
柑橘衰退病毒株对两种橘蚜不同翅型取食行为的影响
2017-08-09 02:35:32赵如娜何应琴鲁卓越陈文龙杨毓银王雪峰周常勇
山地农业生物学报 2017年4期
赵如娜,何应琴,鲁卓越,陈文龙,杨毓银,王雪峰,周常勇
(1.贵州大学 昆虫研究所/贵州省山地农业病虫害重点实验室/农业部贵阳作物有害生物科学观测实验站,贵州 贵阳 550025;2.中国农业科学院柑桔研究所,重庆 北碚 400712)
柑橘衰退病毒株对两种橘蚜不同翅型取食行为的影响
赵如娜1,何应琴1,鲁卓越1,陈文龙1,杨毓银1,王雪峰2,周常勇2
(1.贵州大学 昆虫研究所/贵州省山地农业病虫害重点实验室/农业部贵阳作物有害生物科学观测实验站,贵州 贵阳 550025;2.中国农业科学院柑桔研究所,重庆 北碚 400712)
为明确感染柑橘衰退病毒(Citrustristezavirus,CTV)的柑橘对不同翅型橘蚜取食行为的影响,通过刺探电位图谱技术(electrical penetration graph,EPG)对褐色橘蚜和棉蚜在感染CTV植株上的取食行为进行持续记录,与健康植株进行比较分析。结果表明,褐色橘蚜无翅型在感病植株的参数均与有翅型无显著差异;棉蚜无翅型在感病植株上的非刺探波(no penetration,np波)和路径波(C波)的总持续时间、电势落差(potential drops,pd波)的次数均显著小于有翅型,C波和刺探的次数均极显著小于棉蚜有翅型。褐色橘蚜无翅型在感病植株上的pd波的次数和韧皮部分泌唾液波(E1波)的平均持续时间显著小于棉蚜无翅型;褐色橘蚜有翅型在感病植株上的C波的次数和总持续时间、刺探的次数、pd波的次数和总持续时间均极显著小于棉蚜有翅型,韧皮部被动吸食波(E2波)的总持续时间和平均持续时间均极显著大于棉蚜有翅型。褐色橘蚜无翅型在感病植株上的刺探和pd波的次数均显著高于健康植株,但总刺探时间相反,np波、C波和非韧皮部阶段的总持续时间极显著高于健康植株,E2波的总持续时间极显著短于健康植株。表明柑橘感病状态对无翅型和有翅型的影响是相似的;柑橘感染CTV对褐色橘蚜无翅型的取食行为影响明显,且偏好健康植株,从而促进病毒的传播。
刺吸电位图谱;柑橘衰退病;有翅蚜;无翅蚜;病毒传播
柑橘衰退病毒(Citrustristezavirus,CTV)隶属于长线性病毒科(Closteroviridae)长线性病毒属(Closterovirus),是一种正义单链RNA病毒[1,2],广泛分布于世界各柑橘产地,对全世界的柑橘产业造成了严重威胁。在自然条件下,CTV在田间是通过褐色橘蚜(Toxopteracitricida(Kirkaldy))、棉蚜(AphisgossypiiGlover)、绣线菊蚜(AphiscitricolaVan der Goot)和橘二叉蚜(Toxopteraaurantii(Boyer de Fonscolombe))等传播。一般情况下棉蚜为CTV的有效传播介体,橘蚜为CTV的最有效传播介体。在长期的进化过程中,媒介昆虫、病毒和寄主之间形成了复杂的互作关系。病毒感染植物后会引起生理变化,从而增加植物的吸引力和偏好性[3,4]。
