
温度、体重对斑石鲷耗氧率、排氨率的影响及昼夜节律变化❋
2017-07-17 08:27宋协法程亚伟邢道超翟介明董登攀
中国海洋大学学报(自然科学版) 2017年9期
宋协法, 程亚伟, 邢道超,彭 磊, 翟介明, 李 波, 董登攀
(1.中国海洋大学水产学院,山东 青岛 266003; 2.山东莱州明波水产有限公司,山东 烟台 261418)
温度、体重对斑石鲷耗氧率、排氨率的影响及昼夜节律变化❋
宋协法1, 程亚伟1, 邢道超1,彭 磊1, 翟介明2, 李 波2, 董登攀1
(1.中国海洋大学水产学院,山东 青岛 266003; 2.山东莱州明波水产有限公司,山东 烟台 261418)
采用密闭流水式呼吸实验方法,研究了不同体重组((50.94±5.50) g、(103.61±10.48) g、(154.58±7.56) g、(194.56±14.56) g)斑石鲷(Oplegnathuspunctatus) 在3个不同温度(20、25和30℃)下的耗氧率和排氨率,以及斑石鲷耗氧率、排氨率的昼夜节律变化。研究表明,3个温度条件下斑石鲷耗氧率和排氨率随温度的升高而升高,随体重的增大而降低,斑石鲷白天平均耗氧率为(2.535±0.204) μg·(g·min)-1,夜间平均耗氧率为(2.531±0.064) μg·(g·min)-1,其耗氧率的昼夜差异不明显。白天平均排氨率为(0.104±0.007) μg·(g·min)-1,夜间平均排氨率为(0.083±0.007) μg·(g·min)-1,白天平均排氨率明显大于夜间。投喂饲料后斑石鲷耗氧率最高点出现在投喂饲料后1~3 h,最低点出现在投喂饲料后19~23 h,耗氧率呈逐渐降低的趋势,排氨率呈先升高后降低的趋势,温度升高使斑石鲷排氨率达到峰值的时间提前。
斑石鲷;温度;体重;耗氧率;排氨率;昼夜节律
斑石鲷(Oplegnathuspunctatus)属鲈形目(Perciformes)石鲷科(Oplegnathidae)石鲷属(Oplegnathus),俗称黑金鼓、斑鲷。主要分布于中国和日本的温热带海域,是目前新兴的水产品种,具有重要的市场前景和价值[1]。国内在斑石鲷人工育苗和养殖方面起步较晚,目前对斑石鲷的研究主要集中在早期发育阶段的发育生物学方面,对斑石鲷的呼吸和排泄等新陈代谢的研究尚属空白。
呼吸和排泄是鱼类新陈代谢活动的重要生理指标[2],研究环境因素对鱼类代谢率的影响是研究鱼类生物能量学的重要内容。了解鱼类的耗氧及排氨能力能直接或间接地反映其代谢规律、特点以及生理、生存状况,其中氧氮比(O∶N)(蛋白质供能比值)能够反映生物代谢中能源物质的化学本质, 氧氮比(O∶N)的大小表示蛋白质与脂肪和碳水化合物提供给生物体能量的比率,Q10(温度系数)作为温度变化对鱼类代谢影响的指标之一,反应了生物体内反应速率和温度的关系[3]。斑石鲷代谢率的研究对其能量学研究及饲料营养配制具有更直接的指导作用。目前对于鱼类呼吸代谢的研究报道很多,如侯晓飞[4]对许氏平鮋(Sebastesschlegeli)氮磷排泄的研究,王刚等[5-6]研究了温度、盐度、pH、流速、体重和放养密度等环境因子对卵形鲳鲹(Trachinomsovatus)代谢的影响,闫茂仓等[3]研究了温度、盐度和体重对条石鲷(Oplegnathusfasciatus)耗氧率和排氨率的影响。本研究探讨了不同温度对不同体重的斑石鲷耗氧率和排氨率的影响及斑石鲷代谢率的昼夜节律变化,旨在为斑石鲷循环水养殖过程中的水质调控、鲜活运输以及饲料营养配制提供理论依据。
1 材料与方法
1.1 实验时间和条件
实验于2016年4—5月在莱州明波水产有限公司进行,实验用水为经过沉淀、砂滤后的自然海水和电厂热水勾兑。自然海水温度为(20±1)~(23±1) ℃,电厂热水温度为(32±2) ℃,盐度32,pH=7.5±0.2。
1.2 实验用鱼
实验用鱼为人工苗种,来自莱州明波水产有限公司,体重50~200 g,选择体色正常、无病无伤、行动迅速、生命力强的个体作为实验用鱼。实验前于实验鱼池内驯养7 d,待其状态稳定摄食正常后开始实验。实验鱼的初始体重分组如表1所示。
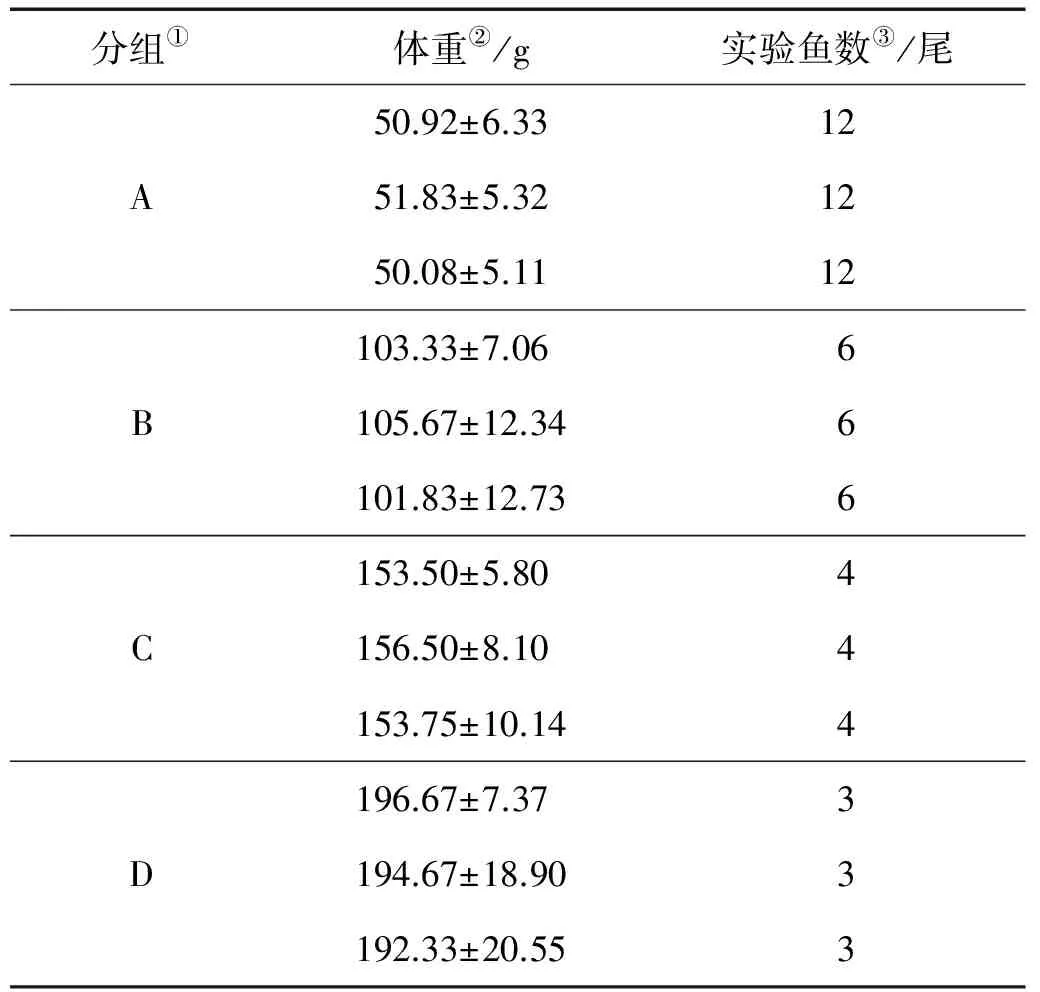
表1 实验鱼的初始体重分组
Note:①Group;②Body weight;③Number of samples
1.3 实验设计
