
脂联素对低氧条件下大鼠肺微动脉内皮细胞NO生成的促进作用及其机制
2017-06-07 08:23徐海军张存娟周宁娟第四军医大学西京医院儿科西安700杨凌示范区医院儿科第四军医大学西京医院老年病科通讯作者mailsunxin6fmmueducn
山西医科大学学报 2017年5期
徐海军,张存娟,周宁娟,苏 慧,孙 新(第四军医大学西京医院儿科,西安 700;杨凌示范区医院儿科;第四军医大学西京医院老年病科;通讯作者,E-mail:sunxin6@fmmu.edu.cn)
脂联素对低氧条件下大鼠肺微动脉内皮细胞NO生成的促进作用及其机制
徐海军1,2,张存娟1,周宁娟1,苏 慧3,孙 新1*
(1第四军医大学西京医院儿科,西安 710032;2杨凌示范区医院儿科;3第四军医大学西京医院老年病科;*通讯作者,E-mail:sunxin6@fmmu.edu.cn)
目的 重组人球状脂联素(APN)促进低氧条件下大鼠肺微血管内皮细胞(PMVECs)NO的生成,探讨其潜在的分子机制研究。 方法 原代培养SD大鼠PMVECs,传至第3代经免疫组化法鉴定细胞传至第3代鉴定细胞;PMVECs分4组,常氧组(210 ml/L O2,37 ℃);低氧组(20 ml/L O2);低氧+APN组(20 ml/L O2+APN 1 μg/ml),低氧+APN+L-NAME组(20 ml/L O2,+APN 1 μg/ml+L-NAME 1 μg/ml)处理,各组细胞同时处理(加药)后,培养12 h收集上清,硝酸还原法测NO浓度,RT-PCR测定eNOS mRNA的基因表达Western blot检测AMPK/p-AMPK、PI3K/p-PI3K、Akt/p-Akt、eNOS/p-eNOS蛋白表达。 结果 鉴定细胞为PMVECs。与常氧组比较,低氧组NO浓度、eNOS mRNA表达水平显著下降(P<0.01);与低氧组比较,低氧+APN组低氧诱导的NO浓度、eNOS mRNA表达水平显著增加(P<0.01);L-NAME可阻断NO的产生和eNOS mRNA的表达。各组AMPK、PI3K、Akt、eNOS总蛋白表达量无差异;与常氧组比较,低氧组中AMPK、PI3K、Akt、eNOS磷酸化表达水平下降(P<0.05);与低氧组比较,低氧+APN组AMPK、PI3K、Akt、eNOS磷酸化表达水平增加(P<0.05);与低氧+APN组比较,低氧+APN+L-NAME组AMPK、PI3K、Akt磷酸化表达水平没有变化(P>0.05);L-NAME可阻断eNOS磷酸化的表达(P<0.01)。 结论 低氧条件下APN可促进PMVECs生成NO,其可能机制是AMPK/PI3K/Akt/eNOS/NO信号通路的激活。
脂联素; 肺微动脉内皮细胞; 信号通路; 一氧化氮
肺动脉高压(pulmonary arterial hypertension,PAH)是起源于肺动脉、最终影响心脏的微小血管疾病[1],主要表现为肺动脉阻力持续性升高、平滑肌增生、原位血栓形成,损害肺细小动脉和肺动脉压力持续上升的恶性肺血管疾病[2],最终死于右心室容量超负荷导致的右心衰竭。低氧性肺动脉高压(hypoxia pulmonary hypertension,HPH)是PAH中最常见类型,低氧引起PMVECs的功能障碍,主要表现为PMVECs生成一氧化氮(NO)减少,后者目前被认为是最重要的舒血管因子[1]。所以,增加PMVECs NO的生成,促进和改善低氧诱导PMVECs功能障碍是治疗低氧性肺动脉高压的关键。
脂联素(APN)是脂肪细胞分泌的一种重要的脂肪因子,研究已经证实,低脂联素血症是心血管疾病的一个独立的危险因素,动物研究表明增加APN浓度能够改善肥胖诱导的内皮细胞功能障碍,降低血压[3]。APN能够改善血管内皮细胞eNOS功能障碍和提高血管对NO的生物利用度[4]。
