CXCR4基因转染对骨髓间充质干细胞体外生物学行为的影响
2017-06-22 14:50王玉莹李秀丽王雅萌李少恒闫宇辉杨静娴闻庆平
中国药理学通报 2017年6期
王玉莹,张 囡,李秀丽,王雅萌,李少恒,闫宇辉,宋 捷,杨静娴,闻庆平
(1.辽宁中医药大学药学院药理学教研室,辽宁 大连 116600;2.大连医科大学,辽宁 大连 116044)
CXCR4基因转染对骨髓间充质干细胞体外生物学行为的影响
王玉莹1,张 囡1,李秀丽1,王雅萌1,李少恒1,闫宇辉1,宋 捷1,杨静娴1,闻庆平2
(1.辽宁中医药大学药学院药理学教研室,辽宁 大连 116600;2.大连医科大学,辽宁 大连 116044)
目的 观察对转染CXCR4基因的骨髓间充质干细胞体外生物学行为的影响。方法 将培养的骨髓间充质干细胞分为3组,GFP组、CXCR4+组、CXCR4-组,转染趋化因子受体CXCR4,并用免疫荧光细胞化学法、流式细胞仪法及Transwell小室细胞趋化实验,体外研究了CXCR4高表达及低表达对MSCs增殖、分化与迁移能力的影响。结果 CXCR4高表达及低表达均不影响MSCs的增殖能力,对其向肺组织分化的能力也没有影响。与GFP-MSCs组相比,CXCR4+-MSCs组迁移的细胞数量明显升高,而CXCR4--MSCs组迁移细胞数量差异无显著性。结论 CXCR4高表达及低表达不改变MSCs的增殖、分化能力,然而CXCR4高表达明显增强MSCs向炎症病灶的迁移能力。说明CXCR4高表达的MSCs移植入体后将会更快速、更大量地到达病变区域参与组织修复,明显增强疗效。
骨髓间充质干细胞;趋化因子受体CXCR4;基因转染;增殖;分化;迁移
间充质干细胞(mesenchemal stem cell,MSCs)是一种取材广泛的成体干细胞,可取自骨髓、脂肪组织、骨骼肌、滑膜、脐带血等组织[1-2],具有多向分化潜能,可分化为成骨细胞、脂肪细胞、肌肉细胞、神经系细胞等[3-5],其中骨髓来源的MSCs有取材简单、来源丰富的特点。骨髓来源的MSCs可以减轻系统炎症、减轻肺组织受损情况、促进模型动物存活,对脂多糖(lipopolysaccharides,LPS)诱导的小鼠急性肺损伤有明显的治疗作用[6]。此外,其在体外与肺组织细胞或肺组织条件培养基共培养可以分化成肺组织相关细胞[7-8],从组织再生的角度为临床上治疗急性肺损伤(acute lung injury,ALI)提供新的思路。然而,体外扩增的MSC表面缺乏特定的趋化因子受体,不能很快地向病变区域迁移,从而限制了它的疗效。
趋化因子是一组低分子量蛋白家族,在炎症病变区域大量表达,能够吸引白细胞向感染部位迁移,在炎症反应中有重要作用。李济元等[9]发现白藜芦醇可以抑制肺癌细胞A549的增殖及CXCR4在其中的表达。小鼠急性肺损伤的炎症区域发现大量表达趋化因子CXCL-1、CXCL-2、CXCL-12和CXCL-15[10],其中CXCL-12又称基质细胞衍生因子-1(stromal-derived factor-1,SDF-1)属于趋化因子蛋白家族,首先在淋巴组织和骨髓中被发现,对CXCR4受体介导的造血干细胞和淋巴细胞的归巢起至关重要的作用[11-12]。此后,SDF-1在更多组织中被发现,尤其是在急性肺损伤和肺纤维症的肺泡中高表达[13]。MSC的表面表达与SDF-1对应的受体CXCR4,然而,随着其在体外扩增该受体的表达逐渐减少,限制了细胞移植后的疗效[14-15]。