感染番茄斑萎病毒(Tomatospottedwiltvirus,TSWV)的植株比健康植株更能吸引西花蓟马Frankliniellaoccidentalis(Pergande)的取食,并且对于未携带病毒的雌性西花蓟马更喜欢在带毒植株上取食和产卵[5,6]。Jiménez-Martínez等[7]研究结果表明,小麦在感染大麦黄矮病毒(Barleyyellowdwarfvirus,BYDV)后相比于健康植株会产生很多挥发物质来吸引禾谷缢管蚜Rhopalosiphumpadi(L.)。在植株-病毒-昆虫之间的关系,病毒也有出现相反的作用。Fiebig等[8]研究结果表明,小麦在感染大麦黄矮病毒后叶片韧皮部总氨基酸的浓度降低,导致麦长管蚜(Sitobionavenae(Fabricius))对植株的吸食效率降低。以上植物病毒对昆虫的影响与其取食行为密切相关,而刺探电位图谱(electrical penetration graph,EPG)是目前研究刺吸式口器昆虫刺吸行为的重要工具,它记录的波形恰好精确地反映了昆虫口针在植物组织内部的进行情况[9,10],已广泛应用于蚜虫、烟粉虱、叶蝉、蓟马、蝽等刺吸式昆虫取食行为的研究,在建立波形基础上可以用于刺吸式口器昆虫对寄主植物的选择性、昆虫传播植物病毒的机制和植物的抗虫机制以及内吸性农药的测定等行为生态学领域[11]。
蚜虫根据环境的不同,产生2种不同的生物型:扩散生物型(有翅蚜)和非扩散生物型(无翅蚜)。其中有翅蚜可以获取较远距离的食物资源,作为病毒病的重要传播介体,对田间作物病毒病的传播起着重要的作用。高伟民等[12]认为有翅蚜的迁入是无翅蚜发生为害和烟草黄瓜花叶病发病率上升的主要因素。并且有翅蚜第一次的迁入量对蚜传病毒病发生程度有显著正效应作用,对病毒的传播更重要[13]。有研究结果表明,与健康植株相比,褐色橘蚜无翅型更偏爱取食感染CTV的柑橘苗。然而有关CTV病株对有翅型蚜虫影响还不清楚,因此,本研究通过EPG技术比较褐色橘蚜、棉蚜有翅型和无翅型成蚜在健康柑橘植株与感染CTV植株上的取食行为差异,探究柑橘植株感染CTV后对其取食行为的影响,为明确病毒-寄主植物-昆虫三者之间的相互关系提供依据。
1 材料与方法
1.1 材料
1.1.1 供试植物 试验植物是温州蜜柑(Citrusunshiu),购自湖南邵阳和谐苗木专业合作社,3年生实生苗。CTV4毒源由中国农业科学院柑橘研究所国家柑橘苗木脱毒中心提供,采用皮接的方法于健康植株接种CTV4病毒,8个月后利用Trizol方法提取叶片总RNA,利用实时荧光定量[14]进行病毒检测,筛选出感染CTV4病毒植株。健康植株、染病植株在温度、光照、湿度的温室内培养,待长出嫩叶时用于实验。1.1.2 供试虫源 褐色橘蚜、棉蚜均采自贵州大学农场柑橘园,于健康嫩梢充足的柑橘苗上,于温度(白天25℃/夜晚20℃)、光照(16L∶8D)、相对湿度(60±10)%的人工气候室(江南仪器厂)内培养。饲养多代后产生有翅蚜,取大小和日龄一致的无翅成蚜、有翅成蚜用于实验,并在实验前饥饿处理1 h。1.2 方法
