
大麦近等基因系苗期根系形态及叶片渗透调节物质 对PEG胁迫的响应
2017-07-19 13:17杜欢马彤彤郭帅张颖白志英李存东
中国农业科学 2017年13期
杜欢,马彤彤,郭帅,张颖,白志英,李存东
大麦近等基因系苗期根系形态及叶片渗透调节物质 对PEG胁迫的响应
杜欢1,2,马彤彤1,2,郭帅2,3,张颖1,2,白志英1,2,李存东2
(1河北农业大学生命科学学院,河北保定 071001;2河北省作物生长调控实验室,河北保定 071001;3河北农业大学农学院,河北保定 071001)
【目的】研究大麦半矮秆基因与抗旱性的关系,为大麦抗旱优质育种提供理论依据。【方法】以两套大麦株高近等基因系(3T、3D和15T、15D)为试验材料,采用营养液水培方式,以不同浓度的PEG6000模拟干旱胁迫,研究不同胁迫程度:对照(0)、轻度(5%)、中度(15%)和重度胁迫(25%)对大麦株高近等基因系幼苗株高、根冠比、根系形态及叶片渗透调节物质的影响。在幼苗长至四叶一心时取样,直尺测定株高;采用烘干称重法获得根系生物量和地上部生物量,二者比值乘以100%得到根冠比;利用根系扫描仪对样根进行扫描测量,并用WinRHIZO软件进行单株总根长、根表面积和根体积的分析;可溶性糖含量用硫酸-蒽酮比色法测定;可溶性蛋白含量用考马斯亮蓝G-250染色法测定;脯氨酸含量用酸性茚三酮显色法测定。【结果】随着PEG6000浓度的增加,大麦幼苗生长受到明显抑制,株高逐渐降低,而根冠比则呈相反的变化趋势。就根系形态而言,随胁迫程度的增加,两套近等基因系表现出不同的变化趋势,在轻度和中度胁迫下3D和15T的变化幅度分别小于3T和15D,在重度胁迫下则表现相反。可溶性糖含量随胁迫浓度的增加呈先升高后降低再升高的变化趋势,而可溶性蛋白含量变化趋势与可溶性糖含量相反;在轻度和中度胁迫下3D和15D的变化幅度分别小于3T和15T,在重度胁迫下也表现相反。但脯氨酸含量随胁迫浓度升高呈现上升趋势,且在不同胁迫浓度下均表现为3D和15D变化幅度较小。相关分析表明:不同胁迫处理下,除可溶性蛋白含量外,株高与其他性状均存在显著相关关系;根冠比与根系形态以及根系形态之间也存在显著相关关系。同时,具有较高可溶性糖含量的品系其脯氨酸含量也较高,而可溶性蛋白含量则较低。此外,根系形态与叶片渗透调节物质之间亦存在一定的相关性,即总根长和根表面积均与可溶性糖含量和脯氨酸含量呈现显著负相关关系。【结论】在根系形态及叶片渗透调节方面,大麦株高近等基因系对苗期不同干旱胁迫的响应存在差异并相互影响。在轻度和中度胁迫程度下,近等基因系3T和3D中半矮秆基因的存在有利于抵御干旱胁迫。
大麦;株高近等基因系;PEG胁迫;根系形态;渗透调节
0 引言
【研究意义】大麦(L.)是中国主要种植作物之一,干旱严重影响了大麦的生长和产量,已成为制约农业生产和发展的主要限制因素之一,而生态环境的不断恶化和人为的气候改变又使干旱日趋严重[1-2]。相比于其他作物,大麦茎秆过高、易倒伏也是造成减产的主要原因。因此,研究大麦株高近等基因系根系及生理特性的抗旱性差异,分析根系形态与叶片渗透调节物质的相互关系,对于实现大麦降秆抗倒伏和抗旱优质育种具有重要意义。【前人研究进展】作物对干旱胁迫的响应包括形态和生理特征的变化,能够反映其本身的遗传多样性和抗旱性[3]。作物根系主要负责养分和水分的吸收并向地上部运输,它直接影响地上部的生长和营养状况及产量水平。干旱胁迫下,根系水分亏缺最终会导致作物产量降低[4],不同作物、不同遗传背景或不同水分胁迫程度,对根系生长和分布的影响均存在一定的差异[5-8]。惠宏杉等[9]研究认为,在干旱胁迫条件下抗旱性强的大麦品种能够保持更庞大的根系和更高的根系吸收能力。此外,作物耐旱的主要生理机制是通过渗透调节作用防止水分的过度流失,保持细胞膨压,从而保证植物的正常生长[10-13]。有机渗透调节物质主要包括脯氨酸、可溶性糖和可溶性蛋白等有机物质。有研究表明,抗旱性强的品种在低水势下能够维持一定的膨压,其渗透调节能力要高于抗旱性弱的品种[14]。汪军成等[15]研究表明,叶片脯氨酸含量和可溶性糖含量与干旱胁迫压力呈正相关。大麦株高近等基因系是通过重组自交得到的除株高不同,其他遗传背景完全相同的成套遗传品系。近年来,前人利用大麦株高近等基因系开展了大量研究,BAI等[16]研究了镰刀菌素冠腐病在大麦株高近等基因系之间的传播差异;杜欢等[17-18]研究了大麦株高近等基因系的籽粒性状、农艺性状及产量性状差异;CHEN等[19-20]研究了大麦株高近等基因系基于3H染色体长臂上影响穗形态的数量性状遗传位点和无赤霉酸响应的半矮秆基因对大麦镰刀菌素冠腐病抗性的影响。【本研究切入点】目前,关于不同干旱胁迫程度对大麦株高近等基因系根系形态和生理特性的影响,以及地下部根系形态与地上部生理特性的相互关系鲜有报道。【拟解决的关键问题】本研究以两套大麦株高近等基因系为试验材料,以期分析不同干旱胁迫程度下大麦株高近等基因系苗期根系形态及叶片渗透调节物质的抗旱性差异,揭示地下部根系形态与地上部渗透调节物质的相互关系,为大麦抗旱优质育种提供理论依据。
