
拟南芥 WOX10 基因启动子的克隆与序列分析
2017-07-03 13:21王占军黄士平欧祖兰徐忠东韩久盼余文敬
黄山学院学报 2017年3期
王占军,黄士平,欧祖兰,徐忠东,韩久盼,余文敬
(合肥师范学院 生命科学学院,安徽 合肥,230601)
拟南芥 WOX10 基因启动子的克隆与序列分析
王占军,黄士平,欧祖兰,徐忠东,韩久盼,余文敬
(合肥师范学院 生命科学学院,安徽 合肥,230601)
为探索功能未知的拟南芥 WOX10 基因的表达规律,开展对拟南芥 WOX10 基因启动子的分子克隆及测序分析,运用 PlantCARE 在线工具预测测序结果中可能含有的顺式作用元件。结果表明,除 TATA-box 和 CAAT-box 外, 拟南芥 WOX10 基因启动子含有生长素、 赤霉素和脱落酸等植物激素以及光信号,干旱、盐分和压力等胁迫相关的顺式作用元件。该研究结果有益于解析拟南芥 WOX10 基因的转录调控规律及其功能。
WOX10 基因;拟南芥;启动子;顺式作用元件
WUSCHEL-RELATED HOMEOBOX (WOX)是仅存在于植物细胞内的一类编码相关转录因子的基因 家 族[1]。 1996 年 ,Laux 等[2]利 用 EMS 诱 变 的 方 法 ,在拟南芥中克隆并鉴定出 WUS基因; 该基因编码291 个氨基酸, 具有维持植物茎端分生组织中干细胞数量的功能。WUS 的同源异型结构域(Homeodomain 区) 是由 66 个氨基酸残基所构成的特殊空间结构(Helix 1-loop-helix 2-turn-helix 3),该 结构及 其 特 殊 功 能 引 起 了 研 究 人 员 的 广 泛 关 注[3]。Haecker 等[4]通 过 对 拟 南 芥 基 因 组 的 开 放 阅 读 框(ORF)进行同源性比对分析,共鉴定出 14 个基因的ORF 区域与 WUS 基因的 Homeodomain 区存在高度的一致性或相似性, 这些基因被命名为 WOX1-WOX14。 目前,WOX 基因家族已在拟南芥、水稻、葡萄 、高 粱 、玉 米 等 30 多 种 植 物 中 鉴 定 出 来[5]; 根 据 功能分析的结果,可将 15 个基因家族成员分为调节植物 干细 胞活动[6]、控 制 细 胞 胚 胎发生[7]、调 节 侧 生 器 官的 生 长 和 发 育[8]以 及 功 能 未 知 WOX10 基 因 共 4 种类型。
启动子是基因表达调控系统中重要的顺式作用元件,控制着基因表达的起始时间与表达程度,是基因表 达 活性的 “开关 ”[9]。 启 动子 一 般是位 于基因编 码区 5'端上游的一段特殊的核苷酸序列,它包含 RNA聚合酶以及一个或多个特定转录因子的结合位点,结 构 上 通 常 包 括 核 心 启 动 子 元 件 (TATA-box、CAAT-box)和应答调控元件,参与转录的起始与调控[10]。
