
野生羊茅与引进羊茅‘猎狗5号’竞争特性的研究
2017-07-30 11:29刘佳星呼丽瑶
草地学报 2017年6期
刘佳星,王 雷,呼丽瑶
(内蒙古农业大学林学院, 内蒙古 呼和浩特 010019)
在我国,针对草坪业发展,草坪草各种条件的生理生化反应、抗逆性及转基因研究的成果较多[1-8];对本土草坪草的开发及推广也有部分研究,但后续效果并不明显[9,10]。内蒙古地区本土草坪草即大青山草地早熟禾(PoapratensisL.‘ Daqingshan’)由内蒙古农牧科学院通过采集野生草坪草经多年培育而成[11]。赵文丽等对内蒙古地区本土草坪草--野生羊茅(FestucaovinaL.)的一些特性及应用进行了研究[12]。
目前,呼和浩特市及周边地区所使用的草坪绝大部分是引进的草地早熟禾(PoapratensisL.)(包括‘午夜2号’、‘新哥来德’、‘纳苏’、‘肯塔基’等)数个品种及少量高羊茅(FestucaarundinacesSchreb.)(包括‘猎狗5号’、‘美洲虎4G’、‘贝克’等几个品种)。引进的冷季型草坪草自播性很强,繁殖力旺盛,对周边生态系统具有很强的侵入性。由于引进草坪草品种的大面积栽植,导致呼市城市绿地系统草坪层结构单一,进而导致城市绿地系统功能不稳定,一旦发生引进草坪草的易感病、虫害,就会导致同期草坪草的大面积退化,城市绿地功能衰退。
因此,对呼市地区本土草坪草与引进草坪草的竞争特性进行研究从而降低本地区外来物种入侵率已经迫在眉睫。在环境适宜性假说(the favourable environment hypothesis)及空间异质化假说(the spatial heterogeneity hypothesis)条件下,拟研究本土草坪草与引进草坪草品种的各种竞争组合模式生物多样性的动态变化,分析各种条件对本土草坪草与引进草坪草竞争能力的影响。探究本土草坪草与引进草坪草品种和谐共存的发展之路,维护绿地系统功能的健全与稳定具有重要意义。
1 试验材料与方法
1.1 供试草坪草品种及试验前期处理
供试材料为内蒙古地区本土冷季型草坪草野生羊茅(FestucaovinaL.)与引进的冷季型草坪草的代表性品种高羊茅(FestucaarundinacesSchreb.)中的‘猎狗5号’(‘Houndog Ⅴ’),见表1。

表1 供试材料一览表Table 1 Information sheet of testing materials
前期,每种草坪草品种都在相同的基质条件下单独培育、挂标签标记(如图1),后期根据实验要求,通过移栽形成实验所需竞争组合模式。

图1 草坪草前期室外培养Fig.1 Outdoor cultivation of turfgrass in the early stage
将培养盆置于内蒙古农业大学东区校园内进行室外培养,发芽后进行常规水肥管理。待草坪草进行过3次修剪后,进行后续实验处理。
1.2 试验设计与管理
1.2.1栽培基质 本研究中各种条件下的栽培基质统一配制,考虑草坪移栽、管理的便捷化,同时考虑草坪栽植基质的匀质化,本研究参照USGA运动场草坪建植基质进行配制(配制为体积比):70%水洗沙(16—64目)+20%有机肥+10%(草炭+氮磷钾全效复合肥+少量微肥)混合物构成。