1.2.1 刺探电位波形记录 用可视直流刺探电位技术(DC-EPG,Giga-8)监测褐色橘蚜、棉蚜有翅型和无翅型分别在健康植株和染病植株上的取食行为。成蚜与柑橘苗分别连入生物电流放大器的昆虫电极和植物电极,昆虫电极是一段长2~3 cm、直径为15 μm的金丝,末端用水溶性导电银胶粘在蚜虫的前胸背板上,植物电极则直接插在柑橘苗生长的塑料花盆中。缚有金丝的成蚜饥饿1h后放在柑橘刚展开的嫩叶反面上,当蚜虫口针刺入植物组织时,回路接通,回路经AD转换器放大转换为数字信号保存于电脑上,再用Probe 3.4软件转化成波形图谱输出在电脑屏幕上保存、判读并转换为可供统计的数字文件[15,16]。所有试验在白天(25±1)℃,法拉第笼里连续记录6 h,1头成蚜和1株柑橘苗只用于1次试验记录,取有效重复数20次进行统计分析。
1.2.2 刺探电位波形分析 EPG的各种波谱代表剌吸式昆虫口针在植物组织内不同的刺探和取食行为,具有不同的生物学:np波为非刺探波形[17,18]。路径波(包括A波、B波、C波)表示蚜虫口针位于表皮与微管束之间,反映的是胞外电势水平。pd波分3个亚波段,即pd-Ⅰ,pd-Ⅱ,pd-Ⅲ,反映了口针刺破细胞膜时所测的膜内外电位差[15]。E1表示蚜虫口针到达筛管后分泌水溶性唾液的相关波形,E2持续时间在8种波形中最多,是蚜虫在韧皮部筛管被动取食的特征波形[19,20]。G波是蚜虫在木质部主动吸食的波形,与维持蚜虫体内水分平衡相关。F波是口针刺探机械阻碍波[18,21]。Stylet+软件(荷兰瓦赫宁根大学,瓦赫宁根,荷兰)记录并输出每一个波形从开始到结束的时间,并用此软件进行数据转化。
1.2.3 数据分析 试验数据采用SPSS 20.0软件进行统计分析。分析前先对检测数据方差齐性和正态性,符合正态分布的数据采用独立样本t检验(P<0.05),不符合正态分布的数据进lg10(n+1)转换或反正弦转换。转换后的数据仍不符合正态分布的采用Mann-Whitney U-test分析。
2 结果与分析
2.1 褐色橘蚜、棉蚜刺探取食全过程的EPG波形
本实验研究结果表明,褐色橘蚜无翅型、有翅型在感染CTV植株上记录到7种波形(np,A,B,C,pd,E,F)(图1),棉蚜无翅型、有翅型分别在感染CTV植株6 h刺探取食中均记录到8种波形(np,A,B,C,pd,E,F,G)(图2),以上波形与蚜虫的典型波形相似。
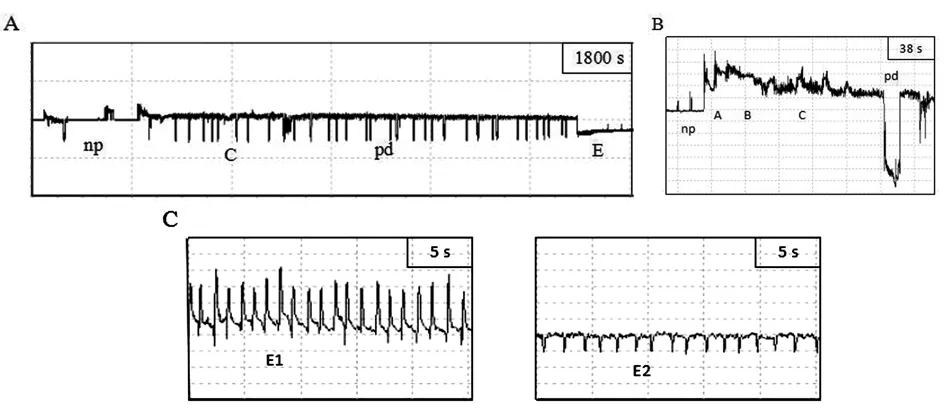
图1 褐色橘蚜有翅型在CTV植株上的刺探取食的波形
注:A:刺探取食过程的整体波形图;B:刺探过程中的路径波;C:韧皮部取食波(E1波、E2波)。
图2 棉蚜有翅型在CTV植株上的刺探取食的波形
注:A:刺探取食过程的整体波形图;B:刺探过程中的路径波;C:机械阻碍波(F波);D:韧皮部取食波(E1波、E2波);E:木质部取食波(G波)。
2.2 褐色橘蚜、棉蚜无翅型和有翅型分别在感病植株上的刺探行为比较