实验采用密闭流水式实验方法,实验装置示意图见图1。呼吸室为玻璃钢鱼池(120 L),在鱼池上方加设透明有机玻璃,密封处理。设置1个高位蓄水池平衡实验水压,保持水流稳定(流量控制在1 500 mL/min左右,以避免水体缺氧和氨氮积累),水池内放置一支温度计随时记录温度,蓄水池内充氧。如图2所示,养殖池设4个体重组,每组设3个平行,加2个空白对照(呼吸室无鱼)。实验开始前每天升高1 ℃逐渐将水温调至实验温度。每日7:00、16:00各投喂一次,投喂饲料后半小时清除桶底残饵粪便。实验开始前逐尾称量体重。
实验当天7:00投喂饲料,半小时后清除残饵粪便,用量筒于出水口测量流量,8:00开始每2 h对养殖池进出水口溶氧和氨氮测定一次,共12次。溶氧的测定采用AZ-8403型溶氧仪(实验前与碘量法GB 17378.4-2007进行对比测定,并对溶氧仪进行盐度系数校正),氨氮测定采用次溴酸盐氧化法(GB 17378.4-2007)。根据进出水口溶氧和氨氮的变化计算斑石鲷耗氧率和排氨率。次日不投饵,8:00开始每4 h对养殖池进出水口溶氧和氨氮测定一次,根据进出水口溶氧和氨氮的变化计算斑石鲷耗氧率和排氨率。每个温度梯度实验做完后正常投饵,每天升温1 ℃逐渐升温至下一个温度梯度,斑石鲷的运动和摄食正常后重复以上实验。

(1.自然海水;2.电厂热水;3.溢流口;4.增氧气盘;5.蓄水池;6.鱼池进水;7.密封盖;8.鱼池出水;9.养殖鱼池;10.排污口。1.Sea water;2.Hot water(power plant);3.Overflow port;4.Aerobic plate;5.Reservoir;6.Inlet;7.Sealing cover;8.Outlet;9.Tank;10.Sewage outlet.)
图1 实验装置示意图
Fig.1 Schematic diagram of experimental device
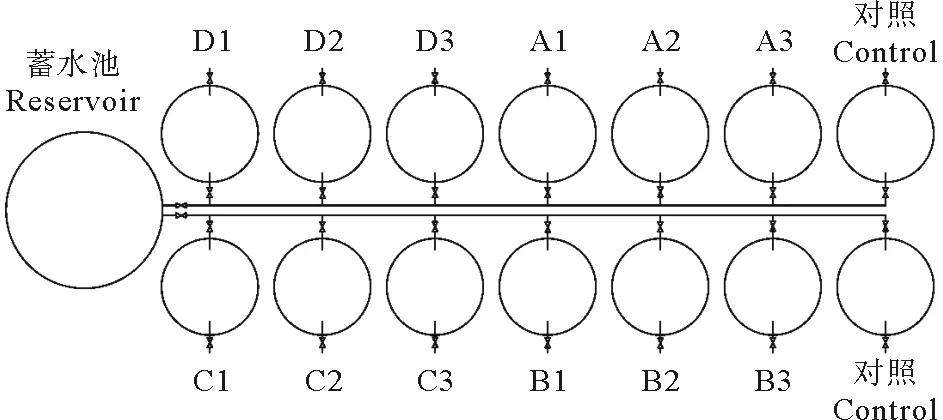
图2 实验装置平面图
1.4 数据计算和处理
耗氧率和排氨率的计算公式如下
RO=(CDO0-CDO1)×Q/W,
(1)
RN=(CN1-CN0)×Q/W。
(2)
式中:RO为耗氧率( μg·(g·min)-1);CDO0为流入呼吸室的海水溶氧量(mg/L);CDO1为流出呼吸室的海水溶氧量(mg/L);RN为排氨率( μg·(g·min)-1);CN0为流入呼吸室的海水氨氮浓度(mg/L);CN1为流出呼吸室的海水氨氮浓度(mg/L);Q为海水经过呼吸室的水
流量(mL/min);W为实验鱼体重(g)。
温度系数(Q10)表示温度每升高10 ℃动物的呼吸或排泄的变化比率,温度系数的计算公式为
Q10=(R2/R1)10/(T2-T1)。
(3)
氧氮比(O∶N)为耗氧率和排氨率的克原子之比,氧氮比的计算公式为
O∶N=RO/RN。
(4)
2 结果与分析
2.1 温度和体重对斑石鲷耗氧率排氨率的影响
2.1.1 温度对斑石鲷耗氧率、排氨率的影响 4组体重下斑石鲷耗氧率在不同温度条件下的变化如图3所示,在3个温度条件下,随着温度的升高,斑石鲷耗氧率呈逐渐升高的趋势,其中A组、C组和D组体重的斑石鲷在20 ℃时的耗氧率与其他温度有极显著差异(P<0.01),25与30 ℃之间差异不显著(P>0.05),B组体重的斑石鲷在3个温度条件下的耗氧率存在极显著差异(P<0.01)。经单因素方差分析结果显示,温度对斑石鲷耗氧率的影响差异极显著(P<0.01) 。

图3 温度对斑石鲷耗氧率的影响
同样,随着温度的升高,4个体重组的斑石鲷排氨率也是逐渐增大(见图4)。温度对各体重组斑石鲷排氨率的影响差异极显著(P<0.01)。其中D组斑石鲷中,20与25 ℃的差异不显著(P>0.05),与30 ℃之间差异极显著(P<0.01)。其他体重组各温度之间均有极显著差异(P<0.01)。
2.1.2 体重对斑石鲷耗氧率、排氨率的影响 体重对斑石鲷耗氧率的影响如图5所示,3个温度条件下随着斑石鲷体重的增加,耗氧率均呈现逐渐减小的趋势。经单因素方差分析得出,20 ℃时体重对斑石鲷耗氧率的影响差异不显著(P>0.05),25和30 ℃时体重对斑石鲷耗氧率的影响存在极显著差异(P<0.01)。其中25 ℃时A组与B、C、D组之间存在极显著差异(P<0.01),B、C、D组之间差异不显著(P>0.05),30 ℃时A组与B、C、D组之间存在极显著差异(P<0.01),B组与C组之间存在显著差异(P<0.05)。体重与耗氧率之间的回归关系可用幂函数RO=aWb表示,3个温度下,体重与耗氧率的回归关系方程分别为:y= 2.310 6x-0.19(R2= 0.958 7);y= 2.310 6x-0.19(R2= 0.958 7);y= 3.481 8x-0.228(R2= 0.989 3)。