1.2.2草坪草竞争组合模式 本试验中将野生羊茅与猎狗5号进行混栽以形成竞争组合模式,试验中均需保证2种草坪草植株数量比例始终为1:1。本试验中,草坪草栽植密度1株·cm-2(应用空间异质化假说进行样品处理时,不同栽植密度条件下实验除外)。为了精确而有效控制竞争组合模式下的草坪栽植密度,本实验光照培养箱中使用的培养皿为穴盘(图2)。

图2 各种规格穴盘Fig.2 The specification of plug trays
本实验中1株·cm-2的栽植密度下使用规格为5*10孔的穴盘(每穴上口径50 mm,下口径22 mm,穴深45 mm),2株·cm-2时使用规格为12*6孔(每穴上口径40 mm,下口径20 mm,穴深45 mm)的穴盘,4株·cm-2时使用规格为12*24孔的穴盘(每穴上口径20 mm,下口径10 mm,穴深45 mm)。草坪草进行移栽后的情况见图3。

图3 草坪草移栽培养Fig.3 Transplanting culture of turfgrass
1.2.3两种假说下的研究方法设计
1.2.3.1 环境适宜性假说下运用的研究方法
本研究实验以光照培养箱及小型气候室为设备基础,进行条件精确控制的可控实验,本组实验中将光照培养箱的各项条件均设定为冷季型草坪草适宜生长的环境。
将光照培养箱及小型气候室温度设定为恒温20℃;每天光照时间为8 h,光照强度12 000 LUX;由于是沙基质草坪,故平均2 d喷水一次,通过湿度感应器,实验样品组合基质湿度控制在45%~55%之间,记录湿度值;平均5 d修剪一次,留茬高度6 cm。
将上文1.2.2中的野生羊茅与猎狗5号形成的竞争组合模式(组合设3个重复)置于条件适宜的光照培养箱中,待培育1星期后:(1)每隔3 d统计组合中每个品种的株数并计算辛普森指数;(2)同时,每隔5 d对组合中每个品种分茎叶(地上部分)、根系(地下部分)分别取样,3次重复,进行生理因子测定分析(主要包括可溶性蛋白、可溶性糖、脯氨酸等草坪营养物质);(3)综合分析本土草坪草与引进草坪草竞争能力以及竞争模式下的生物多样性,共取5次样。
1.2.3.2 空间异质化假说下运用的研究方法
本系列实验同样以光照培养箱为设备基础,进行条件精确控制的可控实验。运用扰动原理,使实验条件分别处于不同的非理想的扰动水平上。
不同修剪频率下:将光照培养箱温度设定为恒温20℃;每天光照时间为8 h,光照强度12 000 LUX;实验样品组合基质湿度控制在45%~55%之间,记录湿度值;修剪留茬高度6 cm。修剪频率设定1次/5 d、1次/10 d和1次/15 d这3个处理水平。
将上文1.2.2中的野生羊茅+‘猎狗5号’的竞争组合模式(组合设3个重复)分别按照不同修剪频率组中的不同处理条件,将竞争模式组合置于光照培养箱中培养,待培育1星期后:(1)每隔5 d统计组合中每个品种的株数并计算辛普森指数;(2)同时,每隔5 d对组合中每个品种分茎叶(地上部分)、根系(地下部分)分别取样,5次重复,进行生理因子测定分析(主要包括可溶性蛋白、可溶性糖、脯氨酸等草坪营养物质);(3)在综合分析空间异质化条件下,本土草坪草与引进草坪草竞争能力以及竞争模式下的生物多样性,每个处理下至少取5次样结束。
1.3 实验方法
1.3.1记录、统计分析 辛普森指数测定采用公式
公式中,S为物种数目;种i的个体数占群落中总个体数的比例(Pi)进行计算。由辛普森指数计算公式可知,辛普森指数既可以反映物种丰富度,也可以间接反映群落的稳定性。