褐色橘蚜无翅型在感病植株上取食的非韧皮部阶段和韧皮部阶段,统计的18个参数中,均与有翅型无差异显著(表1)。与有翅型相比,棉蚜无翅型取食的非韧皮部阶段,统计的12个参数中有2个具有显著性差异(P<0.05),有3个具有极显著性差异(P<0.01);韧皮部阶段统计的6个参数均无显著性差异。
在非韧皮部阶段,棉蚜无翅型在感病植株上的np波的总持续时间、C波的总持续时间、pd波的次数均显著小于有翅型。棉蚜无翅型在感病植株上的C波的次数、刺探的次数均极显著小于有翅型。其他非韧皮部参数均差异不显著(P>0.05)。
2.3 不同橘蚜在感病植株上的刺探行为比较
与棉蚜无翅型相比,褐色橘蚜无翅型在感病植株上取食的非韧皮部阶段,统计的12个参数中有1个具有显著性差异(P<0.05),韧皮部阶段,统计的6个参数中,有1个具有极显著性差异(P<0.01),其余统计参数均无差异显著。其中非韧皮部阶段的pd波的次数和韧皮部阶段的E1波的平均持续时间显著小于棉蚜无翅型(P<0.05)(表1)。
与棉蚜有翅型相比,褐色橘蚜有翅型取食的非韧皮部阶段,统计的12个参数中有5个具有显著性差异(P<0.05),有1个具有极显著性差异(P<0.01);韧皮部阶段统计的6个参数中有1个具有显著性差异(P<0.05),有2个具有极显著性差异(P<0.01)。
在非韧皮部阶段,褐色橘蚜有翅型在感病植株上的刺探次数、C波的次数、pd波的次数、pd波的总持续时间均极显著小于棉蚜有翅型。褐色橘蚜有翅型在感病植株上的第一次刺探至第一次到达韧皮部的时间显著小于棉蚜有翅型。其他非韧皮部参数均差异不显著。韧皮部参数包括与唾液分泌相关的E1波和吸食植物汁液相关E2波参数。褐色橘蚜有翅型在感病植株上的E1波次数、E1波的总持续时间与棉蚜有翅型相比无差异,但E1波的平均持续时间差异显著,且褐色橘蚜有翅型的(1.38±0.40) min大于棉蚜有翅型的(0.63±0.13) min。褐色橘蚜有翅型在感病植株上的E2波的总持续时间和E2波的平均持续时间均极显著大于棉蚜有翅型。
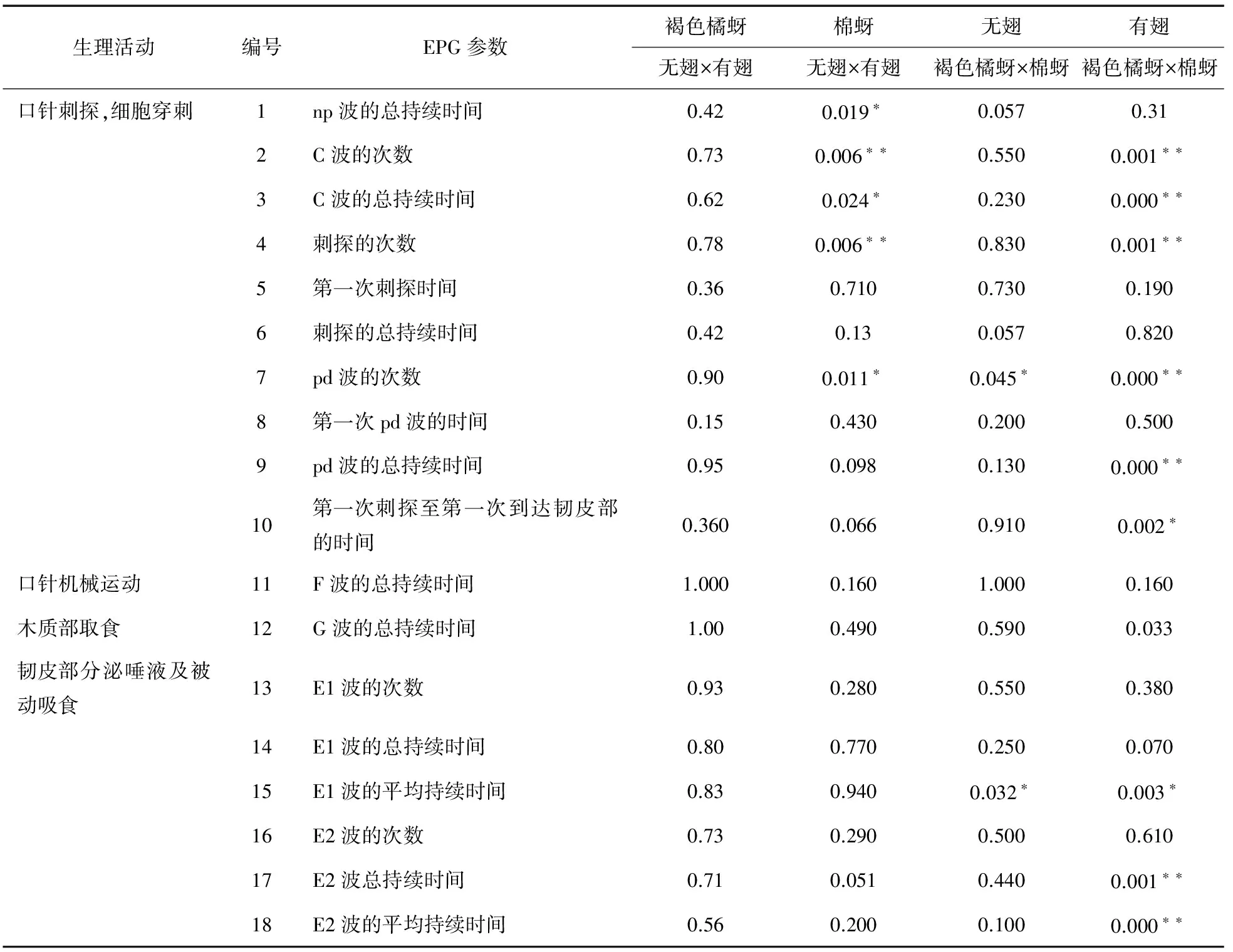
表1 褐色橘蚜、棉蚜无翅型和有翅型在感病植株上的取食行为参数比较
注:表中数据为平均数±标准误。同行数据后“*”和“**”分别表示检验后在P<0.05和P<0.01水平差异显著,下同。
2.4 褐色橘蚜和棉蚜无翅型、有翅型分别在健康和感病植株上的刺探行为比较
在褐色橘蚜无翅型取食的非韧皮部阶段,统计的11个参数中有3个具有显著性差异(P<0.05),有4个具有极显著性差异(P<0.01);韧皮部阶段统计的5个参数中有1个具有极显著性差异(P<0.01)。褐色橘蚜有翅型取食的非韧皮部阶段和韧皮部阶段,统计的参数中均没有显著性差异(P>0.05)。棉蚜无翅型、有翅型在感病植株的非韧皮部阶段和韧皮部阶段的取食行为参数,即统计的16个参数(表2)中,均与健康植株无差异显著。