目前,APN研究的重点主要集中在高血压和心肌疾病上,是否对肺动脉高压有作用还不清楚。本课题组前期通过SD大鼠低氧性肺动脉高压模型,证实了APN对大鼠肺动脉PMVECs具有独立的保护作用[5]。本实验在细胞水平上进一步研究APN通过促进低氧诱导的PMVECs NO生成,降低血管紧张度,达到降低肺动脉高压的作用,并探讨其潜在的分子机制。
1 材料和方法
1.1 实验动物及主要试剂
SPF级6-8周龄SD雄性大鼠,生产许可证编号:SCXK(军)2012-0007,体质量150-180 g,由解放军第四军医大学实验动物中心提供。澳洲胎牛血清(赛奥美细胞技术有限公司,血清产地:澳大利亚),DMEM高糖培养基(美国Gibcro公司),内皮细胞生长因子(50×)(美国Gibcro公司),10%胰蛋白酶(美国Gibcro公司),APN(Recombinant Human gAcrp30/Adipolean)(美国Peprotech公司),CD31抗体(美国Abcam公司),FITC标记的IgG(美国Abbkine公司),DAPI(上海碧云天生物技术研究所),NO检测试剂盒(南京建成生物工程研究所),PVDF膜(美国Biosharp公司),p-AMPK/AMPK抗体、p-PI3K/PI3K抗体、p-Akt/Akt抗体(美国Cell Sinaling 公司),p-eNOS/eNOS抗体(美国BD公司),辣根过氧化物酶(HRP)标记羊抗兔IgG(武汉博士德生物工程有限公司)。
1.2 SD大鼠PMVECs的鉴定和实验分组
取大鼠肺边缘剪成1 mm×1 mm小块组织,制备PMVECs单细胞悬液,原代培养至第3代,玻片上细胞汇合至70%以上进行鉴定。免疫组化检测PMVECs表面标志物(CD31)抗原。PMVECs经CD31抗体(一抗),4 ℃孵化过夜,异硫酸荧光标记植物凝集素(FITC)标记的羊抗兔IgG抗体(二抗)孵育,DAPI核复染,免疫荧光显微镜下鉴定。将第3代PMVECs经10%胰蛋白酶消化后的细胞悬液,细胞浓度调整至(2.5-5.0)×105/ml,以5.0×104/cm2密度接种至4个培养瓶内继续培养,待铺满>60%瓶底时,分为常氧组(210 ml/L O2,37 ℃);低氧组(20 ml/L O2);低氧+APN(1 μg/ml)组,低氧+APN(1 μg/ml)+L-NAME(1 μg/ml)组处理,同时培养12 h。
1.3 各组细胞培养液上清NO测定
将试剂和待检测样品按照NO试剂盒说明书要求分别加入各管混匀,静置10 min,波长550 min,光径0.5 cm,双蒸水调零,测各管OD值,根据公式计算各组细胞上清NO浓度。
1.4 RT-PCR测定eNOS mRNA基因表达
弃去上清的PMVECs瓶内加入1 ml Trizol提细胞总RNA,RT-PCR法检测eNOS mRNA表达,以β-actin作为内参。设计引物为:Forward:5′-CAAGACCGATTACACGACATTGAGA-3′;Reverse:5′-TGAGGACTTGTCCAAACACTCCAC-3′。反转录:37 ℃,15 min;85 ℃,5 s;PCR扩增条件:95 ℃,30 s;变性95 ℃,5 s;退火60 ℃,30 s;共40个循环。以相对数形式表示(2-ΔΔCt),ΔCt=Ct目的基因-Ctβ-actin;ΔΔCt=ΔCt处理组-ΔCt对照组。
1.5 PMVECs的AMPK/p-AMPK、PI3K/p-PI3K、Akt/p-Akt、eNOS/p-eNOS蛋白表达的Western blot检测
