,小白藤最弱。由此可见,小白藤的PSⅡ光合活性最大,黄藤次之,大白藤最低。F0与热耗散呈正相关,由该值的变化情况可推测植物的光合作用反应中心情况[33]。分析可知,小白藤F0值最高,黄藤次之,大白藤最低,说明3种棕榈藤的热耗散程度依次为小白藤>黄藤>大白藤。研究结果将有助于棕榈藤栽培环境条件的选择与控制,对培育棕榈藤良种壮苗具有重要价值。
[1] Johnson D, Sunderland T. Rattan glossaryand compendium glossary with emphasis on Africa[M]. Rome: Food and Agriculture Organization,2004.
[2] 江泽慧. 世界竹藤[M]. 沈阳: 辽宁科学技术出版社, 2002.
[3] 蔡则谟, 许煌灿, 尹光天, 等. 棕榈藤利用的研究与进展[J]. 林业科学研究, 2003, 16(4): 479-487.
[4] 许煌灿, 尹光天, 李意德, 等. 我国棕榈藤天然分布及其利用研究[J]. 林业科学研究, 1993, 6(4): 380-389.
[5] 李荣生, 许煌灿, 李双忠. 世界棕榈藤引种驯化进展[J]. 世界林业研究, 2002, 15(2): 35-41.
[6] 李荣生, 许煌灿, 尹光天, 等. 中国棕榈藤引种进展[J]. 世界林业研究, 2003, 16(2): 48-54.
[7] Umail-Garcia M. Tissue culture of some rattan species[M]//Proceeding of the Rattan Semiar. Kuala Lumpur, Malaysia, 1985: 23-31.
[8] 张方秋. 棕榈藤组培技术研究[J]. 林业科学研究, 1993, 6(5): 486-492.
[9] 曾炳山, 许煌灿, 刘英, 等. 棕榈藤组培苗移植技术研究[J]. 林业科学研究, 1997, 10(6): 563-569.
[10] 曾炳山, 许煌灿, 刘英, 等. 棕榈藤组培苗根的诱导研究[J]. 林业科学研究, 1998, 11(3): 260-264.
[11] 高志民, 范少辉, 蔡春菊, 等. 世界棕榈藤离体快速繁殖研究与发展趋势[J]. 世界竹藤通讯, 2005, 3(3): 1-4.
[12] 杨华. 黄藤和单叶省藤遗传多样性与早期性别鉴定的研究[D]. 北京: 中国林业科学研究院, 2005.
[13] 李荣生, 尹光天, 杨锦昌, 等. 乡土用材棕榈藤种半同胞家系的早期选择[J]. 世界竹藤通讯, 2009, 7(4): 1-5.
[14] Yin G, Xu H. Effect of fertilizers on seedlings ofDaemonoropsmargaritaecommercial rattan species[J].Ric Bulletin, 1992,11(1): 9-12.
[15] 陈康泰. 棕榈藤的科研进展及人工栽培技术研究[J]. 热带作物科技, 1993(2): 23-27.
[16] 张昌顺, 范少辉, 官凤英. 中国棕榈藤培育与利用中的问题及对策[J]. 世界竹藤通讯, 2009, 7(4): 1-4.
[17] 许煌灿, 尹光天, 曾炳山, 等. 黄藤栽培技术研究[J]. 林业科学研究, 1994, 7(2): 239-246.
[18] 杨锦昌. 单叶省藤和黄藤人工林的系统经营技术[D]. 北京: 中国林业科学研究院, 2004.
[19] 蔡则谟. 棕榈藤茎的解剖特性及商用藤归类[J]. 林业科学, 1994, 30(3): 209-213.
[20] 吴玉章. 3种棕榈藤藤材的化学组成[J]. 林业科学研究, 2007, 43(67): 155-158.
[21] 王传贵, 徐鑫, 汪佑宏, 等. 黄藤和单叶省藤的主要物理力学性质[J]. 东北林业大学学报, 2011, 39(12): 132-133,136.
[22] 尚莉莉. 钩叶藤材的基本性能及增强增韧改性研究[D]. 北京: 中国林业科学研究院, 2014.
[23] 官凤英, 范少辉, 刘亚迪, 等. 两种棕榈藤光合日变化及其与环境因子的关系研究[J]. 世界竹藤通讯, 2010, 8(4): 1-6.
[24] 官凤英, 范少辉, 刘亚迪, 等. 高地省藤幼苗光合作用日变化特征[J]. 浙江林业科技, 2010, 30(5): 33-37.
[25] 郭春芳, 孙云. 叶绿素荧光动力学在植物抗性生理研究中的应用[J]. 福建教育学院学报, 2006(7): 120-123.