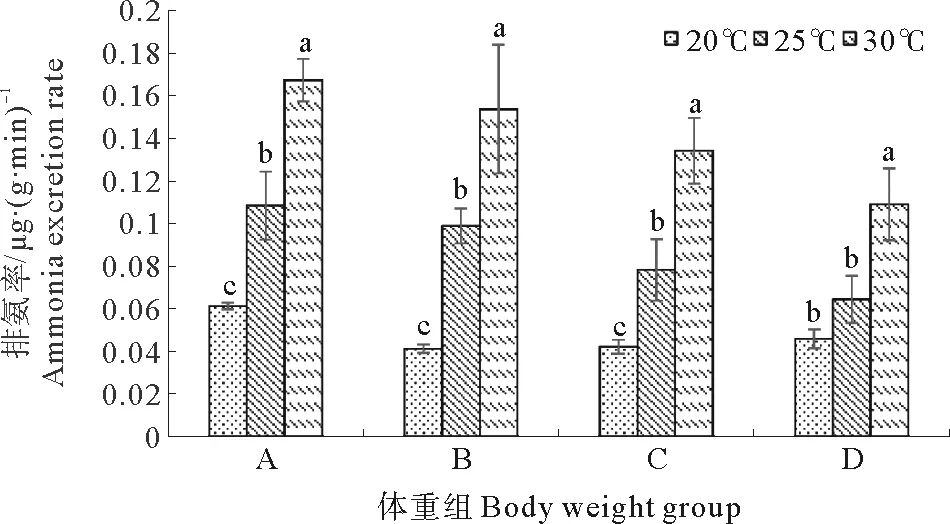
图4 温度对斑石鲷排氨率的影响

图5 体重对斑石鲷耗氧率的影响
体重对斑石鲷排氨率的影响如图6所示,排氨率随鱼体重的增加而降低,其中20 ℃变化趋势不明显。经单因素方差分析得出,体重对斑石鲷排氨率的影响存在极显著差异(P<0.01)。20 ℃时A组与B、C、D组之间存在极显著差异(P<0.01),B、C、D组之间差异不显著(P>0.05),25 ℃时A组与C组间存在显著差异(P<0.05),A组与D组存在极显著差异(P<0.01),B组与D组之间存在显著差异(P<0.05)。30 ℃时A组与D组之间存在极显著差异(P<0.01)。3个温度下,体重与排氨率的相关关系可用如下多项式来表示,分别为:y= 0.006x2- 0.034 4x+ 0.088 7(R2= 0.936 1);y=-0.001 1x2-0.01x+0.120 3(R2= 0.987);y=-0.002 9x2- 0.005 1x+ 0.175 4(R2= 0.976)。

图6 体重对斑石鲷排氨率的影响
2.2 温度和体重对斑石鲷Q10值和O∶N的影响
不同温度和体重下斑石鲷呼吸和排泄的Q10值和O∶N如表2和3所示。
由表2可知,体重50.94~194.56 g的斑石鲷在20~30 ℃时,呼吸Q10的范围是1.126~1.869,平均值为1.494,排泄Q10的范围是1.975~5.787,平均值为3.123。4个体重组的斑石鲷在20~30 ℃范围内随着温度的增加,呼吸Q10和排泄Q10逐渐减小,不同体重间的呼吸Q10变化较小,排泄Q10呈现先增大后减小的趋势。

表2 不同温度和体重下斑石鲷呼吸和排泄的Q10值
Note: ①Body weight group;②Temperature;③Respiration;④Excretion
由表3可知,体重50.94~194.56 g的斑石鲷在20~30 ℃时,O∶N的范围是18.948~47.647,平均值为31.578,随着温度的升高,O∶N逐渐降低,温度对O∶N的影响极显著(P<0.01),体重对O∶N的影响并不显著(P=0.98)。
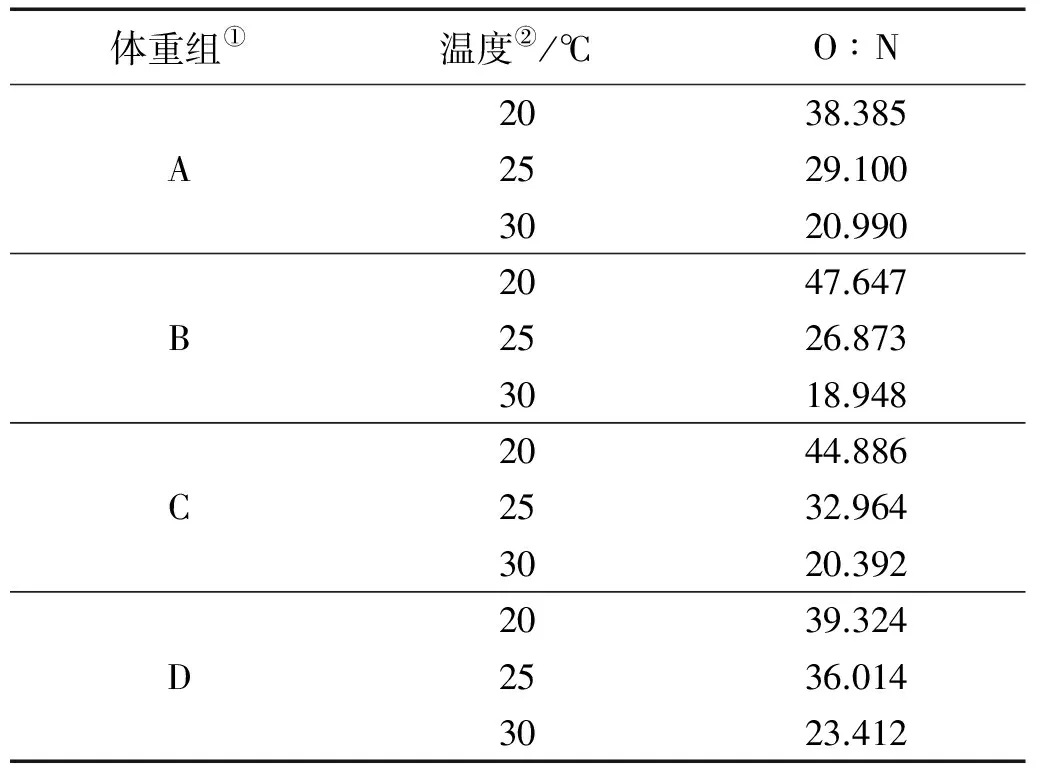
表3 不同温度和体重下斑石鲷的氧氮比(O∶N)
Note: ①Body weight group;②Temperature
2.3 斑石鲷耗氧率和排氨率的昼夜节律变化
2.3.1 斑石鲷耗氧率的昼夜节律变化 停食1 d后,斑石鲷在20~30 ℃时耗氧率的昼夜节律变化如图7所示,最高耗氧率出现在30 ℃时的8:00,为(3.917±0.596) μg·(g·min)-1,最低耗氧率出现在20 ℃的凌晨4:00,为(1.817±0.286) μg·(g·min)-1。20 ℃时耗氧率变化幅度为0.299 μg·(g·min)-1,25 ℃时变化幅度为0.951 μg·(g·min)-1,30 ℃时变化幅度为1.269 μg·(g·min)-1。8:00至18:00划分为白天,20:00至次日8:00划分为夜间,白天平均耗氧率为(2.535±0.204) μg·(g·min)-1,夜间平均耗氧率为(2.531±0.064) μg·(g·min)-1,其耗氧率的昼夜差异不明显。

图7 斑石鲷耗氧率的昼夜节律变化
2.3.2 斑石鲷排氨率的昼夜节律变化 停食1 d后斑石鲷在20~30 ℃时排氨率的昼夜节律变化如图8所示,最高排氨率出现在30 ℃时的8:00,为(0.179±0.049) μg·(g·min)-1,最低排氨率出现在20 ℃的次日8:00,为(0.027±0.009) μg·(g·min)-1。20 ℃时排氨率变化幅度为0.032 μg·(g·min)-1,25 ℃时变化幅度为0.032 μg·(g·min)-1,30 ℃时变化幅度为0.078 μg·(g·min)-1。白天平均排氨率为(0.104±0.007) μg·(g·min)-1,夜间平均排氨率为(0.083±0.007) μg·(g·min)-1,白天平均排氨率明显大于夜间。

图8 斑石鲷排氨率的昼夜节律变化