弃去上清的PMVECs加入裂解液浓度为1 mmol/L,冰上使细胞充分裂解,采用BCA法测蛋白质浓度,取等量蛋白加入5×SDS-PAGE蛋白上样缓冲液混合并煮沸10 min,SDS-PAGE电泳,转膜PVDF后分别加入一抗p-AMPK/AMPK、p-PI3K/PI3K、p-Akt/Akt、p-eNOS/eNOS单克隆抗体,浓度均为1 ∶1 000,4 ℃过夜,羊抗兔IgG抗体(二抗)孵育,清洗3遍,经凝胶成像系统曝光Western blot条带,Image J软件计算和分析各组目的条带灰度值比值。
1.6 统计学分析
2 结果
2.1 PMVECs鉴定
倒置相差显微镜下PMVECs细胞边界清楚,呈典型的“鹅卵石” 样形态或“铺路石”样排列,胞质丰富;CD31表达呈阳性,细胞膜表面发红荧光,经DAPI复染细胞核发蓝色荧光(见图1)。
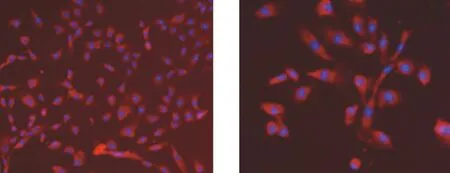
A.×200 B.×400图1 PMVECs免疫组织化学鉴定Figure 1 Identification of PMVECs by immunohistochemical method
2.2 培养液上清NO的测定
与常氧组比较,低氧组NO浓度显著下降(P<0.01);与低氧组比较,低氧+APN组NO浓度显著增加(P<0.01);与常氧组、低氧组、低氧+APN组比较,低氧+APN+L-NAME组NO浓度显著下降(P<0.01,见图2)。
2.3 eNOS mRNA基因的的表达水平
与常氧组比较,低氧组eNOS mRNA基因表达水平显著下降(P<0.01);与低氧组比较,低氧+APN组eNOS mRNA基因表达水平显著增加(P<0.01);与常氧组、低氧组、低氧+APN组比较,低氧+APN+L-NAME组eNOS mRNA基因表达水平显著下降(P<0.01,见图3)。
2.4 AMPK/p-AMPK、PI3K/p-PI3K、Akt/p-Akt、eNOS/p-eNOS的的表达水平水平
四组AMPK、PI3K、Akt、eNOS总蛋白表达不变;与常氧组比较,低氧诱导的AMPK、PI3K、Akt、eNOS磷酸化表达水平下降(P<0.05);与低氧组比较,低氧+APN组增加低氧诱导的AMPK、PI3K、Akt、eNOS磷酸化表达水平(P<0.05);L-NAME可阻断eNOS磷酸化的表达(见图4)。

与常氧组比较,*P<0.01;与低氧组比较,#P<0.01;与其他三组比较,△P<0.01图2 各组细胞培养液上清NO浓度检测(n=7)Figure 2 Concentration of NO in supernatant determined by nitrate reductive enzymatic method(n=7)

与常氧组比较,*P<0.01;与低氧组比较,#P<0.01;与其他三组比较,△P<0.01图3 用RT-PCR测定各组细胞eNOS mRNA基因相对表达量(n=7)Figure 3 Expression of eNOS mRNA by RT-PCR(n=7)
3 讨论
目前认为[7]早期低氧性肺动脉收缩反应(HPV)和后期合并肺动脉结构重建(PVR)是HPH最主要的两个发病环节。长期低氧PMVECs功能障碍,最主要表现NO生成减少引起血管舒缩失衡。因此,从源头上保护血管内皮细胞,增加NO生成;延缓肺动脉血管的重建成为治疗HPH的关键。APN是一种内源性胰岛素增敏剂[8],具有保护血管内皮细胞、调节代谢、 抗炎及保护心肌细胞等多种生物学作用,成为国内外研究的热点。苗兰等[9]在2%O2条件下,大鼠PMVECs经体外培养12 h后,细胞存活率降低,生化指标改变,细胞骨架基本完整,PMVECs功能障碍。本实验在低氧条件下诱导内皮细胞生成NO浓度,给予APN后NO明显增加;在细胞水平上证实了APN可能通过APMK/PI3K/Akt/eNOS/NO通路的激活发挥对肺动脉内皮细胞的作用,达到治疗肺动脉高压的作用。