在非韧皮部阶段,褐色橘蚜无翅型在感病植株上的刺探次数、pd波的次数均显著高于健康植株,但总刺探时间显著短于健康植株。褐色橘蚜无翅型在感病植株上的np波总持续时间、C波的总持续时间、非韧皮部阶段总持续时间极显著高于在健康植株。其他非韧皮部参数,包括从EPG试验开始到第一次刺探的时间、第一次刺探持续时间、pd的平均持续时间以及第一次E波之后刺探的次数均差异不显著。韧皮部参数包括与唾液分泌相关的E1波和吸食植物汁液相关E2波参数。褐色橘蚜无翅型在健康植株上的E1波次数、E1波的总持续时间和E2波次数与在感病植株上相比无差异,但E2波的总持续时间差异极显著,且健康植株上的(295.06±9.99) min大于感病植株上231.30±21.78 min。褐色橘蚜无翅型和有翅型成蚜在健康和感病植株上大于10 minE2波百分比均较高(表2)。
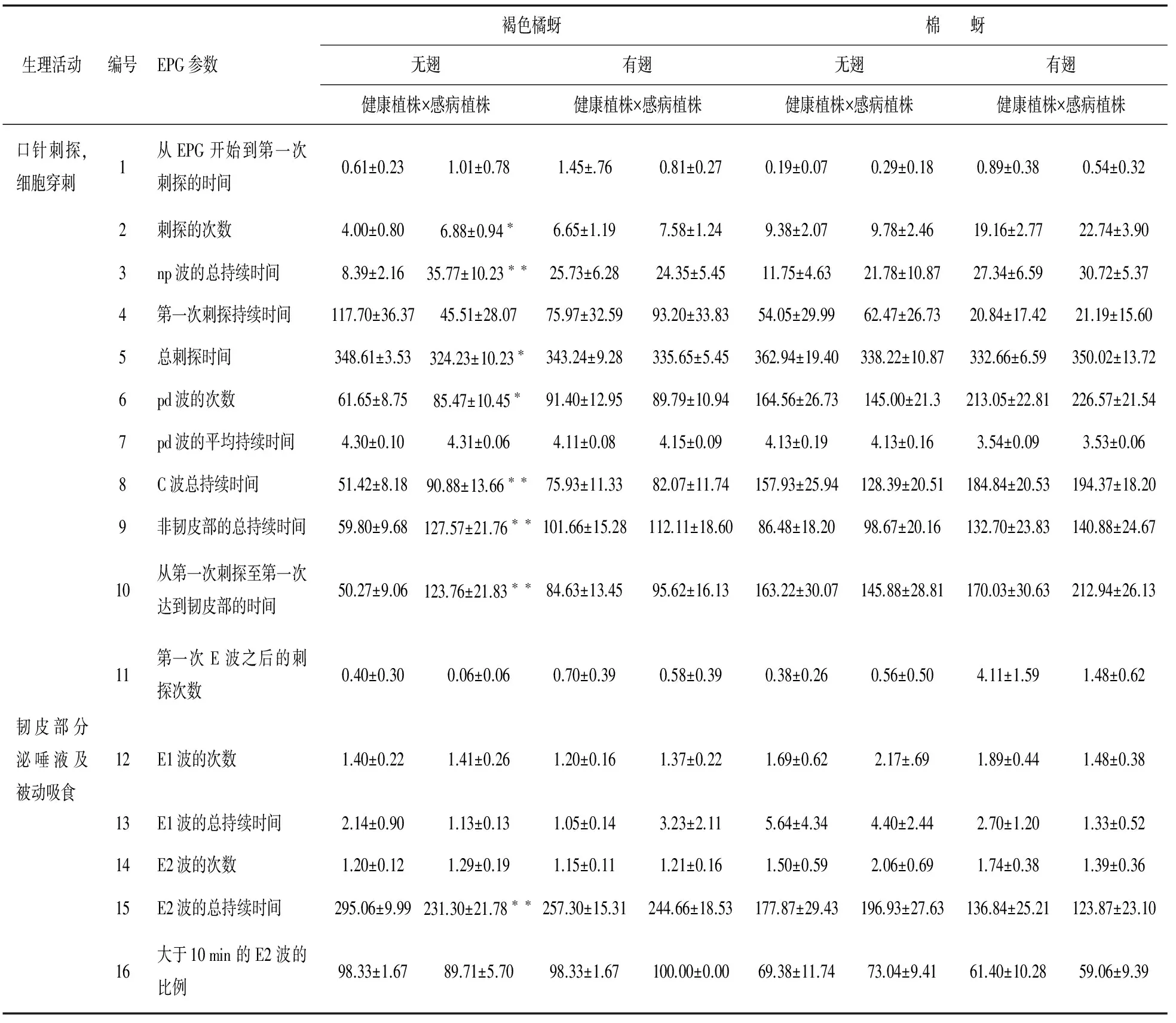
表2 褐色橘蚜和棉蚜无翅型、有翅型分别在健康柑橘植株与感染CTV柑橘植株上的EPG参数比较
3 结论与讨论
3.1 褐色橘蚜、棉蚜无翅型和有翅型在柑橘衰退病植株上的取食行为
在感病植株上,褐色橘蚜无翅型和有翅型没有取食行为差异;棉蚜无翅型较有翅型具有取食行为优势。非韧皮部刺探阶段是蚜虫口针与植物表面接触后刺入表皮通过叶肉细胞间隙的过程[30],蚜虫口针往往需要经过多次刺探才能在到达韧皮部并找到合适的吸食位点[31,32]。若刺探次数越多,刺探时间越长,说明蚜虫越难寻找到合适的吸食位点,取食效率就越低。韧皮部是蚜虫获取生长发育所需能量的唯一途径。韧皮部取食参数中,韧皮部取食时间是最能直接反映蚜虫寄主适应性的参数。 本研究结果显示,棉蚜有翅型在感病植株上需要更多C波的次数、刺探的次数、pd波的次数,需要更长np波的总持续时间、路径波C波的总持续时间,这表明棉蚜有翅型比无翅型取食效率低,这与棉蚜无翅型、有翅型在健康植株上取食行为相似(另文发表)。褐色橘蚜在无翅型、有翅型在感病植株上的取食行为无差异,但是在健康植株上取食行为显示仅np波的总持续时间存在差异。表明,在CTV侵染及健康的柑橘上,褐色橘蚜无翅型、有翅型取食几乎无显著差异,棉蚜无翅型均较有翅型具有取食行为优势,柑橘感病状态对无翅型和有翅型的影响是相似的。刘佰明(2012)报道了无毒的B型和Q型烟粉虱在TYLCV侵染及健康的番茄上的取食行为相似,且Q行型均较B型具有行为优势[33]。
在感病植株上,褐色橘蚜无翅型较棉蚜无翅型具有取食行为优势,棉蚜无翅型在非韧皮阶段需要更多的pd波的次数,在韧皮部被动取食之前平均需要更长时间唾液分泌,且E2波的总吃持续时间差异不显著,这表明棉蚜无翅型比褐色橘蚜无翅型取食效率低;褐色橘蚜有翅型较棉蚜有翅型具有取食行为优势,棉蚜有翅型比褐色橘蚜有翅型在非韧皮阶段具有更多的C波的次数、刺探次数、pd波的次数,并且棉蚜有翅型第一次刺探后较褐色橘蚜有翅型需要更长的时间到达韧皮部,这表明棉蚜有翅型比褐色橘蚜有翅型取食效率低;棉蚜有翅型在韧皮部阶段刺吸的总持续时间极显著小于褐色橘蚜有翅型,说明褐色橘蚜有翅型在感病植株上较棉蚜有翅型适宜性更强。以上结果与褐色橘蚜和棉蚜无翅型、有翅型在健康植株上的的结果相似(另文发表),同时说明褐色橘蚜比棉蚜取食能力强,这也与褐色橘蚜比棉蚜传毒能力强相对应[34]。3.2 柑橘衰退病对褐色橘蚜、棉蚜取食行为的影响
