
不同穗型小麦小花生长发育及同化物分配变化的差异
2017-06-27 08:11:53郑春风孙克刚朱云集
麦类作物学报 2017年6期
郑春风,任 伟,孙克刚,朱云集
(1.河南省农业科学院植物营养与资源环境研究所,河南郑州 450002;2.河南省农业科学院小麦研究所,河南郑州 450002; 3.河南农业大学/国家小麦工程技术研究中心,河南郑州 450002)
不同穗型小麦小花生长发育及同化物分配变化的差异
郑春风1,任 伟2,孙克刚1,朱云集3
(1.河南省农业科学院植物营养与资源环境研究所,河南郑州 450002;2.河南省农业科学院小麦研究所,河南郑州 450002; 3.河南农业大学/国家小麦工程技术研究中心,河南郑州 450002)
为了解小麦小花发育成粒特性与同化物分配间的关系,以大穗型品种周麦16和多穗型品种豫麦49-198为材料,分析了两种穗型小麦小花发育成粒特性及同化物分配的差异。结果表明,周麦16的小花退化、败育速率较豫麦49-198缓慢,且在小穗结实率、小花结实率和可孕小花结实率上均高于后者。在小花发育中后期,大穗型品种周麦16转移到穗部的同化物比多穗型品种豫麦49-198多。经相关分析,花期完善小花(有完整绿色花药的小花)数与穗干重、穗氮积累量、茎干重、茎氮积累量呈微弱的正相关,与穗茎干重比、穗茎氮积累量比呈显著正相关,与穗干重/穗氮积累量比呈极显著正相关。周麦16的穗茎干重比、穗茎氮积累量比显著高于豫麦49-198,二指标均与穗干重/穗氮积累量比呈极显著正相关。进一步分析发现,周麦16的开花期单位面积完善小花数与成熟期单位面积粒数分别较豫麦49-198增加3.85%和4.65%,且品种间差异均达到极显著水平。
小麦;穗型;小花发育;结实性;同化物分配
在当前中高产水平下,需在稳定穗数的前提下,把增加粒重和穗粒数作为进一步提高小麦产量的突破口[1-4]。但由于环境条件和品种遗传特性的制约,增加粒重的空间相对比较有限,穗粒数的提高应成为提高小麦产量的关键[5-8]。由于小麦穗粒数的形成是小花发育、结实成粒等一系列复杂生理过程的最终体现[9-10],因此明确小花发育结实特性和生理变化特征,探明小花发育结实成粒的规律,对于深化小麦产量形成生理的研究和提高穗粒数,具有十分重要的理论和实际意义。李存东等[11]研究表明,小麦的小花发育模式与品种、蘖位及播后的积温关系密切。王兆龙等[12]研究认为,小麦小花的分化动态符合二次曲线方程,小花的退化动态符合一次方程,不同穗型品种在方程参数上有明显区别。特大穗型小麦小花分化速率快,每穗分化小花数多,形成的可孕小花数和结实粒数最多。不同穗型小麦的小穗结实率、小花结实率和可孕花结实率均为穗型越大,结实率越高,其中以可孕花结实率的极差最为明显。不同穗型小麦的强势位小花在结实特性上差异较小,但弱势位小花依穗型不同有明显的极差。特大穗型和大穗型小麦在幼穗分化后期输入穗中的干物质相对较多,穗茎干重比较高,因而穗中较高的干物质输入是大穗形成的物质基础。朱云集等[14]指出,合理施用氮肥能够增加小花的数目,减少小花退化,增加穗粒数。Ariel Ferrante等[15]也发现,硬粒小麦可孕花结实成粒与氮肥有效性密切相关。Demotes等[16]的研究结果显示,小麦小花发育期的穗部氮素积累量与籽粒数呈显著正相关。然而,有关两个不同穗型小麦品种的小花发育结实成粒特性和同化物分配的差异性以及两者间的相互关系鲜见研究报道。本试验重点研究了大穗型和多穗型小麦品种在小花发育成粒特性和同化物分配变化特征上的差异,以期揭示小麦小花发育结实成粒与同化物分配间的关系,为大穗型小麦品种的培育和高产栽培调控提供理论依据。
1 材料与方法
1.1 试验材料与设计