1 材料与方法
1.1 试验材料
以两套大麦株高近等基因系(3T与3D、15T与15D)为试验材料,由CSIRO Plant Industry, Australia提供;其供体亲本为TX9425(含半矮秆基因),受体亲本Franklin和Gairdner均不含半矮秆基因,亲本进行杂交得到的第二代群体TX9425/Franklin和TX9425/Gairdner分别进行自交授粉,筛选出第三代,再分别进行自交授粉,依次进行,直至产生的第8代筛选所得[19-20]。材料中T(Tall)代表无半矮秆基因的高秆基因系,D(Dwarf)代表含半矮秆基因的矮秆基因系。
1.2 试验设计
此外,以儿童在来生信念测量的总分为因变量,以2(父母宗教信仰组别)× 2(年龄:4~5岁、5~6岁)进行方差分析,结果发现,对于儿童来生信念测量的总分,年龄与父母宗教信仰组别交互作用不显著,父母宗教信仰组别(宗教信仰组:M=6.18,SD=3.61;无宗教信仰组:M=8.89,SD=2.98)主效应显著,F(1, 53)=8.20,p=0.006,η2=0.14。
试验于2015—2016年在河北农业大学试验站进行。采用营养液水培方式,塑料盆255 mm×190 mm×65 mm。选取饱满均一的大麦种子,自来水洗去尘土和杂质,0.1% HgCl2消毒15 min,蒸馏水漂洗数次,暗处萌发48 h后于光照培养箱中培养,每天用蒸馏水浇灌;7 d后,去除残粒,选择长势健壮一致的幼苗移入1/4霍格兰德营养液中,于光照室培养,昼夜温度24℃/17℃,光暗周期14 h/10 h,相对湿度50%,光照强度500 μmol·m-2,3 d后更换为1/2霍格兰德营养液,每3天通气1次;待幼苗长至两叶一心时,采用不同浓度的PEG6000(聚乙二醇6000)模拟干旱胁迫,分别为对照(0)、轻度(5%)、中度(15%)和重度(25%)胁迫;3盆重复,每盆50株,共计48盆;待幼苗长至四叶一心时取样测定相关指标[21-22]。
1.3 株高及根冠比的测定
学校每年会有20多项大大小小的学生活动,每项活动都经过精心设计,例如,春季远足,最远徒步20千米看家乡美景;初一年级诚信宣誓仪式,邀请家长和社会诚信典型观礼;初二年级离队入团仪式;18岁成人仪式,邀请法制副校长宣讲《宪法》;初高三年级毕业典礼,毕业生身穿毕业袍,校长以班级为单位签发毕业贺书;迎国庆升旗仪式;青春仪式等。这些活动,对学生影响巨大,留下终生难忘的回忆。通过这些活动,学生获得心灵的成长,人生也会树立更高的目标。
幼苗四叶一心时,取6株幼苗直尺测定株高,并将植株根系与地上部分离,分别于烘箱105℃杀青30 min,80℃烘干至恒重,分别记录根系生物量和地上部生物量,并计算根冠比(根系生物量/地上部生物量×100%),3次重复。
1.4 根系形态的测定
取各个处理下3株幼苗根系,采用EPSON PERFECTION V700 PHOTO彩色图像扫描仪进行根系扫描,并用WinRHIZO软件进行总根长、根表面积和根体积的分析,3次重复。
1.5 叶片渗透调节物质的测定
可溶性糖含量测定采用硫酸-蒽酮比色法[23];可溶性蛋白含量测定采用考马斯亮蓝G-250染色法[23];脯氨酸含量测定采用酸性茚三酮显色法[23]。
1.6 数据处理
由表1可知,不同的PEG胁迫浓度对大麦株高近等基因系株高和根冠比的影响差异不同。随PEG6000胁迫浓度的增加,两套近等基因系株高呈下降趋势且差异显著;在轻度、中度和重度干旱胁迫下,3T株高分别比对照降低了7.5%、25.6%和35.3%,3D株高分别比对照降低了7.1%、30.1%和40.2%,而15T株高分别比对照降低了8.9%、23.7%和34.7%,15D株高分别比对照降低了12.7%、35.1%和47.4%;其中,轻度胁迫下,株高的变化较小,随胁迫程度的增加株高下降幅度增大。由此可知,3T和15T随胁迫程度的增加株高降低程度较小。
采用Excel对数据进行整理分析和绘图,采用SPSS21.0软件[24]对数据进行方差分析和相关性分析。
2 结果
2.1 PEG胁迫对大麦株高近等基因系株高和根冠比的影响
在绿色建筑中,屋顶虽然不能够接受与外墙等同的太阳能,但是其采光性能要比天窗好很多。另外,由于屋顶平行于水平面,并且高于地面,阳光遮挡物较少,因此,在屋顶布局光伏发电系统无须考虑角度问题,只需要保证光电板最大面积覆盖即可。另外,由于屋顶独立于整个绿色建筑之上,容易遭受自然灾害的影响,其使用寿命最短,这些都是设计者需要考虑的问题[4]。