蚜虫开始取食都要试食以确定其寄主是否适合,因此口针刺探是寄主选择的必需步骤[22]。EPG技术可以通过比较昆虫在不同抗性品种上的刺探和取食行为,从而确定抗性因子存在的部位[23],若短暂口针刺探次数、路径刺探时间以及非取食阶段时间较长,说明抗性因子存在于植物表面或表皮层和叶肉层;若唾液分泌时间较长,韧皮部取食时间比较短,抗性因子存在于韧皮部[11]。鲁卓越等[24]研究结果显示,褐色橘蚜在黄皮上需更多时间寻找合适的取食位点,其表面可能存在影响取食的物理因子;蚜虫在黄皮上的第1次持续刺吸时间显著小于其余寄主,第1次到达韧皮部前的刺探次数、短暂刺吸次数(<3 min)、pd波次数均显著大于其余寄主,表明黄皮的表皮-叶肉组织中存在某些抗性因子阻碍或干扰其持续取食。本研究结果表明,褐色橘蚜在感病植株上需要较长的时间进行刺探,刺探次数和pd波次数显著多于健康植株,非韧皮部阶段的总持续时间极显著大于健康植株,推测健康柑橘感染柑橘衰退病8个月后柑橘表面物理因子和叶肉化学因子变化,阻碍褐色橘蚜无翅型成蚜刺吸韧皮部;褐色橘蚜无翅型在感病植株上E2波的取食时间极显著短于健康植株,进一步说明植物抗虫因子在韧皮部以外[25]。
植物病毒侵染寄主植物一般通过改变寄主的特性(营养组成、挥发性有机物的浓度和组分等)来影响介体的取食取食行为[7,25]。关于介体对感染病毒的植物的偏好存在一定的争议。研究感染TSWV的雄虫在烟草上较健康的雄虫摄取更多的食物,摄食频率也增加了3倍,从而增加接种病毒的概率[26]。Mauck等[27]研究结果表明,黄瓜花叶病毒(Cucumbermosaicvirus,CMV)侵染南瓜后,两种蚜虫不偏好CMV侵染的南瓜。本研究结果表明,褐色橘蚜无翅型接触感染CTV4强毒株系8个月后的柑橘苗需花较长的时间到达韧皮部,而在健康植株上能很快到达韧皮部取食;褐色橘蚜无翅型到达感病植株韧皮部,E2的总持续时间极显著短于健康植株,表明感病植株可降低寄主对褐色橘蚜无翅型的适合度,褐色橘蚜无翅型蚜虫偏好健康植株上取食,与何应琴等[28]对褐色橘蚜在接种CTV11弱毒株系3个月后的取食结果不一致。此外,Cen等[29]研究表明柑橘木虱在不同的发病程度或不同病毒含量上的取食行为不同。因此柑橘叶片在不同感病时期的营养成分和病毒含量是否是导致褐色橘蚜取食行为的不同,还有待进一步研究。在感病植株上,棉蚜无翅型与健康植株无显著差异,或许更长的EPG记录时间能够提供在感病柑橘上有利的证据,需要进一步的研究。
任何不利的环境因素均可引起有翅蚜的产生[35],褐色橘蚜无翅型蚜虫不喜欢取食感病植株,将有利于有翅蚜的形成而离开原寄主。而且与健康植株相比,褐色橘蚜有翅型在感病植株上的取食没有显著差异,从而促进病毒传播。
[1] Barjoseph M. The closteroviruses:a distinct group of elongated plant viruses[J].AdvancesinVirusResearch, 1979, 25(25):93-168.