试验于2013-2014年在河南农业大学科教示范园区(34°86′N,113°59′E)进行,田间土壤为壤土,0~20 cm土层有机质含量为10.6 g·kg-1,全氮含量为0.9 g·kg-1,碱解氮含量为82.1 mg·kg-1,速效磷含量为25.6 mg·kg-1,速效钾含量为124.5 mg·kg-1,有效硼含量为0.41 mg·kg-1。总施氮量146 kg·hm-2(尿素,含N 46%),其中50%于播前基施,其余50%于拔节期追施。播前每公顷施磷肥(P2O5)150 kg和钾肥(K2O)120 kg。以大穗型品种周麦16、多穗型品种豫麦49-198为供试材料,均于10月8号播种,基本苗为2.25×106株·hm-2,行距为20 cm,各小区面积为20 m2,随机区组设计,重复3次。试验田栽培管理同一般高产田。
1.2 测定内容与方法
自小麦3叶期开始取样,每隔7 d取样1次,以有完整绿色花药的小花为完善小花,每小区选择生长均匀一致的小麦5株(每处理共计15株),在EMZ-TR解剖镜下观察记载主茎幼穗分化进程,并观察记载分化小穗数、小花数及幼穗分化各阶段的特征。
播后累积生长度日GDD=∑ [(Tmax+Tmin)/2-Tb],Tmax为最高温度,Tmin为最低温度,Tb为基础温度。
于拔节期、拔节后7 d、拔节后14 d、拔节后21 d、拔节后28 d、孕穗期、抽穗期、开花期、成熟期取样,105 ℃下杀青30 min,80 ℃烘干至恒重,粉碎保存,用于植株干物质和氮含量的测定。
成熟期每小区随机取20株,按常规考种法记载每穗结实小穗数、每穗结实粒数和每小穗不同小花位结实粒数。每小区实收5 m2计产。
1.3 数据分析
采用Microsoft Excel 2003和SPSS 17.0软件对数据进行处理分析及绘图。
2 结果与分析
2.1 两种穗型小麦品种的小花发育动态和结实性的差异
两个小麦品种小花发育均可分为小花分化、小花退化及小花败育3个阶段。由图1可看出,小麦单位面积分化小花数随播后GDD的增加呈先逐渐上升、再迅速下降、后缓慢下降的变化过程。两个品种小花分化数均在GDD达到1 150 ℃时最高,且周麦16的分化小花数明显多于豫麦49-198。在小花分化阶段,两个品种小花分化速度均由快到慢;在小花退化和败育阶段,周麦16小花退化速度和败育速度均较豫麦49-198缓慢,最终的可孕小花数较豫麦49-198高(表1)。

表1 两种穗型小麦品种单位面积小花发育的动态模式方程Table 1 Model equations of floret development per unit area of two wheat varieties
**:P<0.01.
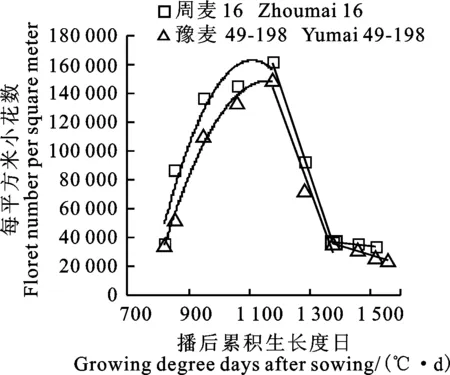
图1 两种穗型小麦品种单位面积小花发育动态变化的差异
周麦16单位面积的小穗结实率、小花结实率和可孕小花结实率均高于豫麦49-198,其中可孕小花结实率在品种间差异显著(图2),表明可孕小花结实率与最终的穗粒数关系密切。
由图3可看出,周麦16小穗上各小花位结实粒数均高于豫麦49-198。两品种在小穗上第1、第2朵强势花位的小花结实粒数差异相对较小,但在第3、第4朵弱势花位的小花结实粒数差异较大,说明弱势花位小花结实粒数的多少对大穗的形成起着重要的作用。
2.2 两种穗型小麦品种的小花发育过程中同化物分配的差异
在拔节后14 d之前,周麦16的穗茎干物重比(茎包含茎、叶、鞘,下同)与豫麦49-198无显著差异。拔节后14 d之后,周麦16的穗茎干物重比显著高于豫麦49-198(图4)。表明在小花发育中后期,大穗型小麦品种周麦16转移到穗部的同化物比多穗型小麦品种豫麦49-198多。

图柱上不同字母表示品种间差异显著(P<0.05)。图5同。
Different letters above columns mean significant difference between varieties(P<0.05).The same in Fig.5.
