
致病疫霉ATP6基因单倍型与地理因素的相关性分析
2017-06-30 20:16张佳峰吴娥娇罗桂火祝雯詹家绥
江苏农业科学 2017年8期
张佳峰+吴娥娇+罗桂火++祝雯++詹家绥
摘要:致病疫霉(Phytophthora infestans)是马铃薯晚疫病的病原菌,对世界马铃薯的生产造成严重影响。ATP6编码ATP合酶α亚基的第6个亚单位,对ATP合酶的功能及生物的生存生殖至关重要。本研究对来自我国7个群体140株马铃薯晚疫病菌的ATP6基因进行核苷酸序列及其遗传结构和地理因素(海拔、经度和年平均气温)的相关分析。结果表明:7个群体中只有2种单倍型Hapl-1和Hapl-2,其中Hapl-1在群体中所占比例与当地的海拔呈显著正相关,与年平均气温和经度呈显著负相关,而Hapl-2在群体中的比例与当地的海拔、年平均气温和经度的相关正好与Hapl-1相反。群体间的遗传分化度(FST)与群体间直线距离呈显著正相关(r=0.659,P=0.001)。本研究表明,马铃薯致病疫霉ATP6基因单倍型分布和群体地理因素之间具有显著相关关系,该结果为了解致病疫霉的进化趋势和致病疫霉的防治提供了理论依据。
关键词:马铃薯晚疫病菌;ATP6基因;单倍型多样性;地理因素;核苷酸序列
中图分类号: S435.32文献标志码: A文章编号:1002-1302(2017)08-0075-04
马铃薯晚疫病是由致病疫霉(Phytophthora infestans de Bary)引起的毁灭性病害,可侵染马铃薯和番茄等茄科植物[1-2],每年造成直接经济损失高达百亿美元[3-4]。21世纪以来,随着马铃薯种植面积的不断扩大,我国马铃薯种植面积和产量已居世界第一,马铃薯已经成为全球性食物,是水稻、小麦和玉米之后的全球第四大粮食作物[5-6]。
线粒体基因组的突变率低,单亲遗传,常被用于研究生物的进化模式[7-10]。目前为止,已有超过150个植物、动物、真菌、原生生物的线粒体基因组被测序[11]。因此,通过了解线粒体基因遗传多样性有助于了解致病疫霉群体结构和变异特点,进而探究不同地理区域下的群体遗传结构。
马铃薯生产具有较强的地域性,在全国不同地区形成各具特色的栽培方式和栽培类型,从生产上来看,中国马铃薯的生产栽培区域为4个,分别是北方一季作区、中原二季作区、南方冬作区、西南混作区[12],各地理区域气候特征差异较大。