为了增强MSCs体内移植后的治疗效果,本研究采用已构建了的CXCR4过表达和低表达载体,经过转染使MSCs高表达或低表达趋化因子受体CXCR4,体外研究CXCR4高表达及低表达对MSCs增殖、分化、迁移能力的影响,为其体内移植和临床应用提供实验依据。
1 材料与方法
1.1 动物 清洁级Sprague-Dawley(SD)大鼠(4周龄,120~150 g)购自大连医科大学实验动物中心[SCXK(辽)2008-0002]。
1.2 试剂 CXCR4质粒、293T细胞(天津医科大学闫亚平教授惠赠),青-链霉素(penicillin-strepotomycin,P/S,Thermo公司),DMEM高糖培养基(Gibco公司),胎牛血清(FBS,Gibco公司),大鼠骨髓间充质干细胞专用胎牛血清(FBS,Cyagen公司),L-谷氨酰胺(L-Glutamine,Cyagen公司),0.25%胰蛋白酶(0.25% Trypsin,Cyagen公司),CD29-PE抗体、CD34-FITC抗体、CD44-FITC抗体、CD45-FITC抗体、CD90-FITC抗体(美国BD),一抗分别为兔抗CXCR4、vWF、SP-C(北京博奥森),二抗为Cy3标记驴抗兔IgG(Jackson)、FITC标记羊抗兔IgG(Jackson),结晶紫粉末(Amresco公司),大鼠重组SDF-1α(PeproTech公司)。
1.3 仪器 Ti-S型荧光显微镜(日本尼康),超低温冰箱(青岛海尔,DIV-86L386),CO2培养箱(NU-AIRE,BPN),流式细胞仪(美国BD公司),双人超净工作台(美国Agilent公司,ZHJH-C1112B),台式高速冷冻离心机(湖南湘仪,TGL-20M)。
1.4 实验分组 将细胞随机分为3组,每组3个复孔,重复3次。分别为:① GFP组;② CXCR4+组;③ CXCR4-组。
1.5 大鼠骨髓间充质干细胞(MSCs)的培养和鉴定 取120~150 g ♂ SD大鼠1只,10%乌拉坦麻醉10 min。75%酒精消毒肚皮,剪开腹股沟,剥离皮肤,分离两后肢长骨(股骨和胫骨),去除骨膜及残留肌肉,浸泡在含2% P/S的PBS缓冲液中,转移至超净台。减去长骨两端,暴露髓腔,用针头以无血清DMEM/LOW反复冲洗骨髓腔,过70 μm孔径的细胞筛网,收集滤液。离心1 000 r·min-1×6 min,弃上清。全骨髓贴壁分离法重悬细胞,计数,5×105个细胞接种于25 cm2培养瓶,镜下观察细胞情况,置37℃、5% CO2培养箱中培养。取P3代细胞,待细胞90%融合时进行鉴定。抗体标记,将单细胞悬液离心洗涤,重悬计数后进行抗体标记,所标记细胞每份不少于5×106个细胞,用CD29-PE、CD34-FITC、CD44-FITC、CD45-FITC、CD90-FITC,按1 μg抗体/106细胞分别标记细胞,将处理好的样本用流式细胞仪检测细胞表面CD29、CD34、CD44、CD45、CD90的表达[4-5,16-17]。
1.6 构建CXCR4高表达质粒、低表达质粒、转染MSCs和CXCR4表达的检测 ① GFP、CXCR4表达及CXCR4低表达质粒:由天津医科大学闫亚平教授前期构建提供。② CXCR4基因转染BM-MSCs及CXCR4表达的检测:转染293T包装细胞,对照组转染L.v.-GFP。培养293T细胞,24 h后荧光显微镜下观察两组细胞都呈现绿色荧光。分别收集72 h内上述293T细胞培养液上清(含病毒),离心、浓缩并检测病毒滴度;用浓缩的病毒上清转染第3代BM-MSCs;转染后3 d的GFP-MSCs和CXCR4+-MSCs、CXCR4--MSCs用4%多聚甲醛固定30 min,PBS洗涤3次,用0.1% Triton X-100透化20 min,5%山羊血清封闭1 h后,加入兔抗大鼠CXCR4一抗,4 ℃孵育过夜,次日用PBS洗涤3次,加入山羊抗兔Cy3荧光二抗,室温孵育1 h,用PBS洗涤3次,0.5 mg·L-1的DAPI室温孵育5 min后用抗荧光淬灭封片剂封片,荧光显微镜下观察摄片[18-19]。