[8] 汪妮娜, 黄敏, 陈德威, 徐世宏, 韦善清, 江立庚. 不同生育期水分胁迫对水稻根系生长及产量的影响. 热带作物学报, 2013, 34(9): 1650-1656.
在全球经济一体化发展的背景下,我国企业逐步实现了与国际接轨的现代化发展,在引进新技术和新设备的同时,也需要建立新的现代化管理体制与新的服务平台。物流采购作为企业经营的重要组成部分,是增加企业的生产效益、提高企业在市场竞争中的占有率、促进企业现代化发展的重中之重,也是需要重点完善的内容。为了建立优质的物流采购管理平台,对物流采购流程等进一步优化,从而促进企业实现物流采购和管理的专业性、标准性,推动企业的信息化发展。
自金融危机爆发以来,各国经济得到了一定的复苏。但是,不能否认的是各国在消费市场、财经合作等方面依旧受到金融危机的影响而转为保守。此种保守的状态具体表现为两个方面:一是国与国之间加强了合作的同时在资源的争夺上也更为激烈,各国在原本“边境措施”的基础上,更加深入的扩展到对别国国内经济领域的渗透;二是各国在面对资本输出以及财经合作上更为的理性,尤其是在大型项目投资、风险资金对冲、虚拟经济以及金融经济管控上相对保守。此种趋势,在去年G20的过程中有所体现。国际社会将注意力集中在我国经济结构与经济发展方式的转变与调整上来,并促使了各国对于我国在税制、汇率、利率等多方面的改革与调整给予更大的希望。
两套近等基因系根冠比随胁迫浓度的增加呈不断上升的趋势。3T和15T根冠比在中度、重度胁迫下显著高于对照,分别比对照增加了42.9%、52.2%和67.5%、69.4%。在轻度、中度和重度胁迫时,15D根冠比均显著高于对照,分别比对照增加了115.8%、161.4%和165.8%;其他品系在不同干旱胁迫下与对照相比均无显著差异。由此可知,3T和15T随胁迫程度的增加根冠比变化幅度较小。

表1 不同PEG胁迫浓度对大麦株高近等基因系株高及根冠比的影响
同一行的不同小写字母代表同一品系不同胁迫浓度下性状值在0.05水平上差异显著
The different small letters after data in the same line mean the significant difference at the 0.05 level on the same variety under different stress concentration
2.2 PEG胁迫对大麦株高近等基因系根系形态的影响
2.2.1 总根长 由图1可知,3T和3D总根长随胁迫浓度的增加呈下降趋势;3T在轻度胁迫时总根长急剧降低,而3D在重度胁迫时总根长急剧降低,表明3T总根长对干旱胁迫比较敏感。在轻度、中度和重度胁迫时,3T总根长与对照相比差异显著,分别比对照降低了51.4%、58.7%和64.1%;而3D总根长只在重度胁迫下与对照相比差异显著,比对照降低66.7%。表明3D在轻度和中度胁迫下总根长变化幅度小于3T,而在重度胁迫下变化幅度大于3T。
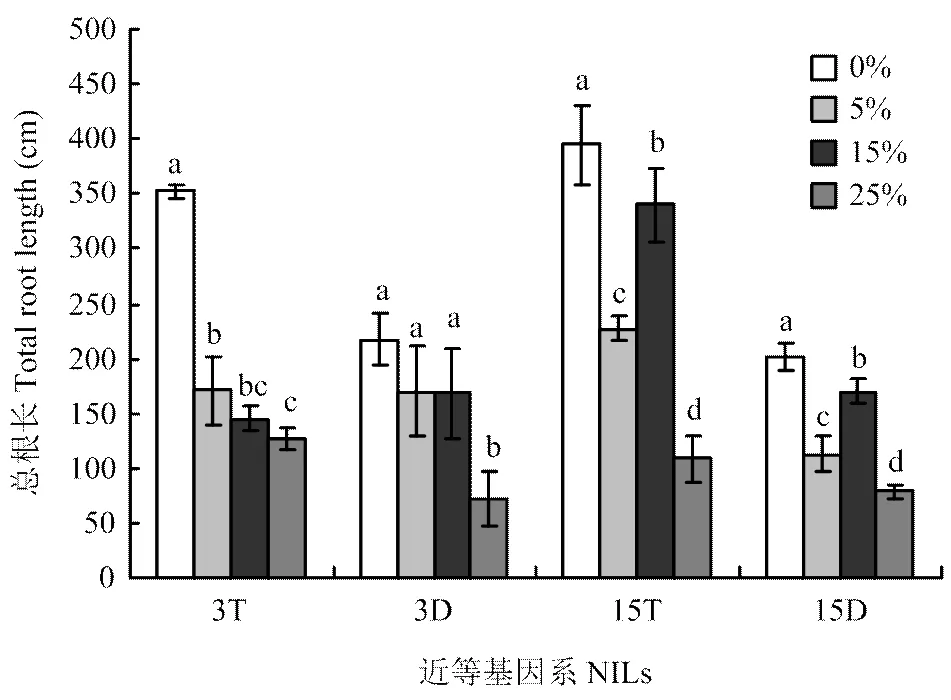
不同小写字母代表同一品系不同胁迫浓度下性状值在0.05水平上差异显著。下同
15T和15D总根长随胁迫浓度的增加呈先降低后升高再降低的变化趋势,并且均在中度胁迫时明显升高。在轻度、中度和重度胁迫时,15T和15D总根长分别与对照相比呈现显著差异;15T总根长分别比对照降低了42.4%、13.9%和72.5%;15D总根长分别比对照降低了44.1%、15.6%和61.1%。表明15T在轻度和中度胁迫下总根长变化幅度小于15D,而在重度胁迫下变化幅度大于15D。
2.2.2 根表面积 由图2可知,3T根表面积随胁迫浓度的增加呈下降趋势;3D根表面积随胁迫浓度的增加呈先降低后升高再降低的变化趋势,并在中度胁迫时升高。在轻度、中度和重度胁迫时,3T和3D根表面积分别与对照相比差异显著;3T根表面积分别比对照降低了50.4%、55.8%和60.3%;而3D根表面积分别比对照降低了37.3%、34.0%和68.4%。表明3D在轻度和中度胁迫下根表面积变化幅度小于3T,而在重度胁迫下表现相反。
[5] 丁红, 张智猛, 戴良香, 宋文武, 康涛, 慈敦伟. 不同抗旱性花生品种的根系形态发育及其对干旱胁迫的响应. 生态学报, 2013, 33(17): 5169-5176.
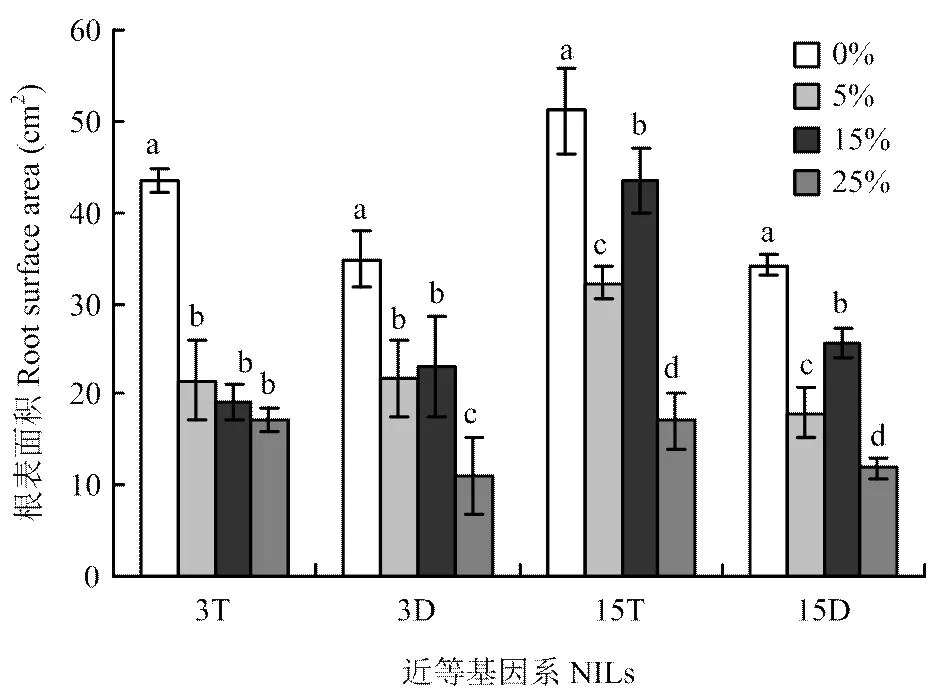
图2 不同PEG胁迫浓度对大麦株高近等基因系根表面积的影响
2.2.3 根体积 由图3可知,3T根体积随胁迫浓度的增加呈下降趋势;3D根体积随胁迫浓度的增加呈先降低后升高再降低的变化趋势,并在中度胁迫时升高。在轻度、中度和重度胁迫时,3T和3D根体积分别与对照相比差异显著;3T根体积分别比对照降低了49.5%、52.8%和56.2%;而3D根体积分别比对照降低了44.8%、39.1%和67.6%。表明3D在轻度和中度胁迫下根体积变化幅度小于3T,而在重度胁迫下表现相反。