鉴于拟南芥 WOX10 基因的功能目前尚未见报道, 本研究针对 WOX10 基因的启动子进行克隆及分析,获得该启动子区域基因表达调控网络的一系列调控元件;此外,旨在基于 WOX10 基因启动子序列的分析结果,为进一步研究 WOX10 基因的表达规律和基因功能提供理论依据。
1 实验部分
1.1 启动子序列的获取
根 据 NCBI (National Center for BiotechnologyInformation,美国国立生物 技术信息中心)公布 的 拟南芥全基因组测序结果,利用 Map Viewer 确定拟南芥 WOX10 (简写为:AtWOX10) 基因序列的位置信息,下 载该 基因 翻 译起始位 点 (ATG)上 游 2000 bp的启动子序列(简写为:pAtWOX10)。
1.2 实验试剂及方法
1.引物设计:依据 pAtWOX10 核苷酸序列,使用Primer Premier v5.0 软件设计启动子克隆引物,具体信息如下:pAtWOX10–F:AATAAACTAAAGATCCCTTTATTCATAAACAAAAG (从 5' 至 3' 端 ),pAt-WOX10-R:CTCCTCTTTCACCACTTTGCCTTCTC(从5'至 3'端)。
2. 利用 BioTeke 试剂盒提取拟南芥幼苗的基因组 DNA, 利用琼脂糖凝胶电泳法检测提取 DNA 的质量。
3. 使用 KOD 高保真酶进行 pAtWOX10 基因启动子序列的 PCR 扩增,反应体系的总体积为 50μL,具 体 包 括 :10 ×PCR Buffer for KOD-Plus-Neo,5.0μL;2 mM dNTPs,5.0μL;25mM MgSO4,3.0μL;pAtWOX10 – F 引 物 ,1.5μL;pAtWOX10 – R 引 物 ,1.5μL;KOD-Plus-Neo 酶 1.0 μL; 拟 南 芥 DNA,3.0μL;ddH2O,30.0μL。
4.PCR 反应程序为:94℃预变性 5min; 循环内:94℃变 性 30sec,50℃退 火 30sec,72℃延 伸 2min,执行 30 次循环;72℃延伸 10 min。
5.PCR 产物检测:使用 1%的琼脂糖凝胶中检测PCR 产物的大小及质量。
6.胶回收及连接、转化:针对 PCR 产物大小与预期目标产物相符的条带进行切胶,使用 Axygen 公司的 DNA 凝胶回收试剂盒进行胶回收,将胶回收产物加 Poly (A) 尾 处 理 后 , 连 接 至 TaKaRa 公 司 的pMDTM19-T 载体,转化感受态细胞,菌液 PCR 检测无误后送至上海英俊生物技术有限公司(简写为:英俊公司)测序。
1.3 序列分析方法
运 用 PlantCARE (http://bioinformatics.psb.ugent. be/webtools/plantcare/html/) 在 线 分 析 软 件 对 At-WOX10 基因启动子的测序结果进行分析。