图2 两种穗型小麦品种间小穗结实率、小花结实率、可孕小花结实率的差异
Fig.2 Difference of spikelet setting rate,floret setting rate and grain setting rate between the two wheat varieties
在拔节后21 d之前,大穗型周麦16的穗茎氮积累量比显著低于多穗型豫麦49-198。而在拔节后21 d之后,周麦16的穗茎氮积累量比显著高于豫麦49-198(图5)。这表明在小花发育后期,大穗型小麦品种周麦16转移到穗部的氮量比多穗型小麦品种豫麦49-198多。
2.3 两种穗型小麦品种的开花期同化物分配与完善小花数及穗粒数的关系
2.3.1 开花期同化物分配与完善小花数及穗粒数的相关性
相关分析(表2)表明,开花期单位面积的完善小花数与穗干重、穗氮积累量、茎干重、茎氮积累量均呈正相关,但相关性不大;与穗茎干重比、穗茎氮积累量比均呈显著正相关;与穗干重/穗氮积累量比的相关系数达到0.9以上,呈极显著正相关。此外,穗粒数与开花期同化物在穗器官的分配量及分配比例均呈显著正相关,与完善小花数呈极显著正相关。
2.3.2 开花期同化物分配与完善小花数及穗粒数的差异
豫麦49-198的开花期单位面积的茎干重、穗氮积累量和茎氮积累量显著高于周麦16,但穗茎干重比、穗茎氮积累量比、穗干重/穗氮积累量比显著或极显著低于周麦16。进一步分析可知,周麦16在开花期单位面积完善小花数、成熟期单位面积粒数较豫麦49-198分别增加3.85%和4.65%,且差异均达到极显著水平。
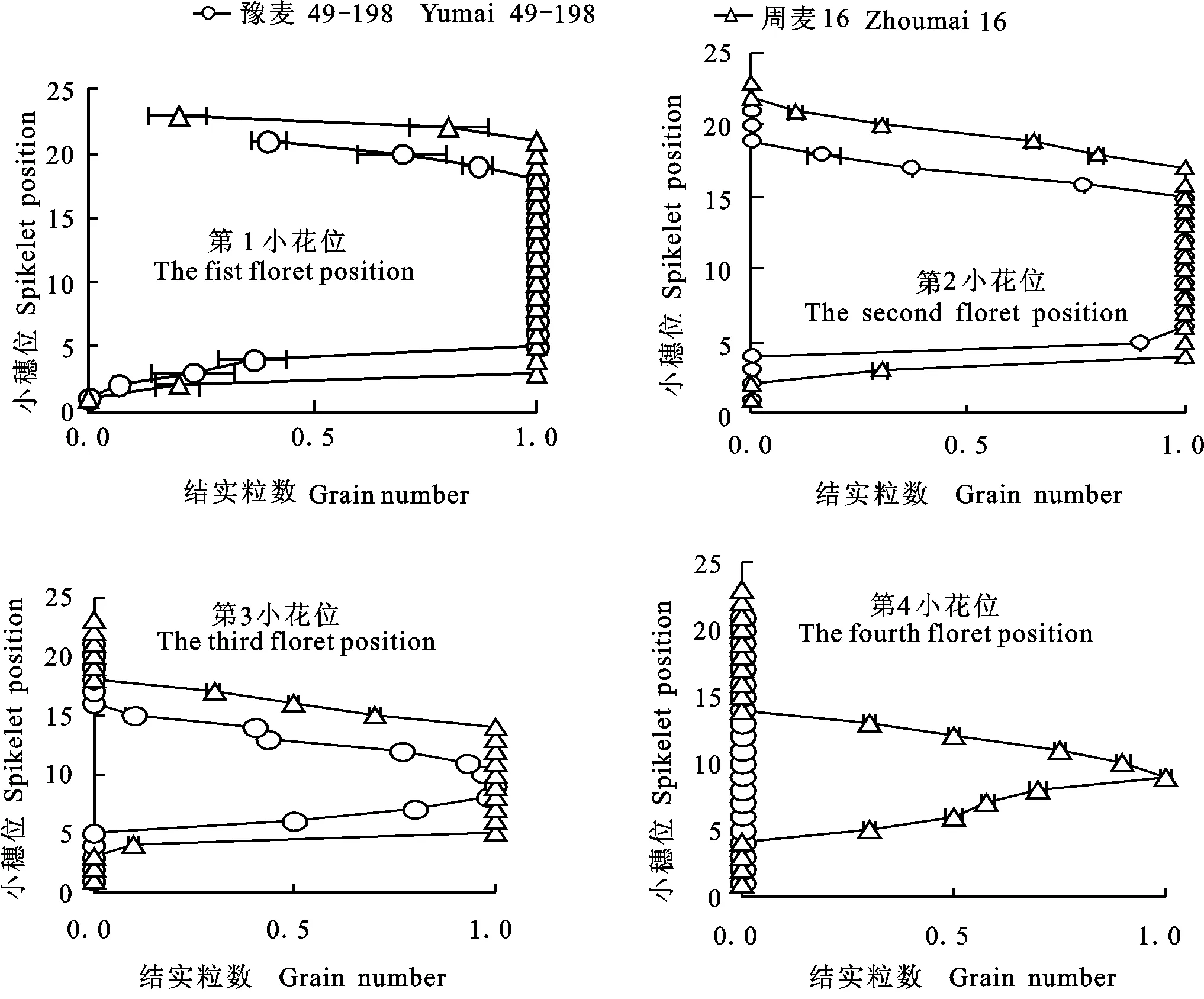
图3 小麦主茎穗上各小穗不同小花位的平均结实粒数

图4 两种穗型小麦品种主茎小花发育期穗茎干物重比的差异