15T和15D根体积随胁迫浓度的增加呈先降低后升高再降低的变化趋势,并且均在中度胁迫时升高。在轻度、中度和重度胁迫时,15T和15D根体积分别与对照相比呈现显著差异;15T根体积分别比对照降低了31.1%、18.5%和59.3%;15D根体积分别比对照降低了44.3%、24.6%和58.7%。表明15T在轻度和中度胁迫下根体积变化幅度小于15D,同样在重度胁迫下表现相反。

图3 不同PEG胁迫浓度对大麦株高近等基因系根体积的影响
2.3 PEG胁迫对大麦株高近等基因系叶片渗透调节物质的影响
2.3.1 可溶性糖含量 由图4可知,两套近等基因系可溶性糖含量均随胁迫浓度的增加呈先升高后降低再升高的变化趋势。3T和3D可溶性糖含量在轻度和重度胁迫下分别显著高于对照;3T分别比对照升高了529.7%和1001.4%;3D分别比对照升高了458.5%和1317.6%。而15T和15D在3种胁迫浓度下可溶性糖含量分别与对照相比均达显著水平;15T可溶性糖含量分别比对照升高82.6%、降低38.8%、升高81.0%;15D可溶性糖含量分别比对照升高14.5%、降低34.2%、升高112.2%。由此可知,在轻度和中度胁迫下3D和15D随胁迫程度的增加可溶性糖含量变化幅度小于3T和15T,而在重度胁迫下表现相反。

图4 不同PEG胁迫浓度对大麦株高近等基因系叶片可溶性糖含量的影响
2.3.2 可溶性蛋白含量 由图5可知,两套近等基因系可溶性蛋白含量均随胁迫浓度的增加呈先降低后升高再降低的变化趋势,且在中度胁迫时达到最大值。3T可溶性蛋白含量在轻度和中度胁迫下与对照相比差异显著,分别比对照降低16.7%和升高16.3%。3D可溶性蛋白含量在中度和重度胁迫下与对照相比差异显著,分别比对照升高13.7%和降低15.7%。此处,15T和15D在中度胁迫下可溶性蛋白含量与对照相比差异也均达显著水平,分别比对照升高了17.0%和16.3%。由此可知,在轻度和中度胁迫下3D和15D随胁迫程度的增加可溶性蛋白含量变化幅度小于3T和15T,同样在重度胁迫下表现相反。
[12] 王淑英, 姜小凤, 苏敏, 李倩. 水分胁迫对春小麦光合和渗透调节物质的影响. 麦类作物学报, 2013, 33(2): 364-367.
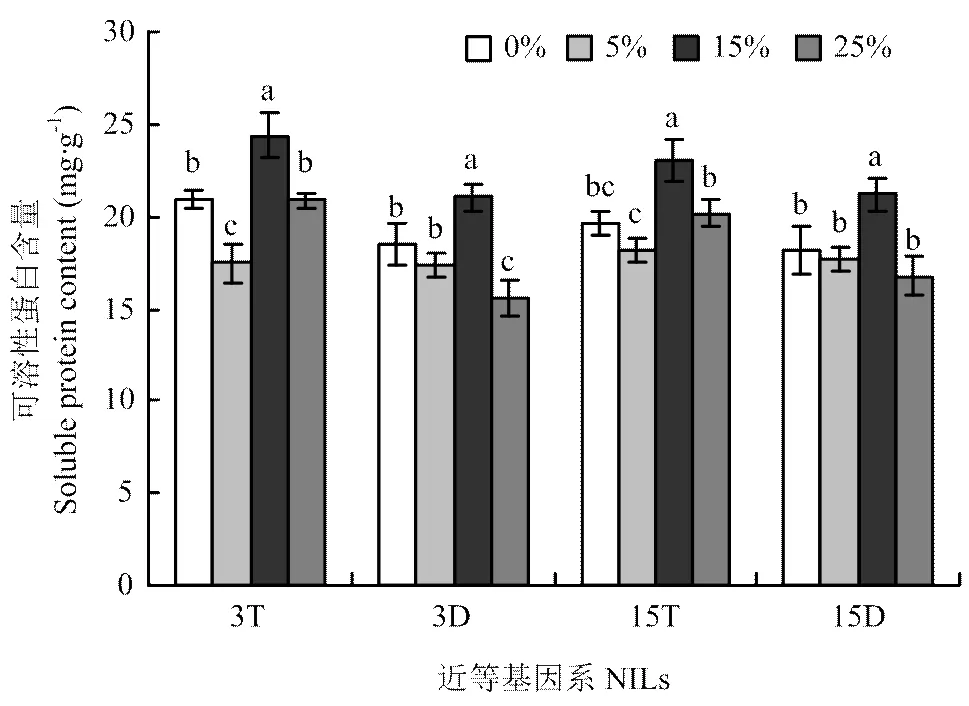
图5 不同PEG胁迫浓度对大麦株高近等基因系叶片可溶性蛋白含量的影响
2.4 各个性状间的相关性分析
由各个性状间的相关分析(表2)可知,不同PEG胁迫浓度处理下,株高与根冠比呈极显著负相关,与总根长、根表面积、根体积呈极显著正相关,与可溶性糖含量和脯氨酸含量呈显著负相关。根冠比与总根长呈显著负相关(-0.596*),与根表面积和根体积呈极显著负相关(-0.685**,-0.726**)。根系形态3个性状两两之间呈极显著正相关。同时,根系形态性状与叶片渗透调节物质之间亦存在一定的相关性,即总根长和根表面积均与可溶性糖含量和脯氨酸含量呈显著负相关,而根体积只与脯氨酸含量呈显著负相关。此外,具有较高可溶性糖含量的品系其脯氨酸含量也较高,而可溶性蛋白含量则较低,即前两者之间呈极显著正相关(0.640**),而可溶性糖含量与可溶性蛋白含量之间呈显著负相关(-0.592*)。
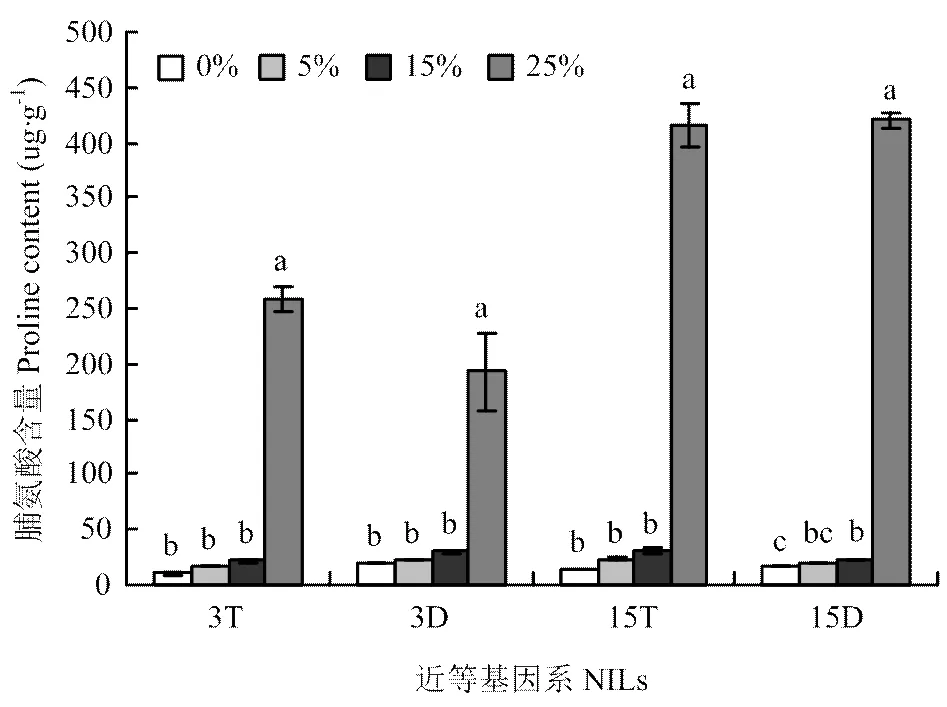
图6 不同PEG胁迫浓度对大麦株高近等基因系叶片脯氨酸含量的影响
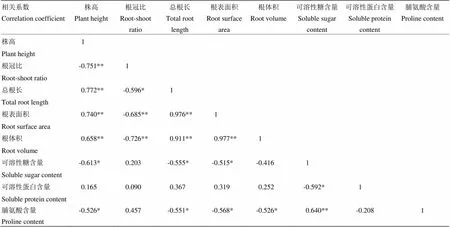
表2 各个性状间的相关性分析
*和**分别表示差异呈显著(<0.05)和极显著(<0.01)水平
*and** mean significant difference and positively significant difference at 0.05 and 0.01 level, respectively
3 讨论
3.1 大麦株高近等基因系株高和根冠比对PEG胁迫响应的差异
作物在水分胁迫状态下,表现为植株矮小、生长缓慢。但同样条件下,植株良好的根冠比能够使其将有限的资源进行最有效的分配,进而促进植株生长[25]。胡雯媚等[26]对小麦苗期的抗旱性进行了鉴定和指标筛选,指出株高和根冠比对小麦的抗旱性具有显著影响。袁蕊等[27]研究认为干旱胁迫导致小麦株高显著降低。惠宏杉等[9]研究表明抗旱性强的大麦品种根冠比随干旱强度的增加而上升且差异显著。本研究表明,两套近等基因系株高随胁迫浓度的增加呈下降趋势,而根冠比随胁迫浓度的增加呈不断上升趋势,这与前人研究结果较为一致;此外,3T和15T随胁迫程度的增加株高降低程度较小、根冠比增加幅度较小,即矮秆基因系对干旱胁迫更为敏感,表明近等基因系中半矮秆基因的存在会降低大麦抵御干旱胁迫的能力。