[2] Koonin E V, Dolja V V. Evolution and taxonomy of positive-strand RNA viruses: implications of comparative analysis of amino acid sequences[J].CriticalReviewsinBiochemistryandMolecularBiology, 1993, 28(5),375.
[3] Fereres A, Moreno A. Behavioural aspects influencing plant virus transmission by Homopteran insects[J].VirusResearch, 2009, 141(2):158-168.
[4] Belliure B, Janssen A, Maris P D,etal.Herbivore arthropods benefit from vectoring plant viruses[J].EcologyLetters, 2005, 8(1):70-79.
[5] Bautista R C, Rfl M, Cho J J,etal. Potential of tomato spotted wilt tospovirus plant hosts in hawaii as virus reservoirs for transmission byFrankliniellaoccidentalis(Thysanoptera: Thripidae)[J].Phytopathology, 1995, 85(9):953-958.
[6] Maris P C, Joosten N N, Goldbach R W,etal. Tomato spotted wilt virus infection improves host suitability for its vectorFrankliniellaoccidentalis[J].Phytopathology, 2004, 94(7):706-716.
[7] Jiménez-Martínez E S, Bosque-Pérez N A, Berger P H,etal.Volatile cues influence the response ofRhopalosiphumpadi(Homoptera: Aphididae) to barley yellow dwarf virus-infected transgenic and untransformed wheat[J].EnvironmentalEntomology, 2004, 33(5):1207-1216.
[8] Fiebig M, Poehling H M,Borgemeister C. Barley yellow dwarf virus, wheat, and sitobion avenae: a case of trilateral interactions[J].EntomologiaExperimentalisEtApplicata, 2004, 110(1):11-21.
[9] Tjallingii W F. Comparison of ac and dc systems for electronic monitoring of stylet penetration activities by Homopterans[J].JournalofEvolutionaryBiology, 2001, 21(04):1655-1663.
[10] 陈建明,俞晓平,程家安.电子取食监测仪在植物抗虫性研究中的应用[J].应用昆虫学报,2005,42(5): 485-490.
[11] 罗 晨,岳 梅,徐洪富,等.EPG技术在昆虫学研究中的应用及进展[J].昆虫学报,2005,48(3):437-443.
[12] 高伟民,刘建阳. 烟蚜与烟草黄瓜花叶病田间消长动态研究[J].福建热作科技,2009,34(2):8-9.
[13] 成巨龙,安德荣,孙 渭.陕西省烟草蚜传病毒的发生发展规律及预测预报模型的初步研究[J].中国烟草学报,1998,(2):43-48.
[14] 李玲娣,周常勇,李中安,等.褐色橘蚜中柑橘衰退病毒实时荧光定量RT-PCR检测方法的建立与应用[J].中国农业科学,2013,46(3):525-533.
[15] Tjalningii W F.Membrance potentials as an indication for plant cell penetration by aphid stylets[J].EntomologiaExperimentalisetApplicata,1985(38):187-193.
[16] 岳 梅,罗 晨,郭晓军,等. B型烟粉虱在甘蓝、西葫芦和辣椒上的刺吸取食行为[J].昆虫学报,2006, 49(4):625-629.
[17] Tjallingii W F. Eletronic recording of penetration behavior by aphids[J].EntomologiaExperimentalisEtApplicata, 1978, 24(3):721-730.
[18] Hu X, Liu X, Zhao H. Development and application of electrical penetration graph (EPG) technique[J].PlantProtection, 2006, 32(3):1-4.
[19] Prado E, Tjallingii W F. Aphid activities during sieve element punctures[J].EntomologiaExperimentalisEtApplicata, 1994, 72(2):157-165.
[20] Spiller N J, Koenders L, Tjallingii W F. Xylem ingestion by aphids -a strategy for maintaining water balance[J].EntomologiaExperimentalisEtApplicata, 1990, 55(2):101-104.
[21] Tjallingii W F. Salivary secretions by aphids interacting with proteins of phloem wound responses[J].JournalofExperimentalBotany, 2006, 57(4):739-745.
[22] Harris K F, Maramorosch K. Aphids as virus vectors[M].AcademicPressNewYork, 1977:1-559.
[23] 雷 宏,徐汝梅. EPG—一种研究植食性刺吸式昆虫刺探行为的有效方法[J].应用昆虫学报,1996,(2):116-120.
[24] 鲁卓越,陈文龙,何应琴,等.褐色橘蚜在五种柑橘上的取食行为差异[J].植物保护学报,2014,41(5):637-638.
[25] 施 艳,王英志,汤清波,等. 昆虫介体行为与植物病毒的传播[J].应用昆虫学报,2013,50(6):1719-1725.
[26] Stafford C A, Ullman D E. Infection with a plant virus modifies vector feeding behavior[J].ProcNatlAcadSciUSA, 2011, 108(23):9350-9355.
[27] Mauck K E, De Moraes C M, Mescher M C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2010, 107(8):3600-3605.
[28] 何应琴,陈文龙,鲁卓越,等. 褐色橘蚜在健康与CTV植株上的EPG比较[J]. 山地农业生物学报,2014,33(2):36-39.