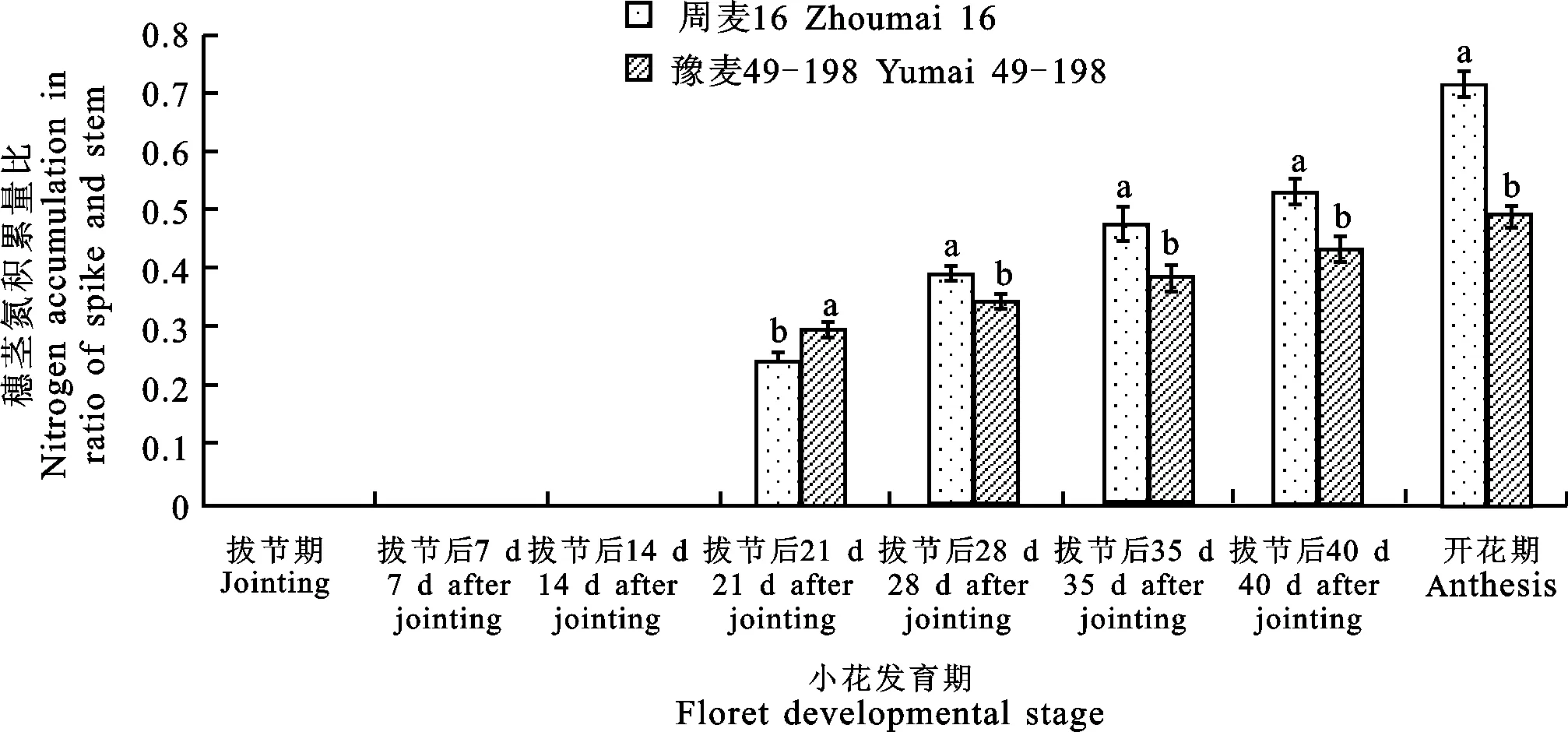
图5 两种穗型小麦品种主茎小花发育期穗茎氮积累量比的差异
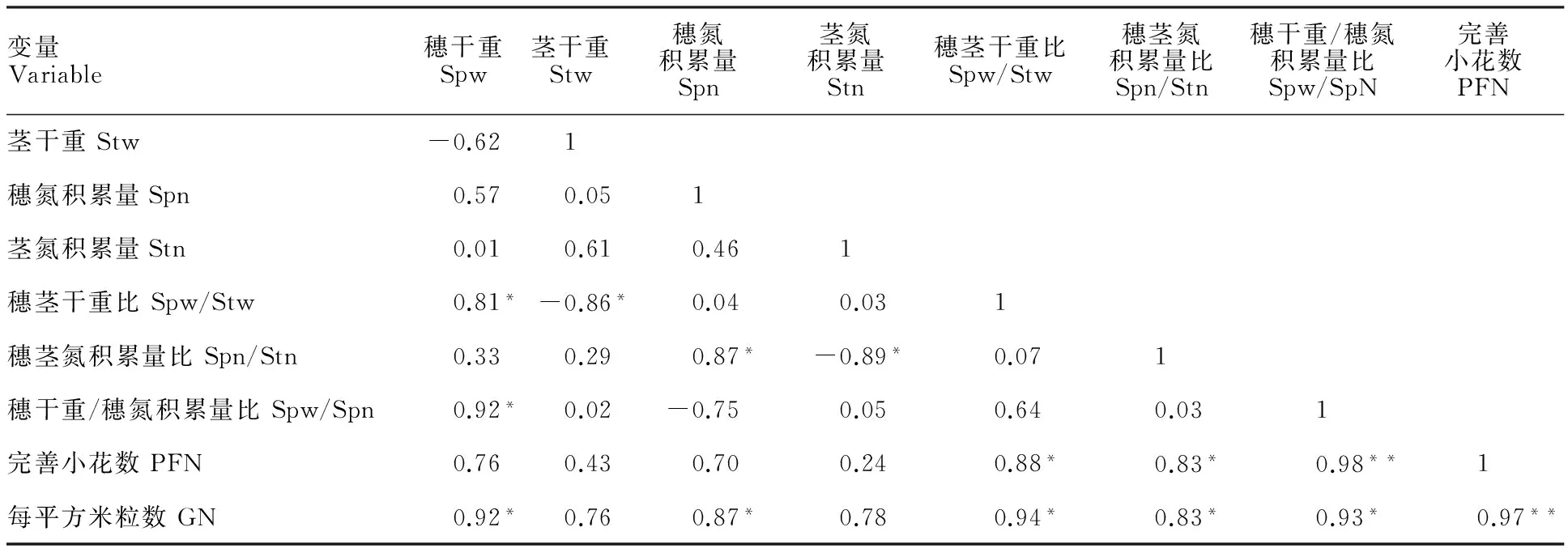
表2 开花期同化物分配与完善小花数及单位面积粒数的相关分析Table 2 Correlation analysis of photosynthate distribution,perfect floret and grain number
*:P<0.05; **:P<0.01. Spw:Spike dry weight; Stw:Stem dry weight; Spn:Spike nitrogen accumulation; Stn:Stem nitrogen accumulation; PFN:Perfect floret number; GN:Grain number.