胡珍珠等对我国北方马铃薯主产区致病疫霉群体遗传结构进行分析,结果表明,我国北方马铃薯生产区致病疫霉群体存在较为丰富的遗传多样性,且其遗传多样性与地理来源密切相关[13];云南马铃薯晚疫病群体遗传多样性在地理分布上也存在差异显著[14];气候变暖对马铃薯晚疫病发生发展的影响研究表明马铃薯晚疫病的發生发展与流行的适应气象条件,单因子作用并不显著[15];短暂的温度起伏也会导致个体产生不同的表型[16];同时,随着地区年均温的改变加剧会导致更多的遗传变异[17-18];经纬度、海拔、年平均气温、降水量以及紫外强度对当地马铃薯晚疫病菌的发生造成很大的影响[19-21]。本研究对我国7个不同地理环境的致病疫霉菌群体的ATP6基因进行遗传多样性分析,以探究致病疫霉遗传多样性的空间分布以及遗传结构与地理因素(海拔、经度和年均温)的相关性,以期为晚疫病的防控提供理论依据。
1材料与方法
1.1供试菌株
根据SSR基因型的不同从源自我国云南曲靖(2010年)、广西南宁(2010年)、贵州安顺(2011年)、福建福州(2010年)、福建霞浦(2011年)、甘肃定西(2010年)和宁夏渭源(2010年)7个马铃薯栽培地区,共计815株致病疫霉菌中选取140株无性系,寄主均为马铃薯。
1.2方法
1.2.1样本的采集与纯化在发病区选1块田地,病株间隔2 m以上,每个病株上选择具有典型症状的单个病斑的病叶,在水琼脂培养基上保湿培养12~24 h,用接菌针挑取单根菌丝,接种于含抗生素(利福平10 mg/L、氨苄青霉素100 mg/L)的选择性黑麦培养基上[22]。
1.2.2培养基的制备黑麦培养基的配置:将50 g黑麦浸泡12 h,匀浆机粉碎,60 ℃水浴浸泡2 h,4层纱布过滤,定容至1 L并且加入12 g琼脂糖加热溶化,于121 ℃高压灭菌 20 min。冷却到60 ℃左右倒平板,接菌后在18 ℃避光培养 7~10 d。将纯化的菌株接种到黑麦冻存管或者黑麦培养基斜面上,13 ℃避光长期保存菌种[23]。
1.2.3致病疫霉DNA的提取和线粒体ATP6基因的扩增收集在黑麦培养基中培养10 d的致病疫霉菌丝,冷冻干燥并且粉碎后使用BIOMIGA试剂公司订购的Bio MIGA Plant g DNA Kit(GD2611-02 250 poeps)试剂盒提取DNA,将提取的基因组DNA保存于-40 ℃备用。利用DNAMAN以及Primer Premier 5软件设计并在上海铂尚生物技术有限公司合成引物(F,5′-GAAGCTGCTGCATGGTATTGG-3′;R,5′-GCGACCTATAGCGTCACAAGC-3′),对致病疫霉7个群体的ATP6基因片段进行扩增。PCR采用Blend Taq DNA聚合酶进行扩增,PCR反应在Life Pro Themeral Cycler PCR仪上进行。反应体系为25 μL,包括1 μL模板DNA、0.25 μL Taq DNA聚合酶、2.5 μL dNTP、2.5 μL 10×buffer,加无菌水补至25 μL。PCR扩增程序为:94 ℃ 3 min;94 ℃ 30 s,55 ℃ 50 s,72 ℃ 90 s,循环35次;72 ℃ 10 min。将PCR产物进行1.2%琼脂糖凝胶电泳后,利用G:BOX凝胶成像系统进行观察拍照。