1.7 CXCR4基因转染对MSCs增殖能力的影响检测 将基因转染5 d后的CXCR4-MSCs 和对照组 GFP-MSCs 以及未转染细胞Mock-MSCs分别以6×106个/孔接种于96孔板中,每隔2 d换液一次。每天取6个复孔的细胞,消化计数,计算平均值,连续观察9 d[20-21]。以细胞培养时间为横轴,细胞数为纵轴,绘制细胞生长曲线。
1.8 CXCR4基因转染对MSCs向肺组织分化能力的影响检测 脂多糖(LPS)诱导的肺组织损伤条件培养基(condition medium,CM)的制备:取120~150 g ♂ SD大鼠,10%乌拉坦麻醉,打开胸腔,心脏灌流至肺部发白后剪下肺叶。肺叶用眼科手术剪剪成约1 mm的组织块,经研磨过70 μm细胞筛网得到肺组织细胞,以5×107·L-1的密度接种于6孔板中,每孔2 mL含10%胎牛血清的完全培养基进行培养。接种24 h后全量换液,根据分组情况加入完全培养基或含10 mg·L-1脂多糖的完全培养基致炎。每组3个复孔,作用4 h后获得细胞上清液,3 000 r·min-1离心10 min去除细胞碎片,所得即LPS诱导的肺组织损伤的条件培养基[22],经ELISA检测CM中的IL-1β和TNF-α水平,与未加LPS处理组的肺组织上清液比较明显升高(数据未在文中显示),表明构建模型成功,-80℃保存待用。体外培养的肺组织细胞经LPS致炎所得的条件培养基(CM)孵育MSCs诱导其分化,24孔板内以5×104·L-1的密度接种MSC,接种24 h后将培养基内加入50% CM,每2 d换液一次,形成CM-MSCs共培养体系,8 d后用免疫荧光细胞化学法检测肺胞中SP-C(肺泡II型上皮细胞标记物)、vWF(肺血管内皮细胞标记物)特异性蛋白的表达。
1.9 免疫荧光细胞化学法检测 体外培养的MSCs和CM诱导8 d的MSC用4%多聚甲醛固定30 min,PBS洗涤3次,用0.1% Triton X-100透化20 min,5%山羊血清封闭1 h后,分别加入大鼠抗SP-C、vWF等一抗,4 ℃孵育过夜,次晨用PBS洗涤3次,分别加入与一抗对应的种属特异性Cy3、FITC荧光二抗,室温孵育1 h,用PBS洗涤3次,0.5 mg·L-1的DAPI室温孵育5 min后用抗荧光淬灭封片剂封片,荧光显微镜下观察摄片[20-21,23]。
1.10 CXCR4转基因MSCs的体外迁移实验 将膜孔孔径为8 μm的Transwell小室悬挂在24孔板中,将转染后5 d的CXCR4+-MSCs、CXCR4--MSCs、GFP-MSCs和未经转染的Mock-MSCs细胞悬液100 μL(含1×104个细胞)加入上室,将含不同浓度趋化因子SDF-1α(0、50、100 μg·L-1)的培养基600 μL加入下室,37 ℃、5% CO2孵育5 h。培养5 h后细胞穿过膜孔粘附在小室膜的下表面。迁移结束后取出小室稍稍水洗,用棉签擦去滤膜上表面未穿膜的细胞,4%多聚甲醛固定15 min,用0.5%结晶紫染色10 min,显微镜下计数10个不同视野的穿膜细胞数,取平均值[24-25]。