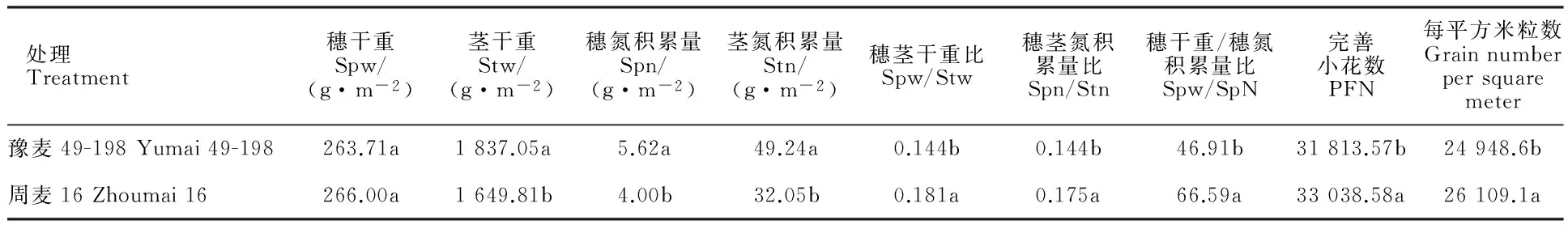
表3 开花期同化物分配与完善小花数及穗粒数的差异分析Table 3 Differential analysis of photosynthate distribution,perfect floret and grain number of two wheat varieties
同列不同小写字母表示品种间在0.05水平上的差异显著。
Different lower-case letters in the same columns indicate significant difference between varieties at 0.05 level.
3 讨论与结论
本研究表明,2个不同穗型小麦材料周麦16和豫麦49-198在小花发育、结实性上存在着明显的差异。大穗型品种周麦16的小花退化速度和败育速度均较多穗型品种豫麦49-198缓慢,并且大穗型品种周麦16最终的可孕小花数高于多穗型品种豫麦49-198,说明大穗型品种周麦16较高的穗粒数与其小花退化缓慢和可孕小花败孕较少有着密切关系。对结实率分析发现,大穗型品种周麦16的小穗结实率、小花结实率和可孕小花结实率均高于多穗型品种豫麦49-198,其中可孕小花结实率显著高于多穗型品种豫麦49-198,表明可孕小花结实性与最终的穗粒数有着密切相关。分析小穗上不同小花位小花结实性发现,大穗型品种周麦16与多穗型品种豫麦49-198在小穗上强势花位的小花结实性差异相对较小,但在弱势花位的小花结实性差异较大。由此推测,弱势花位小花结实性差异是引起大穗型和多穗型小麦品种之间穗粒数不同的一个主要原因。
在开花前,小麦的营养器官处于快速生长期,尤其是在拔节后节间迅速伸长[17-18]。Miralles等[19]认为,限制穗粒数形成的一个重要因素是幼穗与茎秆在快速生长期对同化物的激烈竞争。本研究结果发现,在拔节后14 d之后,大穗型品种周麦16的穗茎干重比显著高于多穗型品种豫麦49-198,在拔节后21 d之后,大穗型品种周麦16的穗茎氮积累量比显著高于多穗型品种豫麦49-198。表明在小花发育中后期,大穗型小麦品种周麦16有较多的同化物转移到穗部,这为大穗的形成奠定了良好的物质基础。此结果与Slafer等[20]得出形成大穗的重要物质基础是要有较多的同化物转移到穗部的研究结果相一致。Ellen等[21]指出,致使小花急剧退化的原因与碳水化合物总供给量关系不大,而与营养器官和穗部的同化物分配比例有很大联系。本研究中,开花期单位面积的完善小花数与穗干重、穗氮积累量呈正相关,但相关性不大,而与穗茎干重比、穗茎氮积累量比呈显著正相关,与穗干重/穗氮积累量比呈极显著正相关,表明开花期完善小花的形成与穗器官同化物供给总量的关系不大,而与同化物在穗部和营养器官的分配比例密切相关。此结果与Ellen等的研究结果一致。本研究进一步发现,开花期完善小花数的形成与同化物在穗干重/穗氮积累量比上的分配比例呈极显著正相关。
本研究通过比较大穗型和多穗型小麦品种在小花发育成粒特性及同化物分配变化上的差异最终发现,小花退化缓慢、可孕花败育较少、可孕花结实率较高是大穗型小麦品种获得较多穗粒数的重要原因,弱势花位小花结实率的提高是增加小麦穗粒数的关键。在小花发育中后期,大穗型品种周麦16较豫麦49-198有较多的同化物转移到穗部。开花期完善小花的形成与穗器官同化物供给总量的关系不大,而与同化物在穗部和营养器官的分配比例密切相关,与同化物在穗部的分配比例关系更加密切。通过分析初步找出了小麦小花发育结实成粒与同化物分配间的关系,为大穗型小麦品种的栽培调控提供了理论依据,但是小花发育成粒的内在生理机制有待进一步研究。
[1] 于振文,田奇卓,潘庆民,等.黄淮麦区冬小麦超高产栽培的理论与实践[J].作物学报,2002,28(5):577.YU Z W,TIAN Q Z,PAN Q M,etal.Theory and practice on cultivation of super high yield of winter wheat in the wheat fields of Yellow River and Huaihe River districts [J].ActaAgronomicaSinica,2002,28(5):577.