(2)改善钻井液润滑性。施工中通过加入1%油基润滑剂来优化钻井液润滑性,作用机理是在亲水性的井壁和钻具表面形成一层疏水性油膜,在增强吸附表面润滑性的同时,也有助于抑制泥页岩吸水膨胀分散,降低摩阻。若是摩阻较大的定向井,可向钻井液中加入10% 原油,并使其充分乳化,以改善钻井液的润滑性能和摩阻,阻止压差卡钻现象的发生[1]。
3.2 大麦株高近等基因系地下部根系形态对PEG胁迫响应的差异
作物根系的生长与遗传因素、生长环境和地上部的生长动态等密切相关[28]。根系的形态特征间接影响地上部植株的生长发育状况,从而影响作物产量。干旱条件下,较好的根系能够较大程度地减小产量损失[29]。据报道,同一小麦品种在不同干旱胁迫程度下,根长和根数存在较大差异;不同的小麦品种在相同的胁迫程度下,根长和根数特征亦存在较大差异[30]。有研究认为,干旱胁迫下冬小麦苗期根长和根体积明显下降[31]。丁红等[5]研究表明,抗旱型品种根系较为发达,通过增加根表面积和根体积等来适应水分胁迫。本研究表明,在中度胁迫程度下,3D、15T和15D根系生长较好;在轻度胁迫下,3T根系生长较好。在轻度和中度胁迫程度下,3D和15T抵御干旱胁迫的能力较强,在重度胁迫程度下表现相反。表明同样亲本重组自交得到的材料遗传背景差异较大,因而根系抵御干旱胁迫的能力有所不同。
3.3 大麦株高近等基因系地上部叶片渗透调节物质对PEG胁迫响应的差异
渗透调节是作物适应干旱胁迫的重要生理机制。植物在轻度或中度干旱胁迫下,细胞通过增加溶质来降低渗透势的主动调节作用[32];而干旱胁迫达到重度时,渗透调节作用减小甚至消失,说明植物的渗透调节能力是有一定限度的[33]。有研究表明,随土壤水分的亏缺,可溶性糖和可溶性蛋白含量呈先增后减趋势,而脯氨酸则呈逐渐增加的趋势[12]。蒋花等[34]认为在低浓度胁迫下,脯氨酸变化小、积累少,而在高浓度胁迫时,积累量增大;而可溶性糖含量随胁迫程度的增加变化趋势不同。本研究表明,4个品系在不同干旱胁迫程度下,单个渗透调节物质变化趋势一致;但在各个胁迫程度下与对照相比存在差异,这与前人研究结果并不完全一致,可能是材料遗传背景存在差异所致。在轻度和中度胁迫程度下,矮秆基因系抵御干旱胁迫的能力高于对应高秆基因系,说明半矮秆基因的存在有利于抵御干旱胁迫。而在重度胁迫程度下,近等基因系之间的差异有待进一步研究。
3.4 大麦株高近等基因系地下部与地上部生长的相关性
植株根系和地上部所处的环境及功能各不相同,但它们在营养物质的分配过程中相互依赖又相互影响。有研究表明,水分亏缺条件下,根系生长影响地上部生物量及作物产量[35]。本试验条件下,近等基因系株高与根系形态呈极显著正相关;叶片脯氨酸含量与根系形态均存在显著负相关关系;叶片可溶性糖含量与总根长和根表面积存在显著负相关关系;但可溶性蛋白含量与根系无显著相关性。刘胜群等[36]研究认为根系与可溶性蛋白和可溶性糖含量均呈显著正相关,本试验结果与其相反,可能是由于材料差异以及胁迫处理不同所致。干旱胁迫对根系形态产生直接影响,导致营养物质运至地上部受到抑制,间接影响叶片渗透调节物质的变化。因此,后续还应对根系渗透调节物质的变化以及影响地上部的途径进行研究,从而阐明大麦株高近等基因系地上部及地下部相互作用的机理。同时,近等基因系的综合抗旱能力以及全生育时期的抗旱情况仍需进一步研究
4 结论
干旱胁迫条件下,大麦株高近等基因系根系形态的变化影响叶片渗透调节物质的改变,进而影响植株的生长发育。在根系形态及叶片渗透调节方面,大麦株高近等基因系对苗期不同干旱胁迫的响应存在差异并相互影响。在轻度和中度胁迫程度下,近等基因系3T和3D中半矮秆基因的存在有利于抵御干旱胁迫,也有利于实现降秆抗倒伏的同时减少干旱对大麦苗期产生的影响。
运用战略管理的思想,根据高校自身的发展目标,对资产管理工作进行准确的定位,统筹资产管理工作,制定详细可行的资产管理规划。坚持勤俭办学,科学决策,优化资源,提升学校的核心竞争力以及可持续发展的能力。
References
ZHANG Z M, DAI L X, SONG W W, DING H, CI D W, KANG T, NING T Y, WAN S B. Effect of drought stresses at different growth stages on peanut leaf protective enzyme activities and osmoregulation substances content., 2013, 39(1): 133-141. (in Chinese)
[2] 丁红, 张智猛, 戴良香, 慈敦伟, 秦斐斐, 宋文武, 刘孟娟, 付晓. 水分胁迫和氮肥对花生根系形态发育及叶片生理活性的影响. 应用生态学报, 2015, 26(2): 450-456.