2.1 MSCs培养和形态观察与鉴定 在倒置显微镜下观察,72 h后可见贴壁细胞呈小圆形及多角形。培养至d 7,多数细胞伸展呈梭形并呈集落式生长,培养到d 10~14融合度达80%~90%。传代培养至第2代(P2)时细胞形态趋于统一,呈现为长梭形细胞束排列。经流式细胞仪检测MSCs表面抗原标记物CD29、CD44、CD90、CD34、CD45,经流式细胞仪鉴定,CD29(99.5%)、CD44(99.1%)、CD90(96.9%)阳性,CD34(0.31%)、CD45(0.14%)阴性,即CD29+/CD44+/CD90+/CD34-CD45-,表明所获得的细胞是MSCs,可用于后续实验(Fig 1)。
2.2 CXCR4基因转染MSCs和CXCR4表达的检测 用免疫细胞化学法检测,结果显示,转染了CXCR4-GFP共表达载体的CXCR4+-MSCs、CXCR4--MSCs和转染了原载体(含GFP)的对照组GFP-MSCs都明显表达 GFP(绿色),说明3组细胞都被病毒质粒有效转染;但CXCR4(红色)只在CXCR4+-MSCs中明显表达,而在CXCR4--MSCs及对照组GFP-MSCs中几乎不表达(Fig 2A)。对免疫荧光的结果进行拍照,对CXCR4阳性细胞进行计数,CXCR4+-MSCs组中CXCR4阳性细胞占90.2%,CXCR4--MSCs组中CXCR4阳性细胞占1.2%,而GFP-MSCs对照组中仅占7.6%,说明CXCR4+-MSCs能高表达CXCR4(Fig 2B,2C)。
2.3 CXCR4基因转染不影响MSCs的增殖能力 基因转染5 d后开始绘制CXCR4+-MSCs、对照组GFP-MSCs和未转基因的Mock-MSCs 3组细胞的生长曲线,结果表明3组细胞的扩增能力无明显差异(P>0.05);以上结果说明,CXCR4高表达不影响MSCs的增殖能力(Fig 3)。
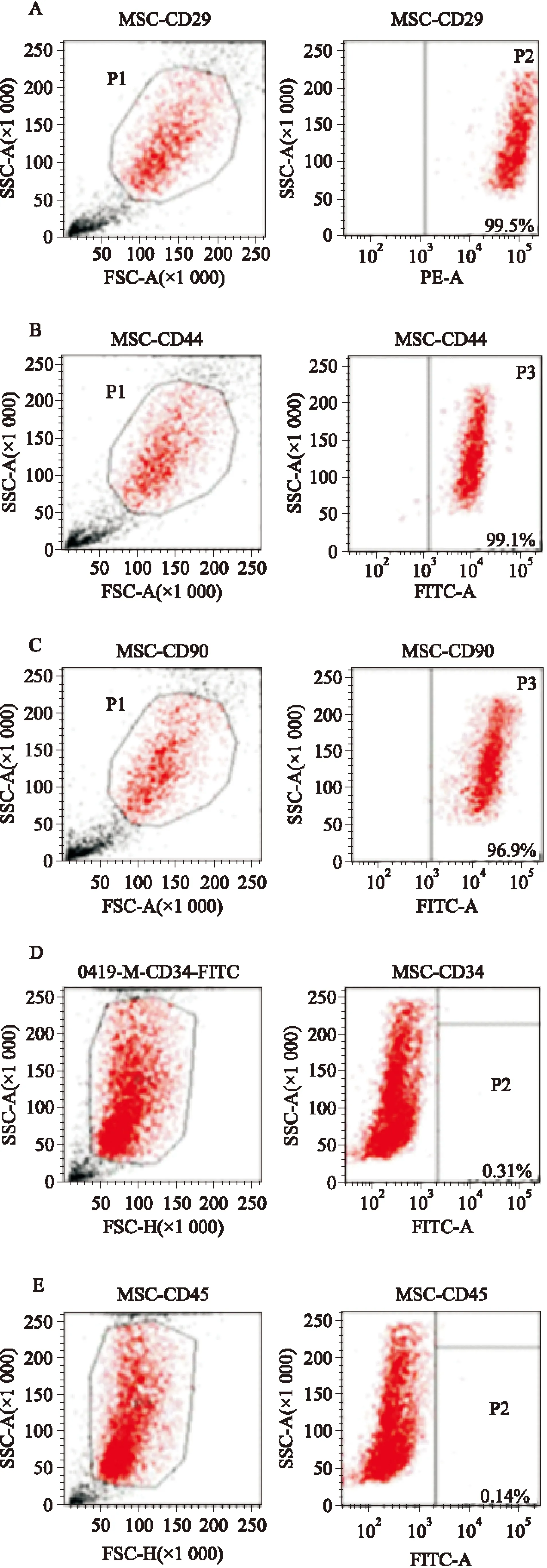
Fig 1 Immunophenotype of MSCs by using flow cytometry
A:CD29 positive;B:CD44 positive; C:CD90 positive;D:CD34 negative;E:CD45 negative
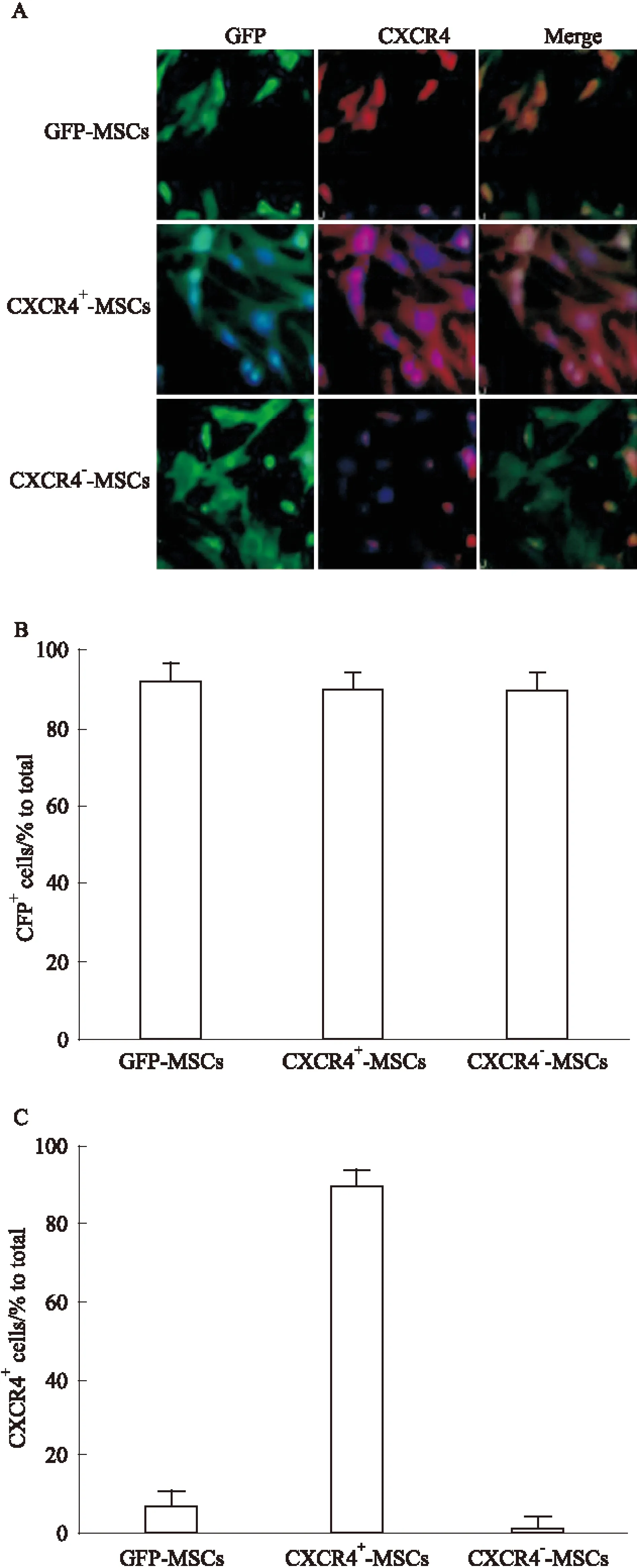
Fig 2 Identification of CXCR4 expression in CXCR4-transduced MSCs
A:Immunocytochemistry staining:GFP(green) expression in MSCs transduced with both vectors was visible by fluorescence microscopy 3 days after infection. Magnification×20.Green:GFP;Red:CXCR4; Blue: DAPI stained nuclei.B:GFP-transduced MSCs were quantitatively analyzed by cell counting. Data were compared with isotype-matched controls. One representative experiment of three is shown.C:CXCR4-transduced MSCs were quantitatively analyzed by cell counting. One representative experiment of three is shown.