测定各处理条件下各组合中的组分株数。
1.3.2植物样品测定
可溶性糖含量的测定:采用蒽酮比色法;
可溶性蛋白含量的测定:考马斯亮蓝G-250染色法;
脯氨酸含量的测定:酸性茚三酮法。
1.4 数据分析
所得数据采用Excel 2010进行整理和简单分析处理后,利用SPSS 17.0统计软件对数据进行单因素方差分析,并釆用折线图和柱状图的方法表示各胁迫处理的生理生化指标变化。
2 结果与分析
2.1 环境适应性假说下的结果与分析
2.1.1辛普森指数的计算结果与分析 对移栽至穴盘的草坪草竞争组合进行环境适应性假说下的条件处理后,每隔5 d统计各品种的株数,统计5次结束。3次重复下2种草坪草5次统计的平均株数、组合的辛普森指数均值及每种草坪草株数占组合总株数的比率(Pi)的均值见表2。
从表2中可以看出,在环境适应性假说下,野生羊茅与猎狗5号移栽组合的辛普森指数为0.500。其中,野生羊茅株数占组合总株数的比率为49%,猎狗5号为51%,由此可见,在环境适应性假说下,2种草坪草占组合总株数的比率相差不大。

表2 环境适应性假说下草坪草统计的平均株数、森普森指数及Pi值Table 2 The average number of turfgrass, Simpson Index and Pi values under the environmental adaptability hypothesis
2.1.2可溶性糖的测定结果与分析
在环境适应性假说处理下,对野生羊茅和猎狗5号各部位进行可溶性糖含量的测定,5次测定下2种草坪草各部位可溶性糖平均含量的均值分别为:野生羊茅地上部分为24.590 mg·g-1,野生羊茅地下部分为10.496 mg·g-1,猎狗5号地上部分为26.278 mg·g-1,猎狗5号地下部分为13.209 mg·g-1。5次测定下的可溶性糖平均含量的变化趋势见图4。由图4可以看出,2种草坪草茎叶部分的可溶性糖平均含量均比根部的含量高,‘猎狗5号’的地上和地下部分的可溶性糖平均含量均比野生羊茅对应部分的可溶性糖平均含量高。且5次测定下各部位的可溶性糖平均含量基本呈缓慢增加趋势。这可能是由于草坪草移栽后,生长逐渐趋于稳定,叶片和根系的生长呈上升趋势,而且叶片的光合作用也在逐渐加强,因此所测得的草坪草各部位的可溶性糖平均含量缓慢增加。

图4 环境适应性假说下野生羊茅和猎狗5号各部位可溶性糖平均含量的变化Fig.4 Changes of soluble sugar contents in different parts of Wild Festuca and ‘Houndog V’ under the environmental adaptability hypothesis
2.1.3可溶性蛋白的测定结果与分析
在环境适应性假说处理下,对野生羊茅和‘猎狗5号’各部位进行可溶性蛋白平均含量的测定,5次测定下2种草坪草各部位可溶性蛋白平均含量的均值分别为:野生羊茅地上部分为3.884 ug·g-1,野生羊茅地下部分为2.819 ug·g-1;‘猎狗5号’地上部分为3.930 ug·g-1,‘猎狗5号’地下部分为2.823 ug·g-1。5次测定下的可溶性蛋白平均含量的变化趋势见图5。由图5可以看出,2种草坪草茎叶部分的可溶性蛋白平均含量均比根部的含量高,猎狗5号的地上和地下部分的可溶性蛋白平均含量微高于野生羊茅对应部分的可溶性蛋白平均含量,差距不显著。且5次测定下各部位的可溶性蛋白平均含量基本呈缓慢增加的趋势。所测得的草坪草各部位的可溶性蛋白平均含量缓慢增加,可能是由于草坪草经过移栽,生长逐渐趋于稳定后,叶片和根系的生长呈上升趋势,从而产生的可溶性蛋白平均含量也逐渐增多。