2.3.3 投喂饲料后斑石鲷耗氧率的变化 投喂饲料之后斑石鲷在20~30 ℃时耗氧率的变化如图9所示,3个温度条件下斑石鲷耗氧率均呈现逐渐下降的趋势,最高点出现在投喂饲料后1~3 h,最低点出现在投喂饲料后19~23 h,投喂饲料后1~12 h的平均耗氧率为(4.099±0.348) μg·(g·min)-1,13~24 h的平均耗氧率为(3.085±0.272) μg·(g·min)-1,显著低于1~12 h时的平均耗氧率。20 ℃时斑石鲷耗氧率的变化幅度为1.669 μg·(g·min)-1,25 ℃时变化幅度为1.942 μg·(g·min)-1,30 ℃时变化幅度为1.637 μg·(g·min)-1,3个温度下耗氧率的变化幅度明显高于停食1 d后的昼夜变化。

图9 投喂饲料后斑石鲷耗氧率的变化
2.3.4 投喂饲料后斑石鲷排氨率的昼夜变化 投喂饲料后斑石鲷在20~30 ℃时排氨率的变化如图10所示,3个温度条件下斑石鲷排氨率均呈现先增大后减小的趋势,20 ℃时排氨率峰值出现在投喂饲料后7 h时,为(0.314±0.060) μg·(g·min)-1,最低点出现在23 h时,为(0.046±0.014) μg·(g·min)-1,变化幅度达到0.269 μg·(g·min)-1,25 ℃时峰值出现在3 h时,为(0.387±0.035) μg·(g·min)-1,最低点出现在23 h时,为(0.126±0.046) μg·(g·min)-1,变化幅度达到0.261 μg·(g·min)-1,30 ℃峰值出现在3 h时,为(0.484±0.094) μg·(g·min)-1,最低点出现在15 h时,为(0.108±0.017) μg·(g·min)-1,变化幅度达到0.376 μg·(g·min)-1,3个温度条件下投喂饲料后排氨率的昼夜变化范围明显高于停食1 d后排氨率的昼夜变化范围。投喂饲料后,随温度增加斑石鲷排氨率达到峰值的时间提前。

图10 投喂饲料后斑石鲷排氨率的变化
3 讨论
3.1 温度对斑石鲷耗氧率和排氨率的影响
水温是影响鱼类生长发育和新陈代谢的重要环境因子,鱼类的体温随水温而变动,鱼体内的代谢是一系列的酶促反应,水温的变化会影响酶的活性,因此代谢率会随之而变化[7]。已有的研究结果表明,鱼类的耗氧率和排氨率与水温有密切的关系,沈勤[8]、张兆琪[9]、刘鹏[10]、刘伟成[11]、戴庆年[12]等,通过对花鲈(Lateolabraxjaponicus)、牙鲆(Paralichthysolivaceus)、大菱鲆(Scophthalmusmaximus)、真鲷(Pagrasamusmajor)、石斑鱼(Epinephelusawoara)的研究,认为鱼类在适宜温度范围内,耗氧率和排氨率随温度的升高而升高。本研究结果表明,在实验温度条件下,斑石鲷耗氧率和排氨率随着温度的升高而升高,与以上研究结果相似,符合鱼类耗氧率和排氨率随水温的上升而增加的一般规律。宋苏祥[13]认为,这是因为随着水温的升高,维持生命的脑、心、肝等重要组织器官的活性增强,各种酶活性提高,鱼类活动强度增大,基础代谢旺盛,表现出的耗氧和氨氮代谢升高的现象。徐奇友[14]认为,温度是影响鱼类生长及生理生化活动最重要的环境因素之一,不同温度条件下鱼类对蛋白质的需求量不同,饲料蛋白质水平显著影响鱼类代谢、蛋白质合成等。本实验过程中,从20 ℃升温到25 ℃时,4个体重组的斑石鲷均出现摄食迅速,活动频繁的现象,从25 ℃升温到30 ℃时,其中C组和D组出现呼吸急促的现象,可能是因为30 ℃超过了斑石鲷适宜温度范围。
温度系数(Q10)也可以反映温度与鱼类代谢速率之间的关系,是温度变化对鱼类代谢的影响指标之一。在适温范围内,水温每升高10 ℃,一般鱼类的耗氧率增加2.3倍,即平均Q10值为2.3[15]。通常认为鱼类的Q10越大,说明其生理活动受温度变化的影响程度越大[16-17]。王波等[18]研究了牙鲆在10.8~21.5 ℃时耗氧率Q10为2.336。James和Robert[19]研究了大眼梭鲈(Stizostedionvvitreum)在20~25 ℃时的排氨率为2.57。闫茂仓[3]等报道了条石鲷在20~25 ℃时呼吸和排泄的Q10分别为1.335和1.345。本研究中斑石鲷呼吸Q10平均是1.494,排泄Q10平均为3.123,均在正常范围内。由表2可知,20~25 ℃时呼吸Q10和排泄Q10均大于25~30 ℃时,表明25~30 ℃条件下,斑石鲷生理活动受温度变化的影响程度小于20~25 ℃时,印证了斑石鲷最适生长温度在25~30 ℃范围内,且30 ℃超出斑石鲷适宜温度范围。