2 结果与讨论
2.1 AtWOX10 基因及其启动子的位置信息
依据 NCBI 数据库中获得的 AtWOX10 基因及其启动子的相关信息,揭示 AtWOX10 基因位于拟南芥的 1 号 染色体上(如图 1A),利 用 GSDS(http://gsds.cbi.pku.edu.cn/)基因结构分析工具解析 AtWOX10基因序列后,发现 AtWOX10 基因全长 783bp,其中编 码 区 长 度 为 594bp, 含 有 3 段 外 显 子 区 域(7184485-7184642;7184731-7185129;7185221-7185267) 和两段内含子区域 (7184643-7184730;7185130-7185220)(见图 1B 和表 1)。
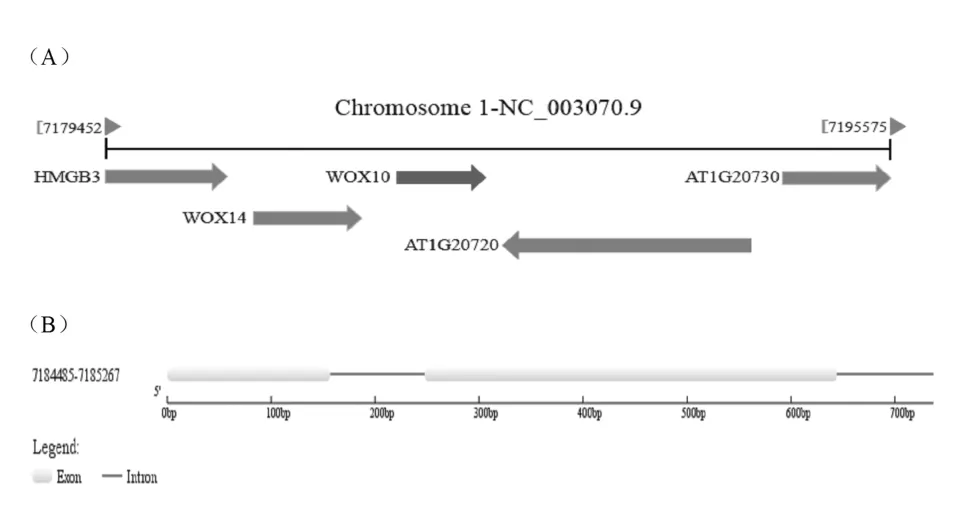
图1 AtWOX10 基因在染色体上位置(A)及基因结构(B)示意图
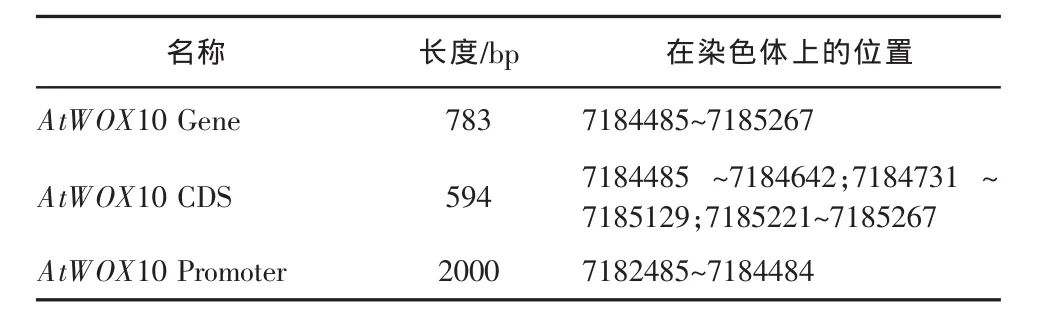
表1 AtWOX10 基因及其启动子信息
2.2 AtWOX10 基因启动子的克隆

图2 AtWOX10 基因启动子序列的PCR 产物(A)和胶回收产物(B)
利用具有高保真性的 KOD 聚合酶和特异性引物克隆 AtWOX10 基因的启动子序列, 由于 PCR 反应体系的退火温度较低,造成产物出现了杂带,针对PCR 产物进行胶回收纯化后, 凝胶电泳检测的条带单一。
2.3 AtWOX10 基因启动子的序列分析
英俊公司测序结果显示, 原长为 2000bp 的 At-WOX10 基因启动子, 实际克隆产物的长度为 1999bp, 在 NCBI 公 布 的 AtWOX10 基 因 启 动 子 -1922 bp 的位置缺失 1 个 A 碱基。 运用 PlantCARE 在线工具分析 AtWOX10 基因启动子的测序结果,详细信息如表 2 所示。 在 ATG 上游 500bp 长度内可能存在 4个 TATA-box(-100;-240;-280;-473)、7 个 CAAT-box(-163;-199;-221;-233;-249;-363;-391),具有真核生物核心启动子元件典型的特征,二者共同介导的 RNA 聚合酶参与的 WOX10 基因转录过程的起始与调控,并且在该序列的多个位点重复出现,这对于提升基因对外界影响因素变化应答速度及反应效率是必不可少的。此外,该序列还存在多种特异性的顺式调控元件,包括光诱导应答反应、激素诱导应答反应、胁迫诱导应答反应以及特异性表达产物调控等多方面功能;其中光应答元件共 7 种,包括 AE-box、E-box、G-box、GT-1、PIF3、SORLIP1AT 和-10 promoter element。 -10 promoter element是对蓝光、白光和紫外光应答的元件,在植物的花器官发育调控中起着重要调控作用。 