图5 环境适应性假说下野生羊茅和‘猎狗5号’各部位可溶性蛋白平均含量的变化Fig.5 Changes of soluble protein contents in different parts of Wild Festuca and ‘Houndog V’ under the environmental adaptability hypothesis
2.1.4脯氨酸的测定结果与分析
在环境适应性假说处理下,对野生羊茅和猎狗5号各部位进行脯氨酸平均含量的测定,5次测定下2种草坪草各部位脯氨酸平均含量的均值分别为:野生羊茅地上部分为98.935 ug·g-1,野生羊茅地下部分为41.761 ug·g-1,‘猎狗5号’地上部分为89.748 ug·g-1,‘猎狗5号’地下部分为45.039 ug·g-1。5次测定下的脯氨酸平均含量的变化趋势见图6。由图6可以看出,2种草坪草茎叶部分的脯氨酸平均含量均比根部的含量高,野生羊茅地上部分的脯氨酸平均含量高于‘猎狗5号’地上部分,而野生羊茅地下部分的脯氨酸平均含量却低于‘猎狗5号’地下部分的脯氨酸平均含量。且5次测定下各部位的脯氨酸平均含量同可溶性蛋白和可溶性糖平均含量的变化趋势相同,基本呈缓慢增加的趋势。

图6 环境适应性假说下野生羊茅和‘猎狗5号’各部位脯氨酸平均含量的变化Fig.6 Changes of proline contents in different parts of Wild Festuca and ‘Houndog V’ under the environmental adaptability hypothesis
全先庆等认为植物体内脯氨酸的积累可能是植物用以贮藏能量的一种手段,因为脯氨酸的降解可以为植物体进行呼吸作用和光合作用提供能量[13]。因此,所测得的草坪草各部位的脯氨酸平均含量缓慢增加可能是由于草坪草经移栽生长逐渐趋于稳定后,叶片和根系的生长呈上升趋势,从而产生较多的脯氨酸为植物体提供能量。
2.2 空间异质化假说下不同修剪频率的结果与分析
2.2.1辛普森指数的计算结果与分析 对移栽至穴盘的草坪草竞争组合进行不同修剪频率的处理后,每隔5 d统计各品种的株数,统计5次结束。3次重复下2种草坪草5次统计的平均株数、组合的辛普森指数均值及每种草坪草株数占组合总株数的比率(Pi)的均值见表3。
从表3中可以看出,当修剪频率降低至1次/10 d时,各指数没有明显变化。而在1次/15 d时移栽组合的辛普森指数有所下降,说明此时组合的物种多样性和群落稳定性在逐渐下降。同时,野生羊茅的个体株数占组合总株数的比率有所下降,而‘猎狗5号’的个体株数占组合总株数的比率有所上升。
此结果反映出当修剪频率降低时,组合的野生羊茅的竞争力弱于‘猎狗5号’,其正常生长受到了抑制,因为野生羊茅是野生种,耐修剪性低于改良后的‘猎狗5号’。
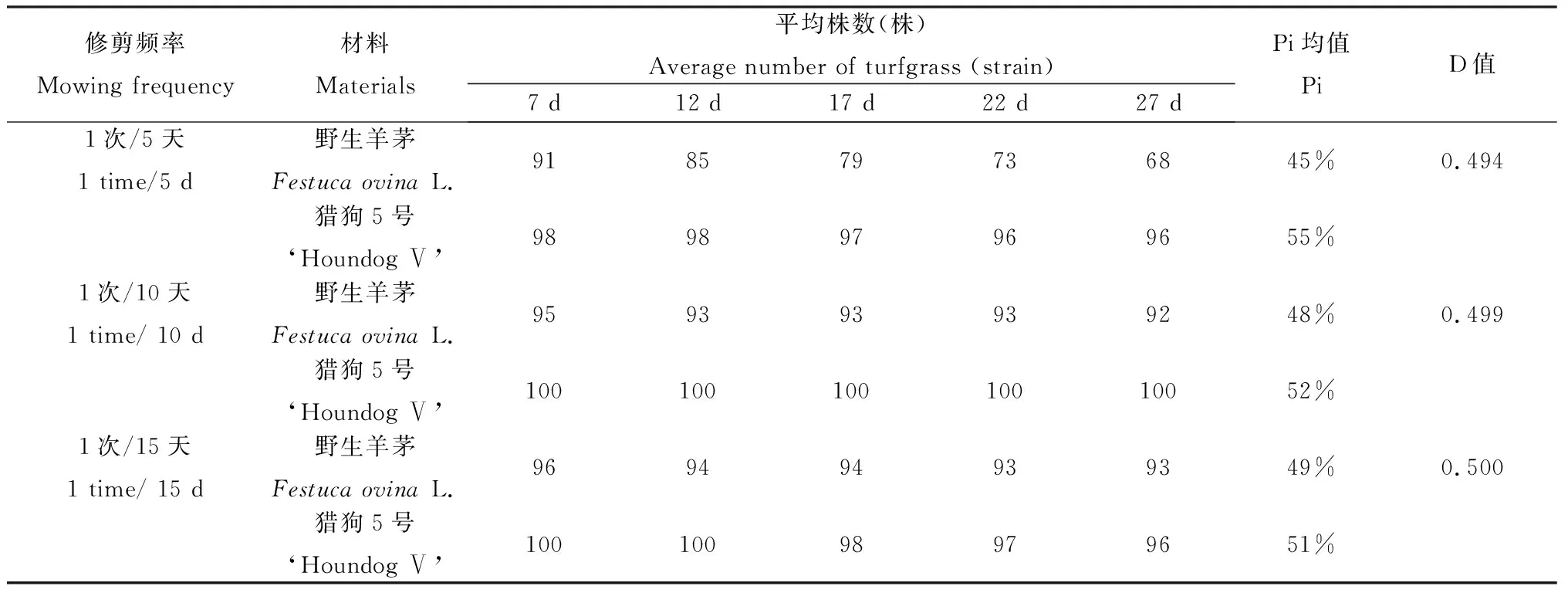