1.2.4测序和数据处理将琼脂糖凝胶电泳检测结果中条带单一的PCR扩增产物样品送于上海铂尚生物有限公司进行PCR产物测序。将测序所获得的序列通过DNAMAN软件进行拼接,使用MEGA 5.05[24]软件中的Muscle(Codons)子程序进行多重比对。通过NCBI在线BLAST比对验证同源性,利用DnaSP 5.10.1[25]软件分析致病疫霉遗传多样性参数来评估遗传多样性,包括单倍型数(number of haplotypes detected,H)、核苷酸多样性(nucleoticide diversity,π)、单倍型多样性(haplotype diversity,Hd)、多态性位点数(number of segregeting site,S)等[26-27]。利用Arlequin v 3.4.1.3[28]软件计算群体间FST,根据FST的值来判断群体分化程度。采样地点当年的年平均气温、海拔以及经度则通过World Climate(http://www.world-climate.com)以及Google Earth(http://www.earthol.com/)来查询[29],同时利用经纬度距离计算器来计算采样点之间的直线距离。
2结果与分析
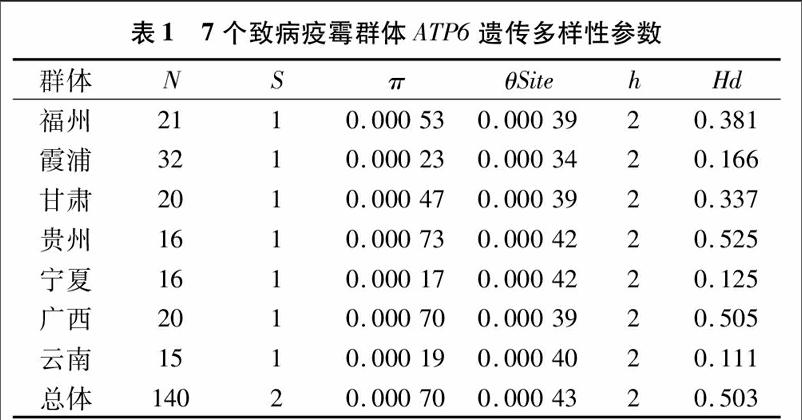
全国7个群体中只出现2个多态性位点(表1)。单倍型多样性(Hd)在地区间存在差异,福州、霞浦、甘肃、宁夏、云南群体的单倍型多样性(Hd)值均小于0.5,核苷酸多样性(π)值均小于0.005,然而贵州、广西群体的单倍型多样性值大于0.5,但是核苷酸多样性值仍然小于0.005。通过对7个群体的单倍型多样性比较发现,贵州群体的单倍型多样性平均值最高,为0.525,云南群体的单倍型多样性平均值最低,为 0.111。贵州群体ATP6基因表现出较高的遗传多样性,云南群体ATP6基因表现出较低的遗传多样性(表1)。
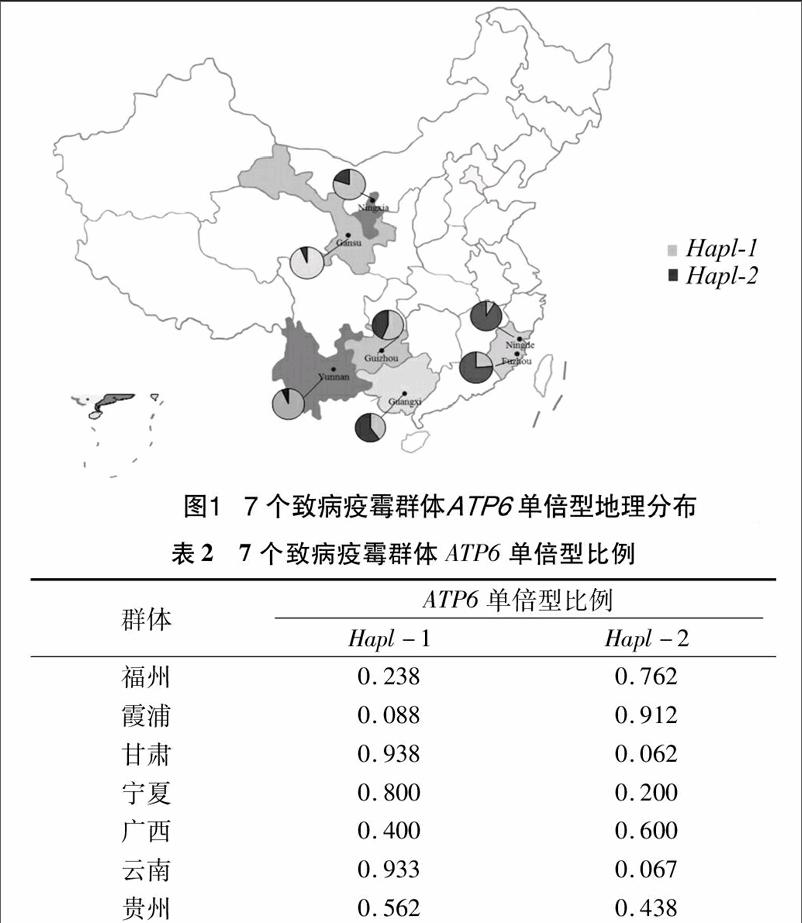
采样地点地理因素因子(表4)以及不同地区单倍型分布比例(图3)的相关性分析表明,Hapl-1比例与采样地区海拔呈现正相关(r=0.951,P=0.001),Hapl-2比例和采样地区海拔呈现负相关(r=-0.951,P=0.001),说明随着采样地区海拔的升高,Hapl-1比例越高,Hapl-2比例越低;Hapl-1比例和采样地区年平均气温呈现负相关(r=-0.752,P=0.051),而Hapl-2比例和采样地区年平均气温呈现正相关(r=0.752,P=0.051),说明Hapl-1比例随着采样地区年平均气温升高而降低,Hapl-2比例随着年平均气温升高而升