[2]王兆龙,曹卫星,戴廷波.小麦穗粒数形成的基因型差异及增粒途径分析[J].作物学报,2001,27(2):236.WANG Z L,CAO W X,DAI T B.Genotypic differences in formation of kernel number per spike and analysis of improvement approaches in wheat [J].ActaAgronomicaSinica,2001,27(2):236.
[3]FISCHER R A.The importance of grain or kernel number in wheat:A reply to Sinclair and Jamieson [J].FieldCropsResearch,2008,105:15.
[4]SINCLAIR T R,JAMIESON P D.Grain number,wheat yield,and bottling beer:An analysis [J].FieldCropsResearch,2006,98:60.
[5]崔金梅,郭天财,朱云集,等.小麦的穗[M].北京:中国农业出版社,2008:284-286.CUI J M,GUO T C,ZHU Y J,etal.Spike of Wheat [M].Beijing:China Agricultural Press,2008:284-286.
[6]马元喜,王晨阳,朱云集.协调小麦幼穗发育三个两极分化过程增加穗粒数[C]//卢良恕.中国小麦栽培研究新近展.北京:中国农业出版社,1991:119-126.MA Y X,WANG C Y,ZHU Y J.Coordinate three polarization process of young spike development to increase grain number[C]//LU L S.China’s Wheat Cultivation Research Progress.Beijing:China Agricultural Press,1991:119-126.
[7]王志敏.小麦穗粒数的调节[D].北京:北京农业大学,1994.WANG Z M.Regulation of grain number in wheat [D].Beijing:Beijing Agricultural University,1994.
[8]郑春风,朱慧杰,朱云集,等.冬小麦小花发育及结实特性对叶面喷硼的响应[J].植物营养与肥料学报,2016,22(2):550.ZHENG C F,ZHU H J,ZHU Y J,etal.Responses of floret development and grain setting characteristics of winter wheat to foliar spray boron [J].JournalofPlantNutritionandFertilizer,2016,22(2):550.
[9]ZHENG C F,ZHU Y J,ZHU H J,etal.Floret development and grain setting characteristics in winter wheat in response to pre-anthesis foliar applications of 6-benzylaminopurine and boron [J].FieldCropsResearch,2014,169:70.
[10]FERRANTE A,SAVIN A,SLAFER G A.Floret development of durum wheat in response to nitrogen availability [J].JournalofExperimentalBotany,2010,61:4351.
[11 ]LI C D,CAO W X,DAI T B.Dynamic characteristics of floret primordium development in wheat [J].FieldCropsResearch,2001,71:71.
[12]王兆龙,曹卫星,戴廷波,等.不同穗型小麦品种小花发育与结实特性研究[J].南京农业大学学报,2000,23(4):9.WANG Z L,CAO W X,DAI T B,etal.Characteristics of floret development and grain set in three wheat genotypes of different spike sizes [J].JournalofNanjingAgriculturalUniversity,2000,23(4):9.
[13]王俊英,赵春江,杨宝祝.小麦小花发育与退化的研究[J].华北农学报,1996,11(2):9.WANG J Y,ZHAO C J,YANG B Z.A study of florets differentiation development and degeneration in winter wheat [J].ActaAgriculturaeBoreali-Sinica,1996,11(2):9.