表3 不同修剪频率下草坪草统计的平均株数、森普森指数及Pi值Table 3 The average number of turf grass, Simpson's Index and Pi values under different trim frequencies
2.2.2可溶性糖的测定结果与分析
注:不同的小写字母表示2种草坪草各部位在不同修剪频率下有显著差异(P<0.05)
Note: Different lowercase letters indicate significant difference at the 0.05 level .The same as below
在不同修剪频率处理下,野生羊茅和猎狗5号各部位每隔5 d可溶性糖平均含量的测定结果见表4。由表4计算可得,修剪频率控制到1次/10 d时,5次测定的野生羊茅地上部分可溶性糖平均含量的均值为21.805 mg·g-1,与1次/5 d的适宜修剪频率下相比降低了11.34%;野生羊茅地下部分为9.551 mg·g-1,与1次/5 d时相比降低了9%;‘猎狗5号’地上部分为22.758 mg·g-1,与1次/5时相比降低了13.40%;‘猎狗5号’地下部分为11.870 mg·g-1,与1次/5 d时相比降低了10.14%。1次/15 d时,5次测定的野生羊茅地上部分可溶性糖平均含量的均值为20.799 mg·g-1,与1次/5 d时相比降低了15.42%;野生羊茅地下部分为9.011 mg·g-1,与1次/5 d时相比降低了14.15%;‘猎狗5号’地上部分为22.281 mg·g-1,与1次/5 d时相比降低了15.21%;‘猎狗5号’地下部分为11.432 mg·g-1,与1次/5 d时相比降低了13.45%。
不同修剪频率下2种草坪草各部分可溶性糖平均含量的变化趋势见图7。由表4和图7可以看出,2种草坪草地上部分的可溶性糖平均含量均比地下部分的可溶性糖平均含量高。随着修剪频率的降低,2种草坪草各部分的可溶性糖平均含量均逐渐减少,且降低幅度逐渐减缓。且2种草坪草地上部分的降低幅度均大于地下部分。其中,1次/10 d时野生羊茅地上部分和地下部分的降低幅度均略低于‘猎狗5号’的相应部分;而1次/15 d时,野生羊茅地上部分和地下部分的降低幅度均高于‘猎狗5号’的相应部分。
由于修剪可以更好的促进草坪草的生长与分蘖。因此随着修剪频率的降低,草坪草的生长速度逐渐减缓,植物体内产生的可溶性糖含量就会逐渐减少。而随着修剪频率的逐渐降低,野生羊茅地上部分和地下部分的降低幅度由差异性较小到逐渐高于‘猎狗5号’的相应部位,可能是由于修剪频率对野生羊茅的影响强于‘猎狗5号’。

图7 不同修剪频率下野生羊茅和猎狗5号各部位可溶性糖平均含量的变化Fig.7 Changes of soluble sugar contents in different parts of Wild Festuca and ‘Houndog V’ under different trim frequences
2.2.3可溶性蛋白的测定结果与分析