[29] Cen Y J, Yang C L, Holford P,etal. Feeding behaviour of the Asiatic citrus psyllid,Diaphorinacitri, on healthy and huanglongbing -infected citrus[J].EntomologiaExperimentalisEtApplicata, 2012, 143(1):13-22.
[30] Gabrys B, Tjallingii W F, Beek T A V. Analysis of EPG Recorded Probing by Cabbage Aphid on Host Plant Parts with Different Glucosinolate Contents[J].JournalofChemicalEcology, 1997, 23(7):1661-1673.
[31] Lei D H, Tjallingii W F, Lenteren J C V. Effect of tethering during EPG recorded probing by adults of the greenhouse whitefly[J].JournalofAppliedEntomology, 1997, 121(4):211-217.
[32] Lei H, Lenteren J C, Xu R M. Effects of plant tissue factors on the acceptance of four greenhouse vegetable host plants by the greenhouse whitefly: an electrical penetration graph (EPG) study[J].EJE, 2001, 98(1):31-36.
[33] 刘佰明. B型和Q型烟粉虱竞争取代的取食行为学机制[D]. 北京:中国农业科学院,2012.
[34] 周 彦,王雪峰,陈洪明,等.柑橘衰退病毒含量对其蚜传效率的影响[J]. 植物保护学报,2015,42(1):93-98.
[35] 罗礼智,李光博.植物病毒病对蚜虫种群增长与翅型分化的影响[J]. 植物保护,1991,17(5):34-36.
Effects of Citrus Tristeza Virus Strains on Feeding Behavior of Two Species of Citrus Aphids with Different Wing Type
ZHAORu-na1,HEYing-qin1,LUZhuo-yue1,CHENWen-long1*,YANGYu-yin1,WANGXue-feng2,ZHOUChang-yong2
(1.GuizhouKeyLaboratoryofPlantDieseaseandPestManagementforMountainAgriculture/InstituteofEntomology,ScintificObservingandExperimentalStationofCropPestinGuiyang,MinistryofAgriculturePRChina,GuizhouUniversity,Guiyang,Guizhou550025,China; 2.CitrusResearchInstitute,ChineseAcademyofAgricultruralSciences,Beibei,Chongqing400712,China)
To evaluate the effects of Citrus tristeza virus-infected citrus plants on the feeding behavior of different wing dimorphisms, feeding behaviors between alate and apterous ofToxopteracitricida(Kirkaldy)andAphisgossypiiGlover on the CTV-infected plants were explored using the electrical penetration graph (EPG) techniques and comparing to healthy citrus plants. The results showed that apterous ofT.citricidaon CTV-infected citrus plants had no significant difference comparing to alate one’s. Some EPG parameters ( total time of np and C、number of potential drops) of apterous ofA.gossypiiGlover on the CTV-infected was significantly smaller than those of alate ofA.gossypiiGlover, while number of probes and C on apterous ofA.gossypiiGlover were extremely significantly smaller than those of alate ofA.gossypiiGlover. The number of pd and duration time of E1 of apterous ofT.citricida(Kirkaldy)on the CTV-infected were significantly smaller than those of apterous ofA.gossypiiGlover. However, EPG parameters of alate ofT.citricida(Kirkaldy) on number and total duration time of C, number of probes, number and total duration time of pd were significantly smaller than those of alate ofA.gossypiiGlover, but bigger on total duration time and average duration time of E2. For apterous ofT.citricida(Kirkaldy)on the CTV-infected plants, number of probes and number of pd were significantly longer than healthy citrus plants, but total duration of probing was opposite. The duration time of np and C waveforms, total duration of no phloematic phase of apterousT.citricidaon CTV-infected citrus plants were longer than healthy citrus plants; the duration time of E2 and percent of E2 were shorter than healthy citrus plants. These results implied that some similarity of EPG parameters of alate and apterous ofT.citricida(Kirkaldy)and Aphis gossypii Glover feeding were observed on the infected plants. The effect of CTV-infected citrus plants on the feeding behavior of apterousT.citricidawas obvious, and apterousT.citricidaprefered healthy citrus plants.
electrical penetration graph;citrustristezavirus; alate aphid; apterous aphid; virus transmission
2017-03-08;
2017-06-01
国家公益性行业(农业)科研专项(201203076)。
S443.3
A
1008-0457(2017)04-0013-08 国际
10.15958/j.cnki.sdnyswxb.2017.04.003
*通讯作者:陈文龙(1965-),男,博士,教授,主要研究方向:生物防治与害虫综合治理;E-mail:cwl001@163.com。
猜你喜欢
植物保护(2023年1期)2023-02-03 10:22:08
农药学学报(2022年6期)2022-12-27 12:00:28
浙江林业科技(2021年3期)2021-07-13 09:03:56
新疆农业科学(2020年11期)2020-10-27 09:05:32
长江大学学报(自科版)(2018年14期)2018-07-17 08:37:02
现代农业(2016年4期)2016-02-28 18:42:12
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
热带作物学报(2014年3期)2014-04-29 18:17:49
中国果业信息(2014年11期)2014-01-23 13:43:33