高;Hapl-1比例与采样地区经度呈现负相关(r=-0.894,P=0.007),Hapl-2比例与采样地区经度呈现正相关(r=0894,P=0.007),说明随着采样地区采样地区经度的升高,
3讨论与结论
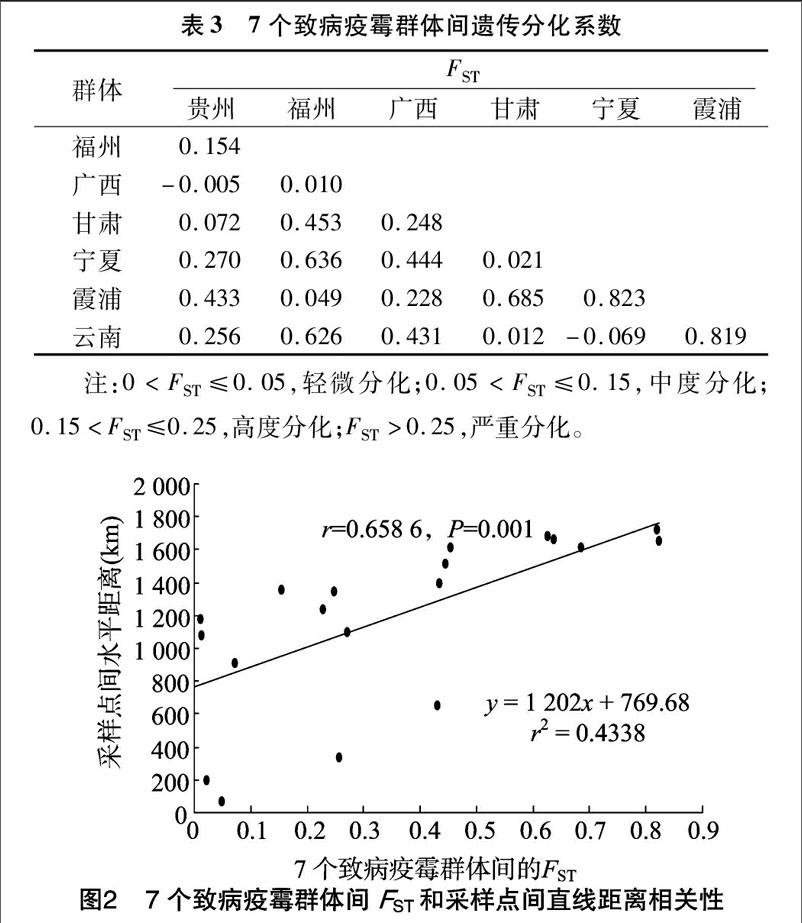
对我国7个不同群体采集筛选的共计140个致病疫霉ATP6基因进行扩增和核苷酸分析,同时进行ATP6基因遗传结构和部分地理因素之间的相关性分析。结果表明,有2种单倍型Hapl-1和Hapl-2,其中Hapl-1与海拔呈正相关,与年平均气温和经度呈负相关,而Hapl-2与海拔呈负相关,与年平均气温和经度呈正相关。群体间的FST与群体间水平距离成显著相关关系。
有研究表明,地理环境因素是影响晚疫病流行的关键因素[30-32],本研究结果也表明,ATP6基因单倍型比例与采样地区海拔、经度以及年平均气温具有相关性。同时ATP6基因与呼吸作用相关[33],海拔、年均氣温和经度均可以影响生物的呼吸作用,本试验间接证明这些地理因素与晚疫病发生和致病疫霉群体分布具有相关性。
从生产上来看,中国马铃薯的生产栽培区域有各自特有的气象因素和农业生态系统,各生态区域气候特征差异较大。地区间致病疫霉群体遗传结构各有不同。群体间FST和采样点间垂直距离呈现正相关(r=0.659,P=0.001)。这一结果证实,进化理论中提到的地区间隔是导致种群地区间遗传分化的重要原因,地理位置越靠近分化程度越低[34-35]。
目前我国对致病疫霉遗传分化的原因多处于对遗传多样性的了解,对地区间分化原因以及形成机制很少探讨。本研究利用分子生物学和群体遗传学原理,初步分析我国不同地区马铃薯晚疫病菌的遗传多样性及形成的可能机制,为马铃薯防治提供依据。但本研究选择单个线粒体基因,在未来研究中,采取多个基因联合方法来对此结论进行进一步验证,为制定长期、有效和环境友好型的致病疫霉防治方案提供科学依据。
参考文献:
[1]Turkensteen L J. Durable resistance of potatoes against Phytophthora infestans[M]//Jacob T,Parlevliet J E. Durability of disease resistance. Berlin:Springer Netherlands,1993:115-124.