在不同修剪频率处理下,野生羊茅和猎狗5号各部位可溶性蛋白平均含量的测定结果见表5。由表5计算可得,修剪频率控制到1次/10 d时,5次测定的野生羊茅地上部分可溶性蛋白平均含量的均3.381 ug·g-1,与1次/5 d时相比降低了12.95%;野生羊茅地下部分为2.577 ug·g-1,与1次/5 d时相比降低了8.51%;‘猎狗5号’地上部分为3.523 ug·g-1,与1次/5 d时相比降低了10.36%;‘猎狗5号’地下部分为2.619 ug·g-1,与1次/5天时相比降低了7.23%。1次/15 d下,5次测定的野生羊茅地上部分可溶性蛋白平均含量的均值为3.213 ug·g-1,与1次/5 d时相比降低了17.28%;野生羊茅地下部分为2.467 ug·g-1,与1次/5 d时相比降低了12.49%;‘猎狗5号’地上部分为3.322 ug·g-1,与1次/5 d时相比降低了15.47%;‘猎狗5号’地下部分为2.545 ug·g-1,与1次/5 d时相比降低了9.85%。
不同修剪频率下2种草坪草各部分可溶性蛋白平均含量的变化趋势见图8。由表5和图8可以看出,2种草坪草地上部分的可溶性蛋白平均含量均比地下部分的可溶性蛋白平均含量高。随着修剪频率的降低,2种草坪草各部分的可溶性蛋白平均含量均逐渐减少,且降低幅度逐渐减缓。且2种草坪草地上部分的增长幅度均大于地下部分。其中,在不同修剪频率下野生羊茅地上和地下部分的降低幅度明显高于‘猎狗5号’的相应部分。
由于修剪可以更好的促进草坪草的生长与分蘖。因此随着修剪频率的降低,草坪草的生长速度逐渐减缓,植物体内产生的可溶性蛋白含量就会逐渐减少。而在不同修剪频率下,野生羊茅地上部分和地下部分的降低幅度均明显高于‘猎狗5号’的相应部位,可能是由于修剪频率对野生羊茅的影响强于‘猎狗5号’。

图8 不同修剪频率下野生羊茅和猎狗5号各部位可溶性蛋白平均含量的变化Fig.8 Changes of soluble protein contents in different parts of Wild Festuca and ‘Houndog V’ under different trim frequencies
2.2.4脯氨酸的测定结果与分析
在不同修剪频率下,野生羊茅和‘猎狗5号’各部位脯氨酸平均含量的测定结果见表6。由表6计算可得,1次/10 d时,5次测定的野生羊茅地上部分脯氨酸平均含量的均值为83.958 ug·g-1,与1次/5 d时相比降低了15.14%;野生羊茅地下部分为38.053 ug·g-1,与1次/5 d时相比降低了8.88%;‘猎狗5号’地上部分为78.072 ug·g-1,与1次/5 d时相比降低了13.01%;‘’猎狗5号’地下部分为41.442 ug·g-1,与1次/5 d时相比降低了7.98%。1次/15 d下,5次测定的野生羊茅地上部分脯氨酸平均含量的均值为79.991 ug·g-1,与1次/5d时相比降低了19.15%;野生羊茅地下部分为35.745 ug·g-1,与1次/5 d时相比降低了14.41%;‘猎狗5号’地上部分为72.783 ug·g-1,与1次/5天时相比降低了18.90%;‘猎狗5号’地下部分为38.881 ug·g-1,与1次/5 d时相比降低了13.67%。

表6 不同修剪频率对脯氨酸平均含量的影响Table 6 Effect of different trim frequencies on Proline content