激素应答调控元件共 8 种, 包括 ARF、ASF1、AtMYB2、DPBF、GARE、MYBATRD22、TGA-element和 W-BOX,其中生长素应答元件 ARF 与赤霉素应答元件 GARE 常在植物的根部转录调控中发挥作用。胁迫诱导应答元件共 9 种,包括 ABRE、ACGT、LTRE、MYB1AT、MYB1LEPR、MYB2、MYBRE、MYC和 SREATMSD,其中 MYB 类水分胁迫与 LTRE 低温胁迫诱导元件是根部生长发育必不可少的调控元件。此外,还发现如调控花器官部位相关基因表达的应答元件 AP1 和 MYBPLANT,根发育调控和木质部形成相关的基因表达的应答元件 RHERPATEXPA7 和 XYLAT,以及细胞周期蛋白表达的调控元件MYBCOREATCYCB1 等其他有关转录起始和特异性表达产物合成控制的应答作用元件,针对这些表达部位明确的应答作用元件结果进行分析,将有利于深入解析 AtWOX10 基因的表达调控规律及其基因功能。

表2 AtWOX10 基因启动子区的顺式作用元件预测结果
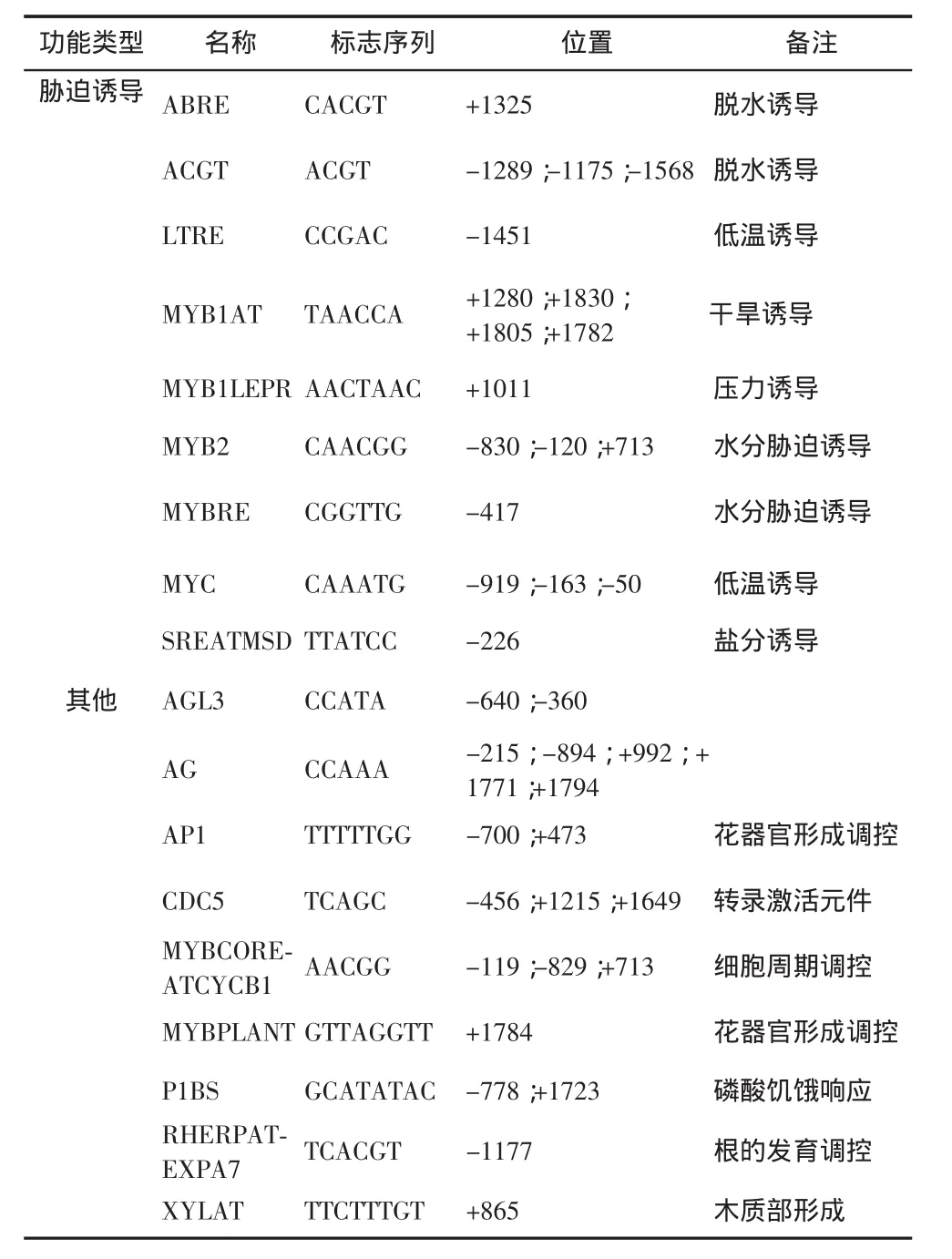
注 :(-)表 示 pAtWOX10 序 列 ,(+)表 示 与 pAtWOX10 互补序列。
3 结论及展望
由上述 AtWOX10 基因启动子中相关顺式作用元件分析结果可知,当外界环境处于干旱、高盐以及压力等逆 境胁 迫时,AtWOX10 基因可能会被激活(MYB1LEPR、MYB1AT、SREATMSD);也可能存在对蓝色、 白色和紫外光等产生相应的应答机制(-10 promoter element);能够被诸如 生长 素 、赤 霉素 和 脱落酸等植物生长激素诱导表达(ARF、TGA-element、GARE、DPBF); 同时,AtWOX10 基因启动子还具有一些特殊的应答作用元件,例如花器官中基因特异性表达的因子 AP1。 综上所述,AtWOX10 基因转录调控可能受到光信号,生长素、赤霉素和脱落酸等植物激素以及胁迫等多种信号途径的协同作用; 但是,At-WOX10 基因具体的转录调控规律有待进一步研究。
本文对 AtWOX10 基因启动子进行了克隆及序列分析,但关于其表达的具体部位,真实存在哪些与其表达相关的光照、激素和温度等因素,有待下一步通过构建植物表达载体,并将其转化至拟南芥中加以验证;同时,本研究也为 AtWOX10 基因的功能解析提供了参考信息。
[1]Lian G,Ding Z,Wang Q,et al.Origins and evolution of WUSCHEL-related homeobox protein family in plant kingdom[J].The Scientific World Journal,2014:534140.