DING H, ZHANG Z M, DAI L X, CI D W, QIN F F, SONG W W, LIU M J, FU X. Effects of water stress and nitrogen fertilization on peanut root morphological development and leaf physiological activities., 2015, 26(2): 450-456. (in Chinese)
[3] NEVO E, CHEN G. Drought and salt tolerances in wild relatives for wheat and barley improvement., 2010, 33(4): 670-685.
[4] SALEKDEH G H, REYNOLDS M, BENNETT J, BOYER J. Conceptual framework for drought phenotyping during molecular breeding., 2009, 14(9): 488-496.
15T和15D根表面积随胁迫浓度的增加呈先降低后升高再降低的变化趋势,并且均在中度胁迫时升高。在轻度、中度和重度胁迫时,15T和15D根表面积分别与对照相比呈现显著差异;15T根表面积分别比对照降低了37.0%、15.3%和66.6%;15D根表面积分别比对照降低了47.3%、24.8%和65.3%。表明15T在轻度和中度胁迫下根表面积变化幅度小于15D,同样在重度胁迫下表现相反。
DING H, ZHANG Z M, DAI L X, SONG W W, KANG T, CI D W. Responses of root morphology of peanut varieties differing in drought tolerance to water-deficient stress., 2013, 33(17): 5169-5176. (in Chinese)
[6] 厉广辉, 万勇善, 刘风珍, 张昆. 不同抗旱性花生品种根系形态及生理特性. 作物学报, 2014, 40(3): 531-541.
LI G H, WAN Y S, LIU F Z, ZHANG K. Morphological and physiological traits of root in different drought resistant peanut cultivars., 2014, 40(3): 531-541. (in Chinese)
[7] 王稼苜, 张志勇, 欧行奇, 胡杨. PEG胁迫对8个不同小麦品种幼苗根系的影响. 河南科技学院学报(自然科学版), 2015, 43(3): 1-5.
WANG J M, ZHANG Z Y, OU X Q, HU Y. Effect of water stress by PEG on the seedling root characteristics of 8 wheat varieties., 2015, 43(3): 1-5. (in Chinese)
看到这些野生的,我连忙说,姐,现在能吃到纯野生的鲫鱼,真是福分啊!不料,姐姐有些忧伤地说,这可能是最后一次送野生鲫鱼给你们吃了,过不了几天,水库就要重新承包给其他人了,听说,他们为了获取更大利益,要用饲料养鱼了。那时,再也没有这活蹦乱跳,味道鲜美的野生鲫鱼了。
WANG N N, HUANG M, CHEN D W, XU S H, WEI S Q, JIANG L G. Effects of water stress on root and yield of rice at different growth stages., 2013, 34(9): 1650-1656. (in Chinese)
[9] 惠宏杉, 林立昊, 齐军仓, 廖乐, 王超龙, 程海涛. 干旱胁迫对大麦幼苗根系的影响. 麦类作物学报, 2015, 35(9): 1291-1297.
HUI H S, LIN L H, QI J C, LIAO L, WANG C L, CHENG H T. Effect of drought stress on the roots of barley seedling., 2015, 35(9): 1291-1297. (in Chinese)
观察两组治疗方案安全性、治疗效果及UPDRS评分情况。采用UPDRS(运动功能障碍评估表)对患者运动障碍情况进行评估,分数越高,代表患者运动功能障碍越严重;治疗总有效率等于有效率加上显效率,无效标准为:患者治疗后UPDRS差比百分率下降幅度低于5%;显效标准为:患者治疗后UPDRS差比百分率下降程度在20%~49%之间;显效标准为:患者治疗后UPDRS差比百分率下降程度在50%~99%之间。
[10] 张智猛, 戴良香, 宋文武, 丁红, 慈敦伟, 康涛, 宁唐原, 万书波. 干旱处理对花生品种叶片保护酶活性和渗透物质含量的影响. 作物学报, 2013, 39(1): 133-141.
[1] WENDELBOE-NELSON C, MORRIS P C. Proteins linked to drought tolerance revealed by DIGE analysis of drought resistant and susceptible barley varieties., 2012, 12(22): 3374-3385.
[11] 方立锋, 丁在松, 赵明. 转ppc基因水稻苗期抗旱特性研究. 作物学报, 2008, 34(7): 1220-1226.
FANG L F, DING Z S, ZHAO M. Characteristics of drought tolerance in ppc over expressed rice seedlings., 2008, 34(7): 1220-1226. (in Chinese)
妈妈一听我要去旅行,直接把筷子摔在桌子上,她指着我说:“你也是三十岁的人了,怎么这么没有心?怎么这么不懂事?”
2.3.3 脯氨酸含量 由图6可知,两套近等基因系脯氨酸含量均随胁迫浓度的增加呈逐渐升高的变化趋势,且在轻度和中度胁迫时增加缓慢,在重度胁迫时急剧增加。两套近等基因系脯氨酸含量均在重度胁迫下分别与对照相比差异显著,在此胁迫浓度下3T、3D、15T和15D分别比对照增加了2401.6%、872.5%、2663.9%和2381.1%。同时,15D脯氨酸含量在中度胁迫下与对照相比差异也达显著水平,比对照升高36.3%。由此可知,在3种胁迫浓度下3D和15D随胁迫程度的增加脯氨酸含量变化幅度小于3T和15T。
WANG S Y, JIANG X F, SU M, LI Q. Effect of water stress on photosynthetic parameters and osmotic regulation substances of spring wheat., 2013, 33(2): 364-367. (in Chinese)
[13] 张维军, 袁汉民, 陈东升, 王小亮, 亢玲, 何进尚. 小麦抗旱性生理生化机制及QTL研究进展. 干旱地区农业研究, 2015, 33(6): 139-148.