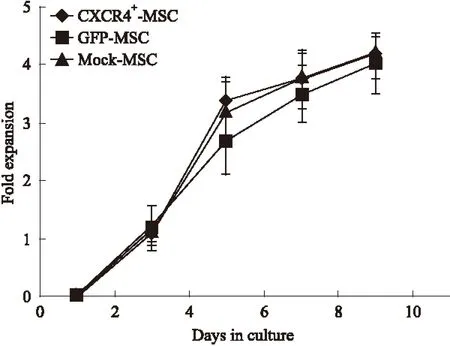
Fig 3 Assay for self-renewal capacity of CXCR4-transduced MSCs
Growth curve:CXCR4+-MSCs,GFP-MSCs or Mock-MSCs were seeded at the same density in 24 well plate 5 days post gene-transfection. At day 1, 3, 5, 7, 9; MSCs in each well were dissociated to single cells, numbers of the cells per well were counted by hemocytometer.A growth curve of MSCs revealed by enumerating the cells at each time point was developed.n=6.
2.4 CXCR4高表达、低表达对MSCs向肺组织细胞分化能力无影响 为了检测CXCR4基因转染是否影响MSCs的分化能力,体外培养的肺组织细胞经LPS致炎所得的条件培养基(CM)孵育MSCs诱导其分化,8 d后用免疫荧光细胞化学法对肺泡Ⅱ型上皮细胞标记蛋白SP-C(红色,Fig 4A)和血管内皮细胞标记蛋白vWF(红色,Fig 4B)进行染色,比较未转染的MSCs(Mock-MSCs)、高表达CXCR4基因的MSCs(CXCR4+-MSCs)和低表达CXCR4基因的MSCs(CXCR4--MSCs)组在CM诱导下的分化能力。定量分析结果表明,CXCR4基因转染对MSCs向肺组织分化的能力没有影响(P>0.05,Fig 4C)。
2.5 CXCR4高表达增强了MSCs的迁移能力,低表达不影响其迁移能力 为检测高表达CXCR4对MSCs迁移能力的影响,将CXCR4+-MSCs、CXCR4--MSCs、GFP-MSCs和未经转染的Mock-MSCs细胞置于Transwell小室上室,下室分别放置不同浓度的趋化因子SDF-1α(0、50、100 μg·L-1),孵育后取出小室用结晶紫染色并观察拍照(穿膜细胞为蓝紫色,Fig 5A),对穿膜细胞进行计数。迁移实验结果显示,CXCR4+-MSCs迁移细胞明显多于GFP-MSCs;定量分析结果显示,100 μg·L-1的SDF-1α对CXCR4+-MSCs的迁移效率作用明显,CXCR4+-MSCs的迁移能力明显高于GFP-MSCs(50、100 μg·L-1SDF-1α),定量分析结果显示,50 μg·L-1SDF-1α因子的趋化下CXCR4+-MSCs迁移数量为55.67±2.83,明显高于GFP-MSCs的迁移细胞数(30.34±4.11,P<0.01,Fig 5B),100 μg·L-1SDF-1α因子的趋化下CXCR4+-MSCs迁移数量为78.06±6.68,明显高于GFP-MSCs的迁移细胞数(40.67±2.87,P<0.01)。其中,CXCR4--MSCs组和GFP-MSCs组比较差异无统计学意义。以上迁移实验结果表明,CXCR4高表达促进MSCs的迁移能力,低表达不影响MSCs的迁移能力(Fig 5)。
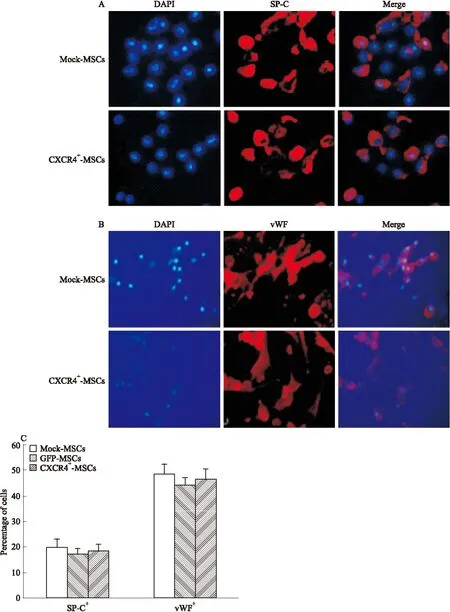
Fig 4 Assay for differentiation of CXCR4-transduced MSCs A and B:CM induced differentiation of Mock-MSCs, CXCR4+-MSCs was identified by immunostaining. Mock-or CXCR4-transduced MSCs were cultured in condition medium for 8 days, then stained with lung cell markers(red) and DAPI(blue). Examples show that CXCR4-MSCs differentiated into Alveolar epithelial cells(SP-C+), vascular endothelial(vWF+) as verified by immunostaining,Magnification×20.C:Percentages of MSCs differentiated into lung cells were calculated by the numbers of each individual cell type among total numbers of MSCs(DAPI+). There was no difference between the groups.