[2]Laux T,Mayer KF,Berger J,et al.The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis[J].Development,1996,122(1):87-96.
[3]Kamiya N,Nagasaki H,Morikami A,et al.Isolation and characterization of a rice WUSCHEL-type homeobox gene that is specifically expressed in the central cells of a quiescent center in the root apical meristem[J].The Plant Journal,2003,35(4):429-441.
[4]HaeckerA,Groβ -HardtR,Geiges B,etal.Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana[J]. Development,2004,131(3):657-668.
[5]Zhang X,Zong J,Liu J,et al.GenomeWide analysisof WOX gene family in Rice,Sorghum,Maize,Arabidopsis and Poplar[J].JournalofIntegrative PlantBiology,2010,52(11):1016-1026.
[6]Mayer KF,Schoof H,Haecker A,et al.Role of WUSCHEL in regulating stem cellfate in the Arabidopsisshoot meristem[J].Cell,1998,95(6):805-815.
[7]Palovaara J,Hakman I.Conifer WOX-related homeodomain transcription factors,developmental consideration and expression dynamic of WOX2 during Picea abies somatic embryogenesis[J].Plant Molecular Biology,2008,66(5):533-549.
[8]Vandenbussche M,Horstman A,Zethof J,et al.Differential recruitment of WOX transcription factors for lateral development and organ fusion in Petunia and Arabidopsis[J]. The Plant Cell,2009,21(8):2269-2283.
[9]聂丽娜,夏兰琴,徐兆师,等.植物基因启动子的克隆及其功能研究进展[J].植物遗传资源学报,2008,9(3):385-391.
[10]王颖,麦 维 军 ,梁承邺,等.高 等 植 物 启 动 子 的 研 究 进 展[J].西北植物学报,2004,23(11):2040-2046.
责任编辑:胡德明
The Cloning and Sequence Analysis of Promoter of WOX Gene Family in Arabidopsis thaliana
Wang Zhanjun,Huang Shiping,Ou Zulan,Xu Zhongdong,Han Jiupan,Yu Wenjing
(School of Life Sciences,Hefei Normal University,Hefei 230601,China)
In order to study the expression rule of WOX10 with unknown function in Arabidopsis thaliana,the promoter of WOX10 gene in Arabidopsis thaliana was cloned and sequenced.Cis-acting elements were predicted by PlantCARE in the promoter of AtWOX10 gene.The results showed that there were TATA-box,CAAT-box and some cis-acting elements in the promoter of AtWOX10 gene,including a variety of plant hormones like auxin,gibberellin and abscisic acid,light responsive element,and also cisacting elements related to drought,salt and stress.The results can also be useful for future studies of AtWOX10 gene in transcriptional regulation and function.
WOX10 gene;Arabidopsis thaliana;promoter;cis-acting element
Q74
:A
:1672-447X(2017)03-0058-04
2016-09-20
安徽省高校自然科学基金重点项目(KJ2015A186);安徽省自然科学基金面上项目 (1708085MC76);合肥师范学院人才科研启动基金项目(2013rcjj01);合肥师范学院产学研项目(2014cxy33);合肥师范学院课程建设项目(2013hxk14)
王占军(1984-),安徽寿县人,合肥师范学院讲师,研究方向为植物细胞分子生物学。
猜你喜欢
疯狂英语·初中天地(2022年2期)2022-07-07
疯狂英语·初中版(2022年2期)2022-05-04
中国特种设备安全(2022年1期)2022-04-26
国际放射医学核医学杂志(2020年4期)2020-07-27
中国核电(2017年2期)2017-08-11
中国自行车(2017年5期)2017-06-24
上海农业学报(2017年3期)2017-04-10
现代工业经济和信息化(2016年2期)2016-05-17
汽车维修与保养(2015年8期)2015-04-17
中共合肥市委党校学报(2014年2期)2014-08-03