3.2 体重对斑石鲷耗氧率和排氨率的影响
体重是影响水生动物耗氧率和排氨率的重要因素之一。本研究中,体重对斑石鲷的耗氧率和排氨率有显著的影响,体重小的斑石鲷耗氧率和排氨率比体重大的要高,这与其他鱼类研究结果类似[3,5,9-10,13]。姜祖辉[20]等认为这可能与水生动物在生长过程中的组织、脏器的比重有关,直接维持生命的组织和脏器如肾脏、肝脏等的新陈代谢高于非直接维持生命的其他组织如肌肉、脂肪等。在动物生长过程中,这2种组织的比率随之减小,即肌肉和脂肪等积累增多,从而引起随个体增大而单位重量的耗氧率和排氨率降低的现象。
3.3 供能物质分析
鱼类体内的主要能源物质为蛋白质和脂肪,而O∶N的大小表示蛋白质与脂肪和碳水化合物提供给生物能量的比率,利用O∶N能分析其代谢过程中能源利用的化学本质[21]。国内外对此做过许多研究,当完全以蛋白质为氧化基质提供能量时,其O∶N为7~10[22];当蛋白质和脂肪共同提供能量时,其O∶N为24[23];随着脂肪和碳水化合物被利用的比例升高,其O∶N也随之增大,当完全以脂肪或碳水化合物为能源物质时,其O∶N将无限大[24]。陈宣雄等[25]研究指出了条石鲷 (Oplegnathusfasciatus)也遵循这一规律,在10~35 ℃范围内条石鲷幼鱼平均O∶N=72.083,表明10~35 ℃时条石鲷幼鱼主要以脂肪和碳水化合物为能源,蛋白质次之。本研究中,体重50.94~194.56 g的斑石鲷在20~30 ℃时平均O∶N分别为42.56、31.24和20.94,表明在20~30 ℃范围内,斑石鲷由脂肪和碳水化合物为主要能源逐渐转变为蛋白质和脂肪共同提供能量,随着温度的升高,O∶N逐渐降低,而体重对O∶N的影响并不显著。
3.4 斑石鲷耗氧率和排氨率的昼夜节律变化
一般将鱼类代谢水平的昼夜差异分为三种类型:(1)白天大于夜间,如平鲷(Rhabdosargussarba)[26];(2)夜间大于白天,如青石斑鱼[12];(3)昼夜差异不明显,如黑鲷(Sparusmacrocephlus)[27]。昼夜节律性是生物体内生物钟作用的结果,是通过长期的进化,生物对这种周期性的环境变化所产生的适应,即所谓生物周期[10]。
本研究中,斑石鲷耗氧率昼夜差异不明显,属于第三种类型,可能是因为实验过程中水温稳定且位于室内,不受光线的影响,因此斑石鲷的活动周期没有昼夜节律的变化,因此耗氧率昼夜差异不明显。Clausen[28]认为鱼类耗氧率有规律的昼夜变化,耗氧率高时表示鱼类进食或进行其他活动,由图7可知斑石鲷耗氧率较高的时刻为8:00、20:00和4:00,这些时刻可能是斑石鲷在自然环境中的摄食时间。排氨率白天大于夜间,属于第一种类型,可能是昼夜间环境因子的周期变化导致斑石鲷生理状态发生相应的变化,也可能与潮汐等气象条件的周期性响应有关,具体原因还需进一步的研究。
投喂饲料后斑石鲷在3个温度条件下的耗氧率呈现逐渐减小的趋势,排氨率呈现先增大后减小的趋势。投喂饲料后斑石鲷摄食运动剧烈,呼吸系统迅速响应,造成耗氧率迅速达到最大值。排氨率经过2~6 h达到峰值,且温度较高时排氨率达到峰值的时间提前,说明在20~30 ℃范围内,随温度的升高,各种消化酶的活性提高,组织代谢加快,其排氨率更快达到最大值。
3.5 斑石鲷代谢对水质的影响
鱼类的耗氧率因种而异,这与各种鱼类的生态习性不同有关[29]。一般而言,栖息于水域中上层的游泳快速的鱼类耗氧率较高,而栖息于底层,活动程度较少的鱼类如鰕虎鱼类等则较低[9];同时,鱼类的耗氧率还跟食性相关,通常肉食性鱼类的耗氧率较高,温和食性鱼类则较低[30]。斑石鲷系温热带近海沿岸中下层鱼类,并且属于肉食性鱼类[1],其耗氧率大于大多数底层鱼类[9,12]和温和食性鱼类[7],但略低于某些肉食性鱼类[30],表明养殖过程中应该同其他中上层肉食性鱼类一样,养殖水体中应保持较高的溶解氧。

4 结语
在20~30 ℃条件下,斑石鲷的耗氧率和排氨率随温度升高而升高,其生理活动受温度变化的影响程度逐渐变小,耗氧率变化范围为(0.179±0.203)~(3.513±0.165) μg·(g·min)-1,排氨率变化范围为(0.041±0.002)~(0.167±0.010) μg·(g·min)-1,斑石鲷最适生长温度在25~30 ℃范围内,且30 ℃超出其鲷适宜温度。在20~30 ℃时,温度升高,斑石鲷由脂肪和碳水化合物为主要能源逐渐转变为蛋白质和脂肪共同提供能量,随着温度的升高,O∶N逐渐降低。