3 讨论
骨髓MSCs具有取材简单、体外扩增能力强等优势,具有很强的自我更新能力和多向分化潜能,是细胞治疗和基因工程的良好载体。MSCs本身通过旁分泌实现的免疫调节作用可以改善急性肺损伤等系统免疫疾病的症状,减轻全身炎症、缓解组织损伤,MSCs还可以迁移到病灶区域通过定植、分化在一定程度上实现组织再生[6,26-27]。
CXCR4广泛地表达于多种细胞和组织,包括免疫细胞、肺、心脏、肾、肝和脾[28]。趋化因子及其受体介导一系列白细胞应答,包括趋化现象和免疫激活[28]。在发生急性损伤的肺病变区域,由于剧烈的炎症反应产生高水平的趋化因子SDF-1α,与炎症细胞表面的趋化因子受体CXCR4结合,引起炎细胞的聚集与激活[29-30]。但随着MSCs的体外扩增,其表面趋化因子受体CXCR4逐渐降低,影响其移植后在体内向产生趋化因子的受损肺组织迁移的能力,从而限制了它的疗效[14-15]。使细胞表面过表达某种趋化因子受体是加快移植细胞迁移的有力方案,因此我们假设,将CXCR4基因转入MSCs,高表达CXCR4的MSCs应该能更快速地迁移到受损的肺组织,缩短起效时间从而增强疗效。
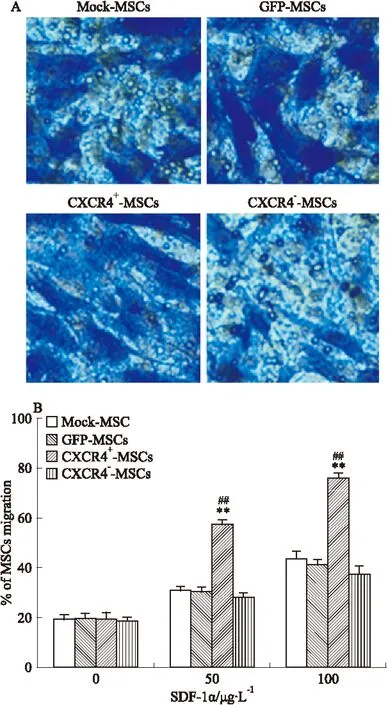
Fig 5 Chemotaxis assay for CXCR4-transduced MSCs

为了验证上述假设,我们首先体外扩增了纯度较高的骨髓MSCs,经流式细胞仪鉴定纯度在90%以上。为了增强MSCs的迁移能力,我们用已构建了的CXCR4-GFP载体质粒,并包装成慢病毒感染MSCs,转染3 d后经细胞免疫荧光化学检测,MSCs既表达GFP又表达CXCR4证明转染成功。体外增殖、分化实验表明,转染了GFP和CXCR4+-GFP、CXCR4--GFP的MSCs与未转染的细胞比较,增殖能力并未受到影响,分化为肺组织细胞的能力差异也无显著性,说明基因转染不影响MSCs的增殖和分化能力。体外趋化实验表明,高表达CXCR4的MSCs比只表达GFP的细胞对趋化因子SDF-1α有更强的趋化能力,表明CXCR4高表达能够使MSCs增强向SDF-1趋化的能力。
综上所述,CXCR4高表达能明显增强BM-MSCs的趋化能力,说明其植入体内后不仅可保持其原有的细胞替代与组织修复作用,而且可快速大量地向病灶部位的迁徙,明显增强治疗作用,是治疗肺损伤等疾病的理想种子细胞,具有很广阔的应用前景。
(致谢:本实验于辽宁中医药大学药学院药理实验室完成。在此向所有参与本实验的人员表示衷心感谢!)