图9 不同修剪频率下野生羊茅和猎狗5号各部位脯氨酸平均含量的变化Fig.9 Changes of proline contents in different parts of Wild Festuca and Houndog V under different trim frequencies
不同修剪频率下两种草坪草各部分脯氨酸平均含量的变化趋势见图9。由表6和图9可以看出,两种草坪草地上部分的脯氨酸平均含量均比地下部分的脯氨酸平均含量高。随着修剪频率的降低,2种草坪草各部分的脯氨酸平均含量均逐渐减少,且降低幅度稍有减缓。2种草坪草地上部分的降低幅度均大于地下部分。其中在不同修剪频率下,野生羊茅地上部分和地下部分的降低幅度均略高于猎狗5号的相应部位。
由于修剪可以更好的促进草坪草的生长与分蘖,随着修剪频率的降低,草坪草的生长速度逐渐减缓,植物体内产生的脯氨酸等有机物质就会逐渐减少。而在不同修剪频率下,野生羊茅地上部分和地下部分的降低幅度均略高于‘猎狗5号’的相应部位,可能是由于修剪频率对野生羊茅的影响强于‘猎狗5号’。
3 讨论与结论
野生羊茅是呼和浩特市本土的野生品种,地区降水量少而不均、冬季漫长而寒冷、寒暑变化剧烈等特点使其具备了较好的抗寒性,因此可以考虑将其用作草坪草育种方向的研究进行改良优化,尝试作为呼和浩特市草坪草。但因其是野生品种,没有进行改良,其坪用性和耐修剪性能较差,所以后续应着重进行此方面的改良。
野生羊茅与‘猎狗5号’移栽组合的辛普森指数为0.500。其中,野生羊茅株数占组合总株数的比率为49%,‘猎狗5号’为51%。由此可见,在环境适应性假说下,2种草坪草占组合总株数的比率相差不大。
环境适应性假说处理下,5次测定的草坪草可溶性糖、可溶性蛋白及脯氨酸的平均含量变化趋势相似:5次测定下含量基本呈缓慢增加的趋势,且2种草坪草茎叶部分的含量均比根部的含量高;‘猎狗5号’的地上和地下部分的可溶性糖和可溶性蛋白含量均不同程度地高于野生羊茅对应部分。而脯氨酸含量地测定趋势稍有不同,野生羊茅地上部分的脯氨酸含量高于‘猎狗5号’地上部分,地下部分却低于‘猎狗5号’。
当修剪频率降低至1次/10 d时,各指数没有明显变化。而在1次/15 d时移栽组合的辛普森指数有所下降,说明此时组合的物种多样性和群落稳定性在逐渐下降。同时,野生羊茅的个体株数占组合总株数的比率有所下降,而‘猎狗5号’的个体株数占组合总株数的比率有所上升。因而,修剪频率的降低使得组合的物种多样性和群落稳定性逐渐下降,修剪频率对野生羊茅的影响大于‘猎狗5号’。
总体来说,随着修剪频率的逐渐降低,两种草坪草所测得的可溶性糖、可溶性蛋白及脯氨酸含量均逐渐降低。且2种草坪草地上部分的降低幅度均大于地下部分。其中野生羊茅地上和地下部分的降低幅度大体上均高于‘猎狗5号’的相应部分,即修剪频率对野生羊茅的影响大于‘猎狗5号’。
猜你喜欢
中老年保健(2022年3期)2022-11-21
求学·理科版(2020年10期)2020-10-20
临床检验杂志(电子版)(2020年1期)2020-04-03
幽默大师(2019年11期)2019-11-23
科学与财富(2019年15期)2019-10-21
新农民(2019年9期)2019-02-19
作文周刊·小学五年级版(2016年39期)2017-04-17
湖南林业科技(2017年1期)2017-02-06
唐山文学(2016年12期)2016-11-26
学苑创造·B版(2015年12期)2016-06-23