斑石鲷耗的氧率和排氨率随体重的增加而降低,但体重对O∶N影响并不显著,表明在实验鱼体重范围内,斑石鲷无食性转化。
本研究中,斑石鲷耗氧率昼夜差异不明显,排氨率的昼夜差异白天大于夜间。投喂饲料后斑石鲷耗氧率最高点出现在投喂饲料后1~3 h,最低点出现在投喂饲料后19~23 h,呈逐渐降低的趋势,排氨率呈现先升高后降低的趋势,温度升高导致斑石鲷排氨率达到峰值的时间提前。
[1] 王雨福. 斑石鲷(Oplegnathuspunctatus)早期生长特征及消化系统发育的研究[D]. 青岛: 中国科学院研究生院(海洋研究所), 2015. Wang Y F. Studies on Early Growth and Development of Digestive System in Spotted Knifejaw (Oplegnathuspunctatus)[D]. Qingdao: Institute of Oceanology, Chinese Academy of Science, 2015.
[2] 李加儿, 曹守花, 区又君, 等. 温度、盐度和pH对鲻幼鱼耗氧率、排氨率以及窒息点的影响[J]. 中国水产科学, 2014, 21(5): 954-962. Li J E, Cao S H, Qu Y J, et al. Influence of temperature, salinity, and pH on oxygen consumption rate, ammonia excretion rate, and suffocation point injuvenileMugilcephalus[J]. Journal of Fishery Sciences of China, 2014, 21(5): 954-962.
[3] 闫茂仓, 单乐州, 谢起浪, 等. 温度、盐度及体重对条石鲷幼鱼耗氧率和排氨率的影响[J]. 海洋科学进展, 2008, 26(4): 486-496. Yan M C, Shan L Z, Xie Q L, et al. Influence of temperature, salinity and body weight on oxygen consumption and ammonia excretion ofOplegnathusfasciatusjuvenile[J]. Advances in Marine Science, 2008, 26(4): 486-496.
[4] 侯晓飞. 半现场条件下温度、投喂水平和短期盐度胁迫对许氏平鲉(Sebastesschlegeli)氨氮、磷排泄影响的研究[D]. 青岛: 中国海洋大学, 2008. Hou X F. Research on Effects of Temperature, Feeding Level and Salinity on Rates of Ammonia Excretion and Phosphate Excretion ofSebastesschlegeliUnder Farm-like Conditions[D]. Qingdao: Ocean University of China, 2008.
[5] 王刚, 李加儿, 区又君, 等. 环境因子对卵形鲳鲹幼鱼耗氧率和排氨率的影响[J]. 动物学杂志, 2011, 46(6): 80-87. Wang G, Li J E, Qu Y J, et al. Influence of environmental factors on oxygen consumption and ammonia excretion of juvenileTrachinotusovatus[J]. Chinese Journal of Zoology, 2011, 46(6): 80-87.
[6] 王刚, 李加儿, 区又君, 等. 卵形鲳鲹幼鱼耗氧率和排氨率的初步研究[J]. 动物学杂志, 2010, 45(3): 116-121. Wang G, Li J E, Qu Y J, et al. Oxygen consumption and ammonia excretion of the juvenileTrachinomsovatus[J]. Chinese Journal of Zoology, 2010, 45(3): 116-121.