[2]Erwin D C,Ribeiro O K. Phytophthora diseases worldwide[J]. Plant Pathology,1998,47(2):224-225.
[3]李汝刚. 马铃薯抗晚疫病遗传工程研究获得重大突破[J]. 植物保护,1997,23(5):12-13.
[4]Fry W E,Goodwin S B. Re-emergence of potato and tomato late blight in the United States[J]. Plant Disease,1997,81(12):1349-1357.
[5]Birch P J,Bryan G,Fenton B,et al. Crops that feed the world 8:potato are the trends of increaed global production sustainable?[J]. Food Security,2012,4(4):477-508.
[6]杨丽娜,段国华,覃雁瑜,等. 2012年云南省会泽县马铃薯晚疫病菌小种结构分析[J]. 热带作物学报,2016,37(1):158-163.
[7]Avila-Adame C,Gómez-Alpizar L,Zismann V,et al. Mitochondrial genome sequences and molecular evolution of the Irish potato famine pathogen,Phytophthora infestans[J]. Current Genetics,2006,49(1):39-46.
[8]Segawa R D,Aotsuka T. The mitochondrial genome of the Japanese freshwater crab,Geothelphusa dehaani(Crustacea:Brachyura):evidence for its evolution via gene duplication[J]. Gene,2005,355(355):28-39.
[9]Rogaev E I,Moliaka Y K,Malyarchuk B A,et al. Complete mitochondrial genome and phylogeny of Pleistocene mammoth Mammuthus primigenius[J]. PLoS Biology,2006,4(3):e73.
[10]Boore J L. Complete mitochondrial genome sequence of Urechis caupo,a representative of the phylum Echiura[J]. BMC Genomics,2004,5(1):67.
[11]Chesnick J M,Goff M,Graham J,et al. The mitochondrial genome of the stramenopile alga Chrysodidymus synuroideus. Complete sequence,gene content and genome organization[J]. Nucleic Acids Research,2000,28(13):2512-2518.
[12]Wang B,Ma Y,Zhang Z,et al. Potato viruses in China[J]. Crop Protection,2011,30(9):1117-1123.
[13]胡珍珠,杨志辉,丁明亚,等. 我国北方马铃薯生产区致病疫霉群体遗传结构分析[J]. 河南农业科学,2015,44(7):83-88.
[14]赵志坚,曹继芬,杨明英,等. 用两个微卫星标记分析云南马铃薯晚疫病菌的遗传多样性[J]. 中国农业科学,2008,41(11):3610-3617.
[15]姚晓红,许彦平,姚新琪. 气候变暖对马铃薯晚疫病发生发展的影响[J]. 安徽农业科学,2010,38(21):11315-11317,11444.
[16]Winkler W C,Breaker R R. Regulation of bacterial gene expression by riboswitches[J]. Annual Review of Microbiology,2005,59(59):487-517.
[17]Lee C E. Evolutionary genetics of invasive species[J]. Trends in Ecology & Evolution,2002,17(8):386-391.