[1] Keating A.Mesenchymal stromal cells: new directions[J].CellStemCell,2012,10(6): 709-16.
[2] Hayes M,Curley G,Laffey J G. Mesenchymal stem cells-a promising therapy for acute respiratory distress syndrome[J].F1000MedRep,2012,4:2.
[3] Sensebé L,Krampera M,Schrezenmeier H,et al. Mesenchymal stem cells for clinical application[J].VoxSang,2010,98(2):93-107.
[4] Griffin M,Iqbal S A,Bayat A. Exploring the application of mesenchymal stem cells in bone repair and regeneration[J].JBoneJointSurgBr,2011,93(4):427-34.
[5] Dominici M,Le Blanc K,Mueller I,et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement[J].Cytotherapy,2006,8(4):315-7.
[6] Gupta N,Su X,Popov B,et al. Intrapulmonary delivery of bone marrow-derived mesenchymal stem cells improves survival and attenuates endotoxin-induced acute lung injury in mice[J].JImmunol,2007,179(3):1855-63.
[7] Kotton D N,Ma B Y,Cardoso W V,et al. Bone marrow-derived cells as progenitors of lung alveolar epithelium[J].Development,2001,128(24):5181-8.
[8] Rojas M,Xu J,Woods C R,et al. Bone marrow-derived mesenchymal stem cells in repair of the injured lung[J].AmJRespirCellMolBiol,2005,33(2):145-52.
[9] 李济元,王卫华,张彦梅,等.白藜芦醇对肺腺癌细胞增殖和CXCR4表达的影响[J]. 中国药理学通报,2012,28(4):588-9.
[9] Li J Y,Wang W H, Zhang Y M,et al.Effects of resveratrol on proliferation and expression of CXCR4 in lung adenocarcinoma cells[J].ChinPharmacolBull,2012,28(4):588-9.
[10]Petty J M,Sueblinvong V,Lenox C C,et al. Pulmonary stromal-derived factor-1 expression and effect on neutrophil recruitment during acute lung injury[J].JImmunol,2007,178(12):8148-57.
[11]Bleul C C,Farzan M,Choe H,et al. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry[J].Nature,1996,382(6594):829-33.
[12]Oberlin E,Amara A,Bachelerie F,et al. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1[J].Nature,1996,382(6594):833-5.
[13]Phillips R J,Burdick M D,Hong K,et al. Circulating fibrocytes traffic to the lungs in response to CXCL12 and mediate fibrosis[J].JClinInvest,2004,114(3):438-46.
[14]Honczarenko M,Le Y,Swierkowski M,et al. Human bone marrow stromal cells express a distinct set of biologically functional chemokine receptors[J].StemCells,2006,24(4):1030-41.
[15]Wynn R F,Hart C A,Corradi-Perini C,et al. A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow[J].Blood,2004,104(9):2643-5.
[16]Liang Z X,Sun J P,Wang P,et al. Bone marrow-derived mesenchymal stem cells protect rats from endotoxin-induced acute lung injury[J].ChinMedJ(Engl),2011,124(17):2715-22.
[17]Curley G F,Hayes M,Ansari B,et al. Mesenchymal stem cells enhance recovery and repair following ventilator-induced lung injury in the rat[J].Thorax,2012,67(6):496-501.
[18]Yang J, Yan Y, Xia Y, Kang T, et al. Neurotrophin 3 transduction augments remyelinating and immunomodulatory capacity of neural stem cells[J].MolTher,2014, 22(2):440-50.
[19]Yang J,Jiang Z,Fitzgerald D C,et al. Adult neural stem cells expressing IL-10 confer potent immunomodulation and remyelination in experimental autoimmune encephalitis[J].JClinInvest,2009,119(12): 3678-91.
[20]Zhang N,Kang T,Xia Y,et al. Effects of salvianolic acid B on survival,self-renewal and neuronal differentiation of bone marrow derived neural stem cells[J].EurJPharmacol,2012, 697(1-3):32-9.
[21]Yang J,Yan Y,Ma C G,et al. Accelerated and enhanced effect of CCR5-transduced bone marrow neural stem cells on autoimmune encephalomyelitis[J].ActaNeuropathol,2012,124(4):491-503.[22]Zhao J, He D,Su Y,et al. Lysophosphatidic acid receptor 1 modulates lipopolysaccharide-induced inflammation in alveolar epithelial cells and murine lungs[J].AmJPhysiolLungCellMolPhysiol,2011,301(4):547-56.
[23]Yang J,iang Z,Fitzgerald D C,et al. Adult neural stem cells expressing IL-10 confer potent immunomodulation and remyelination in experimental autoimmune encephalitis[J].JClinInvest,2009,119(12):3678-91.
[24]Du Z,Wei C,Yan J,et al. Mesenchymal stem cells overexpressing C-X-C chemokine receptor type 4 improve early liver regeneration of small-for-size liver grafts[J].LiverTranspl,2013,19(2):215-25.