[7] 周洪琪, 潘兆龙, 李世钦, 等. 草鱼代谢能的研究[J]. 水产学报, 1998, 22(1): 29-33. Zhou H Q, Pan Z L, Li S Q, et al. Metabolic energy ofCtenopharyngodonidella[J]. Journal of Fisheries of China, 1998, 22(1): 29-33.
[8] 沈勤, 徐善良, 严小军, 等. 温度对花鲈饥饿代谢的影响[J]. 中国水产科学, 2008, 15(3): 500-505. Shen Q, Xu S L, Yan X J, et al. Effects of water temperature on metabolizability of starvedLateolabraxjaponicus[J]. Journal of Fishery Sciences of China, 2008, 15(3): 500-505.
[9] 张兆琪, 张美昭, 李吉清, 等. 牙鲆鱼耗氧率、氮排泄率与体重及温度的关系[J]. 青岛海洋大学学报, 1997, 27(4): 49-55. Zhang Z Q, Zhang M Z, Li J Q, et al. Oxygen consumption and nitrogen excretion ofParalichthysolivaceuswith different body weights at different water temperature[J]. Jouranl of Ocean University of Qingdao, 1997, 27(4): 49-55.
[10] 刘鹏, 宋协法, 葛长字. 温度、体重和昼夜节律对大菱鲆幼鱼代谢的影响[J]. 河北渔业, 2009, 5: 1-4, 27. Liu p, Song X F, Ge C Y. Effect of temperature, body weight and diurnal rhythm on metabolism ofScophthalmusmaximus[J]. Hebei Fisheries, 2009, 5: 1-4, 27.
[11] 刘伟成, 冀德伟, 张赛乐, 等. 温度、盐度和体重对美国红鱼幼鱼呼吸和排泄的影响[J]. 浙江农业学报, 2014, 26(6): 1478-1483. Liu W C, Ji D W, Zhang S L, et al. Effects of different temperature, salinity and body weight on respiration and excretion of juvenileSciaenopsocellatus[J]. Acta Agriculturae Zhejiangensis, 2014, 26(6): 1478-1483.
[12] 戴庆年, 赵莉莉. 青石斑鱼耗氧率研究[J]. 水产科学, 1994, 13(3): 6-9. Dai Q N, Zhao L L. Study on the oxygen consumption rate ofEpinephelusawoara[J]. Fisheries Science, 1994, 13(3): 6-9.
[13] 宋苏祥, 刘洪柏, 孙大江, 等. 史氏鲟稚鱼的耗氧率和窒息点[J]. 中国水产科学, 1997, 4(5): 101-104. Song S X, Liu H B, Sun D J, et al. The asphyxiation point and oxygen consumption rate ofAcipenserschrencki[J]. Journal of Fishery Sciences of China, 1997, 4(5): 101-104.
[14] 徐奇友, 黄金凤. 环境温度和蛋白质水平对鱼类生长和代谢的影响[J]. 东北农业大学学报, 2011, 42(12): 1-8. Xu Q Y, Huang J F. Effect of environmental temperature and protein level on growth and metabolism of fish[J]. Journal of Northeast Agricultural University, 2011, 42(12): 1-8.
[15] 童裳亮. 鱼类生理学[M]. 北京: 北京科学出版社,1988: 242-243. Tong S L. Fish Physiology[M]. Beijing: Science Press of Beijing,1988: 242-243.
[16] 徐钢春, 顾若波, 闻海波. 温度对花(鱼骨)耗氧率和排氨率的影响[J]. 湖泊科学, 2006, 18(4): 431-436. Chen G C, Gu R B, Wen H B. Effect of temperature on oxygen consumption rate and ammonia excretion rate ofHemibarbusmaculatusBleeker[J]. Journal of Lake Sciences, 2006, 18(4): 431-436.
[17] 廖志洪, 林小涛, 王春, 等. 黄颡鱼仔、稚、幼鱼耗氧率及氨氮排泄率的初步研究[J]. 生态科学, 2004, 23(3): 223-226. Liao Z H, Lin X T, Wang C, et al. Primary study on oxygen consumption and NH3-N excretion rate of larvae, juvenile and youngPelteobogrusfulvidraco[J]. Ecologic Science, 2004, 23(3): 223-226.
[18] 王波, 李继强, 曹志海, 等. 犬齿牙鲆幼鱼呼吸耗氧率的研究[J]. 海洋科学进展, 2003, 21(3): 325-330. Wang B, Li J Q, Cao Z H, et al. Study on respiration and oxygen consumption rate of summer flounder (Paralichthysdentatus) juvenile[J]. Advances in Marine Science, 2003, 21(3): 325-330.
[19] Jamsaf A F, Robert C S. Effects of temperature on dielammonia excretion of fingerling walleye[J]. Aquaculture, 1992, 102: 115-126.
[20] 姜祖辉, 王俊, 唐启升. 菲律宾蛤仔生理生态学研究Ⅰ. 温度、体重及摄食状态对耗氧率及排氨率的影响[J]. 海洋水产研究, 1999, 21(1): 40-44. Jiang Z H, Wang J, Tang Q S. Studies on physiological ecology of short-necked clam (Ruditapesphilippinarum) I.effects of temperature, body weight and feeding state on oxygen consumption rate and ammonia excretion rate[J]. Marine Fisheries Research, 1999, 20(1): 40-44.
[21] Harris E. The nitrogen cycle in long island sound[J]. Bull Bingharm Oceanogr Collect, 1959, 17: 31-65.
[22] Myxand P. Respiration and nitrogen excretion of zooplankton IV: The influence of starvation on the metabolism and biochemical composition of som species[J]. Mar Biol, 1976, 37: 47-58.
[23] Ikeda T. Nutritional ecology of marine zooplankton[J]. Mem Fac Fish Hokkaido Univ, 1974, 22: 1-97.
[24] Conover R I, Cover E D S. Respiration and nitrogen excretion by some marine zooplankton in relation to their life cycels[J]. J Mar BiolAssoc U K, 1968, 48: 49-75.