[26] 江泽慧, 王慷林. 中国棕榈藤[M]. 北京: 科学出版社, 2013.
[27] Demmig-Adams B, Adams W W I. Chlorophyll and carotenoid composition in leaves ofEuonymuskiautschovicusacclimated to different degrees of light stress in the field[J]. Functional Plant Biology, 1996, 23(5): 649-659.
[28] Kramer D M, Johnson G, Kiirats O, et al. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes[J]. Photosynthesis Research. 2004, 79(2): 209-218.
[29] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学报, 1999, 16(4): 444-448.
[30] 孙化雨, 娄永峰, 赵韩生, 等. 开花麻竹叶绿素荧光参数测定[J]. 世界竹藤通讯, 2014, 12(3): 12-16.
[31] 吕芳德, 徐德聪, 侯红波, 等. 5种红山茶叶绿素荧光特性的比较研究[J]. 经济林研究, 2003, 21(4): 4-7.
[32] 王可玢, 许春辉, 赵福洪, 等. 水分胁迫对小麦旗叶某些体内叶绿素a荧光参数的影响[J]. 生物物理学报, 1997, 13(2): 273-278.
[33] 冯建灿, 胡秀丽, 毛训甲. 叶绿素荧光动力学在研究植物逆境生理中的应用[J]. 经济林研究, 2002(4): 14-18.
A Study on Chlorophyll Fluorescence Characteristics of Three Rattan Species
Yang Yihong1,2Li Lichao1Sun Huayu1Zhao Hansheng1Gao Zhimin1
(1 State Forestry Administration Key Open Laboratory on the Science and Technology of Bamboo and Rattan,Institute of Gene Science for Bamboo and Rattan Resources, International Center for Bamboo and Rattan, Beijing 100102, China;2 College of Horticulture, AgriculturalUniversity of Hebei, Baoding 071001, China)
As the important forest resource of tropical and subtropical plants, high quality rattans are important materials for processing and utilization which have great economic value. Leaf photosynthetic capacity has an important effect on the formation of rattan. The chlorophyll fluorescence parameters ofDaemonoropsjenkinsiana,CalamusfaberiiandC.balansaeanuswere measured using chlorophyll fluorescence spectrometer, which will provide reference for the study on photosynthetic capacity of rattan and the selection of appropriate cultivation conditions. The result indicated that under laboratory conditions the value of photochemical efficiencyY(Ⅰ) inC.balansaeanuswas the highest one, followed by those inC.faberiiandD.jenkinsiana, and the value ofY(Ⅱ) inC.faberiiwas the highest one, followed by those inD.jenkinsianaandC.balansaeanus. The non-photochemical quenching coefficient (qN) ranked from high to low was inC.faberii,D.jenkinsianaandC.balansaeanus, and the photochemical quenching coefficient (qP) inC.balansaeanuswais the highest one, followed by those inD.jenkinsianaandC.faberii. Besides, the electron transport rate (ETR) of PSⅠranked from high to low was inC.balansaeanus,C.faberiiandD.jenkinsiana, while that of PSⅡinC.faberiiwas the highest one, followed by those inD.jenkinsianaandC.balansaeanus. The average values ofFv/Fmof three rattan species ranged from 0.78 to 0.8, in which the value ofFv/FminC.faberiiwas obviously higher than those in other rattan species(P<0.01). Therefore, it could be concluded that under the laboratory conditions the photosynthetic efficiency ofC.balansaeanuswas the highest, followed byD.jenkinsianaandC.faberii, andC.faberiihad the highest ability of photoprotection, followed byD.jenkinsianaandC.balansaeanus.
rattan,Daemonoropsjenkinsiana,Calamusfaberii,C.balansaeanus, chlorophyll fluorescence characteristics
“十二五”农村领域国家科技计划项目“竹藤种质资源创新利用研究”第一课题“竹藤优异种质创制创新与种苗培育标准化示范”(编号:
杨意宏(1993-),女,在读硕士生,从事竹藤花卉分子育种方面的研究。
高志民,男,研究员,博士生导师,研究方向为竹藤生长发育的分子基础。E-mail: gaozhimin@icbr.ac.cn。
10.13640/j.cnki.wbr.2017.04.004
2015BAD04B01)。
猜你喜欢
世界竹藤通讯(2022年2期)2022-05-07
世界竹藤通讯(2020年6期)2021-01-08
小天使·一年级语数英综合(2020年11期)2020-12-16
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
今日农业(2019年16期)2019-01-03
江西建材(2018年7期)2018-02-14
文理导航·科普童话(2017年4期)2018-02-10
散文诗(2017年17期)2018-01-31
生物学教学(2017年9期)2017-08-20