[25]张 悦,欧来良,程兆康,等. CXCR4基因修饰骨髓间充质干细胞体外迁移实验[J]. 生物医学工程学杂志,2009,26(3):595-600.
[25]Zhang Y,Ou L L,Cheng Z K,et al.CXCR4 gene modification between bone marrow mesenchymal stem cell migration in vitro experiment[J].JBiomedlEngin,2009,26(3):595-600.
[26]Zhu Y G,Feng X M,Abbott J,et al. Human Mesenchymal Stem Cell Microvesicles for Treatment of Escherichia coli Endotoxin-Induced Acute Lung Injury in Mice[J].StemCells,2014,32(1):116-25.
[27]Inamdar A C,Inamdar A A. Mesenchymal stem cell therapy in lung disorders: pathogenesis of lung diseases and mechanism of action of mesenchymal stem cell[J].ExpLungRes,2013,39(8):315-27.
[28]崔红霞,冯一中,顾振纶,等. CXCL12/CXCR4生物轴与特发性肺纤维化研究进展[J].中国药理学通报,2010,26(3):298-301.
[28]Cui H X,Feng Y Z,Gu Z L,et al.Research progress of CXCL12/CXCR4 biological axis and idiopathic pulmonary fibrosis[J].ChinPharmacolBull,2010,26(3):298-301.
[29]Yellowley C.CXCL12/CXCR4 signaling and other recruitment and homing pathways in fracture repair[J].BonekeyRep,2013,13(2):300.
[30]Bhatia M,Zemans R L,Jeyaseelan S. Role of chemokines in the pathogenesis of acute lung injury[J].AmJRespirCellMolBiol,2012,46(5):566-72.
Effects of CXCR4 gene transfection on biological behavior of bone marrow mesenchymal stem cells (invitro)
WANG Yu-ying1,ZHANG Nan1,LI Xiu-li1,WANG Ya-meng1,LI Shao-heng1,YAN Yu-hui1,SONG Jie1,YANG Jing-xian1,WEN Qing-ping2
(1.DeptofPharmacology,SchoolofPharmacy,LiaoningUniversityofTraditionalChineseMedicine,DalianLiaoning116600,China; 2.DalianMedicalUniversity,DalianLiaoning116044,China)
Aim To observate the effect of chemokine receptor(CXCR4) gene transfection on biological behavior of bone marrow mesenchymal stem cellsinvitro.Methods Firstly, bone marrow mesenchymal stem cells were divided into three groups:GFP(transfected GFP into MSCs), CXCR4+(transfected CXCR4+into MSCs) and CXCR4-(transfected CXCR4-into MSCs) group.Then, their capacity of proliferation, differentiation and migration ability (invitro) was assessed with immunofluorescence cytochemistry method, flow cytometry assay and Transwell cell chemotaxis test.Results The high or low expression of CXCR4 had no effect on their ability of proliferation and differentiation into lung tissue. Compared with GFP group, however, CXCR4+-MSCs group significantly increased the number of migrating cells, while CXCR4--MSCs group showed no change in the number of migrating cells.Conclusions The proliferation and differentiation capacities are not affected by the high or low expression of CXCR4. The high expression of CXCR4 can significantly enhance the migration ability of MSCs to inflammatory lesions, and the low one has no effect on the migration of the cells. After the transplantation of MSCs, CXCR4′s high expression will access to the lesion area to participate in tissue repairing rapidly and largely, significantly enhancing the therapeutic efficacy.
bone marrow mesenchymal stem cells; chemokine receptor CXCR4; gene transfection; proliferation; differentiation; Migration
时间:2017-5-25 17:44 网络出版地址:http://kns.cnki.net/kcms/detail/34.1086.R.20170525.1744.026.html
2017-01-22,
2017-03-30
国家自然科学基金资助项目(No 81273923)
王玉莹(1992-),女,硕士生,研究方向:神经药理学,E-mail:799363692@qq.com; 杨静娴(1963-),女,博士,教授,博士生导师,研究方向:神经药理学,通讯作者,E-mail:jingxianyang@yahoo.com; 闻庆平(1971-),男,博士,教授,博士生导师,研究方向:心血管麻醉和危重症麻醉,通讯作者,E-mail: wenqp126@126.com
10.3969/j.issn.1001-1978.2017.06.013
A
1001-1978(2017)06-0806-08
R322.35;R329.24;R392.11;R394.2;R563.8;R977.6
猜你喜欢
生物化工(2022年4期)2022-09-20
中风与神经疾病杂志(2021年9期)2021-11-08
昆明医科大学学报(2021年3期)2021-07-22
昆明医科大学学报(2021年5期)2021-07-22
现代临床医学(2021年2期)2021-03-29
世界科学技术-中医药现代化(2021年10期)2021-03-02
中华养生保健(2020年8期)2021-01-14
世界最新医学信息文摘(2021年55期)2021-01-07
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13