[25] 陈宣雄, 徐善良, 沈庞幼, 等. 温度对条石鲷幼鱼代谢率、排泄率及窒息点的影响[J]. 海洋学研究, 2012, 30(1): 95-101. Cheng X X, Xu S L, Sheng P Y, et al. Impacts of temperature on metabolic, excretion rate and asphyxiation point of juvenileOplegnathusfasciatus[J]. Journal of Marine Sciences, 2012, 30(1): 95-101.
[26] 丁彦文, 李加儿. 平鲷Rhabdosargussarba(Forsk I)幼鱼耗氧率的初步研究[J]. 湛江海洋大学学报, 2000, 20(3): 8-12. Ding Y W, Li J E. A preliminary study on the oxygen consumption of fry of flat breamRhabdosargussarba(Forsk I)[J]. Journal of Zhanjiang Ocean University, 2000, 20(3): 8-12.
[27] 朱爱意, 谢佳彦, 章韶兵. 黑鲷耗氧率昼夜变化及与体重、水温的关系研究[J]. 南方水产, 2007, 3(6): 63-66. Zhu A Y, Xie J Y, Zhang S B, Diurnal rhythm of oxygen consumption and effects of body weight and water temperature on oxygen consumption rate ofSparusmacrocephalus[J]. South China Fisheries Science, 2007, 3(6): 63-66.
[28] Clause N R G. Oxygen consumption in fresh water fishes[J]. Ecol, 1936, 17(2): 216-226.
[29] 川本信之.魚類生理生態学[M].//水産学全集13.東京都: 恒星社厚生閣刊, 1960: 9-24.
[30] 王跃斌, 孙忠, 余方平, 等. 温度对黑鳍棘鲷耗氧率与排氨率的影响[J]. 海洋渔业, 2007, 29(4): 375-379. Wang Y B, Sun Z, Yu F P, et al. Effects of temperature on oxygen consumption rate andammonia excretion rate ofAcanthopagrusschlegeli[J]. Marine Fisheries, 2007, 29(4): 375-379.
[31] Brafield A E. Laboratory Studies of Energy Budgets. In Fish Energetic: New Perspectives[M]. London: Croom Helm, 1985: 257-282.
[32] Jobling M. Fish Bioenergetics[M]. Great Britain: Edmunds Bury Press, 1994.
[33] 柳瑶. 生物流化床养殖污水处理系统的设计与实验研究[D]. 青岛: 中国海洋大学, 2013. Liu Y. Design and Experimental Study of Cyclo-bio Fluidized Sand Bed in Recirculating Aquaculture Systems[D]. Qingdao: Ocean University of China, 2013.
Abstract: Through closed water breathing experiment, we evaluated the oxygen consumption rate and ammonium excretion rate ofOplegnathuspunctatusand their circadian rhythms under the effects of water temperature (20、25 and 30 ℃) and different body weights( (50.94±5.50), (103.61±10.48), (154.58±7.56) and (194.56±14.56)g). The results indicated that the oxygen consumption and ammonium excretion rates ofO.punctatusincreased with temperature and weight but were inversely proportional to weight, The data showed that daily average oxygen consumption rate during the day was (2.535±0.204) μg·(g·min)-1and the average oxygen consumption rate in the night was (2.531±0.064) μg·(g·min)-1. Therefore, the oxygen consumption rate of day and night is not different significantly. Moreover, we found that the average of ammonium excretion rate in the day was (0.104±0.007) μg·(g·min)-1and the averageof ammonia excretion rate in the night was (0.083±0.007) μg·(g·min)-1, and the average of ammonium excretion rate in the daytime was obviously higher. Our further studies found a trend of gradual reduction of the oxygen consumption rate ofO.punctatusafter feeding with the highest appeared between 1 and 3 h and the lowest between 19 and 23 h, and ammonium excretion rate increased first and then decreased. In addition, we also found that the highest rate of ammonia excretion time advanced when the temperature increased.
Key words:Oplegnathuspunctatus; temperature; weight; oxygen consumption rate; ammonium excretion rate; circadian rhythm
责任编辑 朱宝象
Influences of Temperature and Weight on Oxygen Consumption Rate and Ammonium Excretion ofOplegnathuspunctatusand Their Circadian Rhythms
SONG Xie-Fa1, CHENG Ya-Wei1, XING Dao-Chao1, PENG Lei1, ZHAI Jie-Ming2, LI Bo2, DONG Deng-Pan1
(1. College of Fisheries, Ocean University of China, Qingdao 266003, China;2. Laizhou Mingbo Aquatic Co. Ltd, Yantai 261418, China)
青岛市海洋经济创新发展区域示范项目资助 Supported by the Regional Demonstration Project of Qingdao Marine Economy Innovation and Development
2016-11-10;
2017-02-08
宋协法(1964-),男,博士,教授,主要从事设施渔业和养殖污水处理研究工作。E-mail: yuchuan@ouc.edu.cn
S917.4;S965.399
A
1672-5174(2017)09-021-09
10.16441/j.cnki.hdxb. 20160380
宋协法, 程亚伟, 邢道超, 等. 温度、体重对斑石鲷耗氧率、排氨率的影响及昼夜节律变化[J]. 中国海洋大学学报(自然科学版), 2017, 47(9): 21-29.
SONG Xie-Fa, CHENG Ya-Wei, XING Dao-Chao,et al. Influences of temperature and weight on oxygen consumption rate and ammonium excretion ofOplegnathuspunctatusand their circadian rhythms [J].Periodical of Ocean University of China, 2017, 47(9): 21-29.
猜你喜欢
小哥白尼(野生动物)(2021年9期)2022-01-17
好孩子画报(2020年10期)2020-11-02
故事作文·低年级(2020年10期)2020-10-21
儿童时代·幸福宝宝(2020年9期)2020-09-08
学生天地(2020年14期)2020-08-25
数学小灵通(1-2年级)(2020年6期)2020-06-24
特别文摘(2018年3期)2018-08-08
探索科学(2017年5期)2017-06-21
诗选刊(2015年6期)2015-10-26
中国火炬(2012年2期)2012-07-24