ZHANG W J, YUAN H M, CHEN D S, WANG X L, KANG L, HE J S. Advances in studies on physiological and biochemical indexes and QTL for drought resistance in wheat., 2015, 33(6): 139-148. (in Chinese)
[14] 李永华, 王玮, 马千全, 邹琦珑. 干旱胁迫下抗旱高产小麦新品系旱丰9703的渗透调节与光合特性. 作物学报, 2003, 29(5): 759-764.
(1)随着冲击能量的增大,弹性地基中位移影响范围逐渐扩大,由板底部2/3区域(图3a);逐渐扩大到板底整个区域(图3b);直到超过板底的区域范围(图3c)。
LI Y H, WANG W, MA Q Q, ZOU Q L. The osmotic adjustment and photosynthesis of a wheat cultivar hanfeng 9703 with high yield drought resistance under drought stress., 2003, 29(5): 759-764. (in Chinese)
[15] 汪军成, 孟亚雄, 徐先良, 王晋, 赖勇, 李葆春, 马小乐, 王化俊. 大麦苗期抗旱性鉴定及评价. 干旱地区农业研究, 2013, 31(4): 135-143.
WANG J C, MENG Y X, XU X L, WANG J, LAI Y, LI B C, MA X L, WANG H J. Identification and assessment on drought-resistance ofL. at seedling stage., 2013, 31(4): 135-143. (in Chinese)
[16] BAI Z Y, LIU C J. Histological evidence for different spread of Fusarium crown rot in barley genotypes with different heights., 2015, 163(2): 91-97.
黄庭坚爱茶,爱的是茶的文化属性。俗人饮酒,雅士品茗。他是在清茶一杯中,参透了茶中渗透的自然天成之性,养生保健之功,承载礼雅平和,沟通世界之仪的礼乐内韵。他是在茶读伴思清的功用中,以茶会友,茶悟人生;以茶修身,致清导和;以茶励志,审己自省;以茶倡廉,构建和谐;因茶完成修身治平的人格与道德的和美仪式,与茶达到人文追求的高度契合。这一切,都是山谷从中国传统儒道释文化中悟化而来,是茶道最高的追求。
[17] 杜欢, 张颖, 薛梦瑶, 靖姣姣, 白志英, 李存东. 大麦株高近等基因系的籽粒性状差异及相关性分析. 华北农学报, 2015, 30(5): 97-103.
DU H, ZHANG Y, XUE M Y, JING J J, BAI Z Y, LI C D. Difference and correlation analysis of grain traits in the near-isogenic line of plant height of barley., 2015, 30(5): 97-103. (in Chinese)
[18] 杜欢, 马彤彤, 侯晓梦, 张颖, 白志英, 李存东. 20对大麦株高近等基因系农艺与产量性状差异及相关性分析. 华北农学报, 2016, 31(5): 114-121.
DU H, MA T T, HOU X M, ZHANG Y, BAI Z Y, LI C D. Difference and correlation analysis of agronomic and yield characters in twenty pairs near-isogenic line of plant height of barley., 2016, 31(5): 114-121. (in Chinese)
[19] CHEN G D, LI H B, ZHENG Z, WEI Y M, ZHENG Y L, MCLNTYRE C L, ZHOU M X, LIU C J. Characterization of a QTL affecting spike morphology on the long arm of chromosome 3H in barley (L.) based on near isogenic lines and a NIL-derived population., 2012, 125(7): 1385-1392.
[20] CHEN G D, YAN W, LIU Y X, WEI Y M, ZHOU M X, ZHENG Y L, MANNERS J M, LIU C J. The non-gibberellic acid-responsive semi-dwarfing geneaffects Fusarium crown rot resistance in barley., 2014, 14(1): 1-8.
[21] 米少艳, 路斌, 张颖, 杜欢, 白志英, 李存东. 小麦代换系幼苗根系对低磷胁迫的生理响应暗示的染色体效应. 植物营养与肥料学报, 2016, 22(5): 1204-1211.
MI S Y, LU B, ZHANG Y, DU H, BAI Z Y, LI C D. Chromosome control implied by the physiological responses of root in wheat substitution lines seedlings under phosphorus deficiency stress., 2016, 22(5): 1204-1211. (in Chinese)
用具有一定厚度内壁的没有缝隙的钢管,刚度和强度较强的接头管并且保持关内的直径总体小于墙体的厚度10到20毫米。如果直径低于这个范围则采用30-50t的起重机进行设备的起拔,如果接头管的内直径厚度大于20毫米就需要用到专业的更大的起重机,以保障最终接头管的接头工作顺利进行。整体的起拔工作过程中,为使得工作在限制的时间内完成,需要保证接头管起拔工作在初凝之前完工。如果碰上混凝土的出现接头管断裂现象,依照高压喷射所具备的特性可以及时的对断裂处进行补救。
[22] 靖姣姣, 张颖, 白志英, 李存东. 盐胁迫对小麦代换系渗透调节物质的影响及染色体效应. 植物遗传资源学报, 2015, 16(4): 743-750.
JING J J, ZHANG Y, BAI Z Y, LI C D. The effects of salt stress on osmoregulation substance and chromosome of wheat substitution lines., 2015, 16(4): 743-750.(in Chinese)
[23] 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2000: 111-162.
ZOU Q.. Beijing: China Agriculture Press, 2000: 111-162. (in Chinese)
[24] 张力. SPSS19.0在生物统计中的应用. 厦门: 厦门大学出版社, 2013.
ZHANG L.. Xiamen: Xiamen University Press, 2013. (in Chinese)
[25] MAGNANI F, MENCUCCINI M, GRACE J. Age-related decline in stand productivity: the role of structural acclimation under hydraulic constraints., 2000, 23(3): 251-263.
[26] 胡雯媚, 王思宇, 樊高琼, 刘运军, 郑文, 王强生, 马宏亮. 西南麦区小麦品种苗期抗旱性鉴定及其指标筛选. 麦类作物学报, 2016, 36(2): 182-189.
HU W M, WANG S Y, FAN G Q, LIU Y J, ZHENG W, WANG Q S, MA H L. Analysis on the drought resistance and screening of drought resistance appraisal indexes of wheat cultivars in seedling stage in southwest area., 2016, 36(2): 182-189. (in Chinese)
[27] 袁蕊, 李萍, 胡晓雪, 宗毓铮, 孙敏, 董琦, 郝兴宇. 干旱胁迫对小麦生理特性及产量的影响. 山西农业科学, 2016, 44(10): 1446-1449.