[14]朱云集,崔金梅,王晨阳,等.小麦不同生育时期施氮对穗花发育和产量的影响[J].中国农业科学,2002,35(11):1325.ZHU Y J,CUI J M,WANG C Y,etal.Effects of nitrogen application at different wheat growth stages on floret development and grain yield of winter wheat [J].ScientiaAgriculturaSinica,2002,35(11):1325.
[15]FERRANTE A,SAVIN A,SLAFER G A.Floret development and grain setting differences between modern durum wheat under contrasting nitrogen availability [J].JournalofExperimentalBotany,2013,64:169.
[16]DEMOTES-MAINARD S,JEUFFROY M H.Effects of nitrogen and radiation on dry matter and nitrogen accumulation in the spike of winter wheat [J].FieldCropsResearch,2004,87:221.
[17]GONZALEZ F G,SLAFER G A,MIRALLES D J.Floret development and spike growth as affected by photoperiod during stem elongation in wheat [J].FieldCropsResearch,2003,81:29.
[18]SLAFER G A,ABELEDO L G,MIRALLES D J,etal.Photoperiod sensitivity during stem elongation as an avenue to raise potential yield in wheat [J].Euphytica,2001,119:191.
[19]MIRALLES D J,KATZ S D,COLLOCA A,etal.Floret development in near isogenic wheat lines differing in plant height [J].FieldCropsResearch,1998,59:21.
[20]SLAFER G A,ANDRADE F H.Physiological attributes to the generation of grain yield in bread wheat cultivars released at different eras [J].FieldCropsResearch,1993,31:351.
[21]ELLEN J.Effects of nitrogen and plant density on growth,yield and chemical composition of two winter wheat cultivars [J].AgronomyandCropScience,1990,164:174.
Differential Analysis of Floret Growing and Development and Photosynthate Distribution in Wheat Varieties with the Different Spike Types
ZHENG Chunfeng1,REN Wei2,SUN Kegang1,ZHU Yunji3
(1.Institute of Plant Nutrition Agricultural Resources and Environmental Science,Henan Academy of Agricultural Science,Zhengzhou,Henan 450002,China; 2.Institute of Wheat,Henan Academy of Agricultural Science,Zhengzhou,Henan 450002,China; 3.Henan Agricultural University/National Engineering Research Center for Wheat,Zhengzhou,Henan 450002,China)
This study explored the difference of floret development and photosynthate distribution between large-spike and multiple spike type wheat varieties. The results showed that the abortion and degradation rate of floret of Zhoumai 16 with large spike were slower than those of Yumai 49-198 with multiple spike,and the final fertile floret number of Zhoumai 16 was higher than that of Yumai 49-198. The setting rates of spikelet,floret and fertile floret of Zhoumai 16 were also higher than those of Yumai 49-198. Analysis on the difference of photosynthate distribution during floret development found that at late period of floret development,compared to Yumai 49-198,more photosynthate transferred to the spike in Zhoumai 16. It was further found that,the number of perfect florets and grains per unit area of Zhoumai 16 were significantly increased by 3.85% and 4.65%,compared to that of Yumai 49-198,respectively.
Wheat; Spike type; Floret development; Grain-setting characteristic; Photosynthate distribution
10.7606/j.issn.1009-1041.2017.06.11
时间:2017-06-07
2016-12-19
2017-02-04
国家自然科学基金项目(31571607);河南省财政预算项目(2015BAD23B02,2013BAD07B07,YCY20167817);农业部行业专项(201303103)
E-mail:zhengfeng198822@163.com(郑春风);E-mail:hnxxrenwei@163.com(任 伟,与
孙克刚(E-mail:kgsun@ipni.ac.cn)
S512.1;S311
A
1009-1041(2017)06-0801-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170607.1005.022.html
第一作者同等贡献)
猜你喜欢
扬州大学学报(农业与生命科学版)(2023年4期)2024-01-09 13:54:56
Computer Modeling In Engineering&Sciences(2022年6期)2022-07-02 07:43:24
种子(2022年1期)2022-02-24 04:30:50
河南农业科学(2021年10期)2021-12-03 08:48:42
种子(2020年9期)2020-10-22 13:22:48
中国种业(2019年6期)2019-06-25 02:28:44
江苏农业科学(2017年17期)2017-11-15 02:46:54
植物营养与肥料学报(2013年1期)2013-10-26 03:00:24
农家参谋·种业大观(2013年7期)2013-04-29 08:55:48
农家参谋·种业大观(2013年9期)2013-04-29 00:44:03