[18]Somero G N. The physiology of climate change:how potentials for acclimatization and genetic adaptation will determine ‘winners and ‘losers[J]. The Journal of Experimental Biology,2010,213(6):912-920.
[19]Alizon S,Hurford A,Mideo N,et al. Virulence evolution and the trade-off hypothesis:history,current state of affairs and the future[J]. Journal of Evolutionary Biology,2009,22(2):245-259.
[20]Altizer S,Ostfeld R S,Johnson P T,et al. Climate change and infectious diseases:from evidence to a predictive framework[J]. Science,2013,341(6145):514-519.
[21]Baumann H,Conover D O. Adaptation to climate change:contrasting patterns of thermal-reaction-norm evolution in Pacific versus Atlantic silversides[J]. Proceedings Biological Sciences,2011,278(1716):2265-2273.
[22]Wu E J,Yang L N,Zhu W,et al. Diverse mechanisms shape the evolution of virulence factors in the potato late blight pathogen Phytophthora infestans sampled from China[J]. Scientific Reports,2016,6:26182.
[23]吳娥娇,沈国桢,刘训达,等. 水杨肟酸对致病疫霉生长及其对嘧菌酯敏感性的影响[J]. 农药学学报,2015,17(4):384-390.
[24]Tamura K,Peterson D,Peterson N,et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J]. Molecular Biology and Evolution,2011,28(10):2731-2739.
[25]Sgarbi G,Baracca A,Lenaz G,et al. Inefficient coupling between proton transport and ATP synthesis May be the pathogenic mechanism for NARP and Leigh syndrome resulting from the T8993G mutation in mtDNA[J]. The Biochemical Journal,2006,395(3):493-500.
[26]Kung M L,Ga H B,James A C,et al. Distribution patterns and introduction pathways of the Cosmopolitan brown alga Colpomenia peregrina using mtcox3 and atp6 sequences[J]. Journal of Applied Phycology,2014,26(1):491-504.
[27]Cárdenas M,Grajales A,Sierra R,et al. Genetic diversity of Phytophthora infestans in the Northern Andean region[J]. BMC Genetics,2011,12(1):23.
[28]Excoffier L,Lischer H E. Arlequin suite ver 3.5:a new series of programs to perform population genetics analyses under Linux and Windows[J]. Molecular Ecology Resources,2010,10(3):564-567.
[29]Yang L N,Zhu W,Wu E J,et al. Trade-offs and evolution of thermal adaptation in the Irish potato famine pathogen Phytophthora infestans[J]. Molecular Ecology,2016,25(16):4047-4058.
[30]譚宗九,王文泽,丁明亚,等. 气象因素对马铃薯晚疫病发生流行的影响[J]. 中国马铃薯,2001,15(2):96-98.
[31]王利亚,孙茂林,杨艳丽,等. 云南马铃薯晚疫病区域性流行学的研究[J]. 西南农业学报,2005,18(2):157-162.
[32]池再香,杜正静,陈忠明,等. 2009—2010年贵州秋、冬、春季干旱气象要素与环流特征分析[J]. 高原气象,2012,31(1):176-184.
[33]杨福愉. ATP合酶:一个最小的蛋白质分子转动马达[J]. 医学分子生物学杂志,2005,2(4):243-249.
[34]祝雯,付海静,杨丽娜,等. 福建省部分地区马铃薯治病疫霉群体遗传多样性分析[J]. 激光生物学报,2013,22(3):267-272.
[35]祝雯,詹家绥. 植物病原物的群体遗传学[J]. 遗传,2012,34(2):157-166.
猜你喜欢
中学政史地·高中文综(2023年3期)2023-04-14
中学政史地(2023年8期)2023-04-14
今日农业(2021年14期)2021-10-14
今日农业(2021年7期)2021-07-28
绵阳师范学院学报(2021年5期)2021-05-28
今日农业(2020年15期)2020-09-25
河北大学学报(自然科学版)(2020年1期)2020-01-15
广东农业科学(2017年5期)2017-08-29
中国马铃薯(2015年3期)2015-12-19
世界科学(2014年8期)2014-02-28