YUAN R, LI P, HU X X, ZONG Y Z, SUN M, DONG Q, HAO X Y. Effects of drought stress on physiological characteristics and yield of wheat., 2016, 44(10): 1446-1449. (in Chinese)
[28] 邱新强, 高阳, 黄玲, 李新强, 孙景生, 段爱旺. 冬小麦根系形态性状及分布. 中国农业科学, 2013, 46(11): 2211-2219.
QIU X Q, GAO Y, HUANG L, LI X Q, SUN J S, DUAN A W. Temporal and spatial distribution of root morphology of winter wheat., 2013, 46(11): 2211-2219. (in Chinese)
[29] KOOLACHART R, JOGLOY S, VORASOOT N, WONGKAEW S, HOLBROOK C C, JONGRUNGKLANG N, KESMALA T, PATANOTHAI A. Rooting traits of peanut genotypes with different yield responses to terminal drought., 2013, 149: 366-378.
[30] 王瑾, 刘桂茹, 杨学举. PEG胁迫下小麦再生植株根系特性与抗旱性的关系. 麦类作物学报, 2006, 26(3): 117-119.
WANG J, LIU G R, YANG X J. Relationships between root characters and drought tolerance of regenerated plantlets of wheat under PEG water-stress., 2006, 26(3): 117-119. (in Chinese)
[31] SHAN C J, LIANG Z S. Effects of soil drought on root growth and physiological characteristics of winter wheat seedlings., 2007, 15(5): 38-41.
[32] 魏志刚, 王玉成. 植物干旱胁迫响应机制. 北京: 科学出版社, 2015: 34-47.
WEI Z G, WANG Y C.. Beijing: Science Press, 2015: 34-47. (in Chinese)
[33] 李吉跃. 植物耐旱性及其机理. 北京林业大学学报, 1991, 13(3): 92-97.
LI J Y. Plant drought resistance and its mechanism., 1991, 13(3): 92-97. (in Chinese)
[34] 蒋花, 王占红, 张小燕. PEG渗透胁迫下3份大麦材料幼苗叶片抗旱生理特性分析. 干旱地区农业研究, 2011, 29(5): 100-105.
JIANG H, WANG Z H, ZHANG X Y. Analysis on drought resistance of leaves of barley seedling under the condition of PEG osmotic stress., 2011, 29(5): 100-105. (in Chinese)
[35] 杨彩玲, 刘立龙, TIEN C N, 秦华东, 赵泉, 陈德威, 徐世宏, 黄敏, 江立庚. 土壤水分对免耕水稻根系生长的影响. 华中农业大学学报(自然科学版), 2016, 35(1): 8-16.
YANG C L, LIU L L, TIEN C N, QIN H D, ZHAO Q, CHEN D W, XU S H, HUANG M, JIANG L G. Soil moisture on the effects of no-tillage rice root growth., 2016, 35(1): 8-16. (in Chinese)
[36] 刘胜群, 宋凤斌, 王燕. 玉米根系性状与地上部性状的相关性研究. 吉林农业大学学报, 2007, 29(1): 1-6.
LIU S Q, SONG F B, WANG Y. Correlations between characters of roots and those of aerial parts of maize varieties., 2007, 29(1): 1-6. (in Chinese)
(责任编辑 李莉)
Response of Root Morphology and Leaf Osmoregulation Substances of Seedling in Barley Genotypes with Different Heights to PEG Stress
DU Huan1,2, MA TongTong1,2, GUO Shuai2,3, ZHANG Ying1,2, BAI ZhiYing1,2, LI CunDong2
(1College of Life Sciences, Hebei Agricultural University, Baoding 071001, Hebei;2Key Laboratory of Hebei Crop Growth Regulation, Baoding 071001, Hebei;3College of Agronomy, Hebei Agricultural University, Baoding 071001, Hebei)
【Objective】The aim of this study was to assess the relationship between the semi-dwarf geneof barley and the drought resistance and to provide a theoretical basis for drought resistance and high quality breeding of barley.【Method】A hydroponic experiment was carried out to study the plant height, root-shoot ratio, root morphology and leaf osmoregulation substances of seedling of two pairs barley genotypes with different heights under different drought stress levels resulted by PEG6000 at different concentrations (0, 5%, 15%, 25%). Samples were taken during seedlings grew to four leaves and a heart. Plant height was measured by ruler. Root-shoot ratio was calculated by root biomass to aboveground biomass ratio multiplied by 100%, which were obtained by drying and weighing. Total root length, root surface area and volume were measured and analyzed by root system scanner and the WinRHIZO software. The soluble sugar, soluble protein and proline content were determined by sulfuric acid-anthrone colorimetry, coomassie brilliant blue G-250 dyeing method and acidic indene three ketone chromogenic method, respectively.【Result】Analysis showed that barley seedling growth was significantly inhibited, and the plant height decreased gradually, while the opposite trend showed in the root-shoot ratio with the increase of stress concentration. In terms of root morphology, as the stress concentration increased, two pairs near-isogenic line showed different trends. The change range of 3D and 15T was less than that of 3T and 15D, respectively, under the mild and moderates tress, while the change range of 3D and 15T showed an opposite trend under severe stress. Furthermore, soluble sugar content showed an increase at first and then a decrease and an increase again with the increase of the stress concentration, while the opposite trend showed in soluble protein content. The change range was characterized by 3D<3T, 15D<15T under the mild and moderate stress, and the change range of 3D and 15D also showed an opposite trend under severe stress. But the proline content presented a tendency of increasing with the increase of the stress concentration, and the small variations were characterized in 3D and 15D under different treatments. Correlation analysis indicated that plant height showed significant correlation with other traits in addition to the soluble protein content. there was also a significant relationship between root-shoot ratio and root morphology under different treatments. Meanwhile, the strains with higher soluble sugar content also had higher proline content, while the soluble protein content was low. Moreover, root morphology also had certain correlation with leaf osmoregulation substances, namely, the total root length and root surface area had significant negative relationship with soluble sugar content and proline content, respectively.【Conclusion】In the matter of root morphology and leaf osmoregulation, there are different responses under drought stress and influence with each other at seedling stage of barley genotypes with different heights. Under the mild and moderate stress, the semi-dwarf gene in the near-isogenic lines 3T and 3D can resist drought stress.
barley; near-isogenic line of plant height; PEG stress; root morphology; osmoregulation
2016-12-07;接受日期:2017-02-07
河北省自然科学基金(C2011204016,C2015204066)
杜欢,E-mail:huan741858931@126.com。通信作者白志英,E-mail:zhiyingbai@126.com。通信作者李存东,E-mail:nxylcd@ mail.hebau.edu.cn
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
小小说月刊·下半月(2021年8期)2021-09-05
疯狂英语·新读写(2021年2期)2021-02-25
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
动漫界·幼教365(大班)(2018年3期)2018-05-14
现代农业科技(2017年16期)2017-09-22
湖南大学学报·自然科学版(2016年12期)2017-05-12
江苏农业科学(2016年4期)2016-06-14