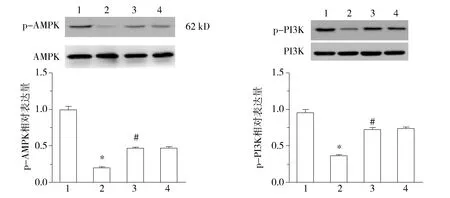

1.常氧组; 2.低氧组; 3.低氧+APN组; 4.低氧+APN+L-NAME组;与常氧组比较,*P<0.05;与低氧组比较,#P<0.05;与低氧+APN组比较,△△P<0.01图4 各组细胞总AMPK、Akt、PI3K、eNOS水平和磷酸化表达的表达的水平 (n=7)Figure 4 Protein levels of AMPK/p-AMPK,PI3K/p-PI3K,Akt/p-Akt and eNOS/p-eNOS detected by Western blot (n=7)
Kajimoto等[10]认为,内皮细胞呈现典型的“鹅卵石”样形态,内皮细胞膜表面特异性CD31高表达。我们的实验显示所培养的细胞为PMVECs呈典型的“鹅卵石”样,细胞膜表面特异性CD31高表达。内皮细胞合成和分泌多种生物活性物质,包括细胞因子、趋化因子、生长因子等调节血管稳态[11]。内皮细胞通过L-Arg-NO途径由一氧化氮合酶(NOS)催化L-精氨酸生成NO[12],NO单纯扩散进入血管平滑肌,平滑肌舒张,紧张度下降使血管舒张。研究[13]表明,低氧减少内皮细胞依赖的eNOS基因表达和诱发内皮细胞功能障碍,最主要表现为NO生成减少[14]和生物活性下降[15]导致的内皮细胞依赖的平滑肌舒张降低。我们的实验表明,与常氧组比较,低氧组NO浓度显著下降(P<0.01),eNOS mRNA基因表达水平显著下降(P<0.01),表明低氧诱导内皮细胞功能障碍;与低氧组比较,低氧+APN组NO浓度显著增加(P<0.01),eNOS mRNA基因表达水平显著增加(P<0.01),表明APN能够改善低氧诱导内皮细胞功能障碍,增加NO生成;与常氧、低氧、低氧+APN组比较,低氧+APN+L-NAME组NO浓度显著下降(P<0.01),eNOS mRNA基因表达水平显著下降(P<0.01),APN改善低氧诱导内皮细胞功能障碍可被 eNOS 抑制剂 L-NAME 阻断。Chen等[16]认为APN通过AMPK通路激活eNOS,使其磷酸化,增加NO舒张血管。 Western blot结果显示,各组AMPK、PI3K、Akt、eNOS总蛋白表达不变;与常氧组比较,低氧组AMPK、PI3K、Akt、eNOS磷酸化水平下降(P<0.05);与低氧组比较,低氧+APN组AMPK、PI3K、Akt、eNOS磷酸化水平增加(P<0.05);与低氧+APN组比较,低氧+APN+L-NAME组AMPK、PI3K、Akt磷酸化水平没有变化(P>0.05);L-NAME可阻断eNOS磷酸化水平(P<0.01),说明APN对低氧诱导的内皮细胞保护作用可能是通过AMPK/PI3K/Akt/eNOS/NO通路实现。
生理情况下,内皮细胞通过AMPK/PI3K/Akt/eNOS通路生成NO维持血管的舒张作用;低氧可诱导PMVECs功能障碍,主要表现为NO生成减少,血管舒张功能降低,紧张度增加形成肺动脉高压。本实验证实了,APN能够通过AMPK/PI3K/Akt/eNOS/NO通路纠正低氧诱导PMVECs功能障碍,增加eNOS mRNA基因表达和NO生成,血管舒张功能,降低肺动脉高压。
本研究表明,本研究结果表明,在细胞水平上,APN具有独立地降低低氧性肺动脉高压的作用,其可能的机制为AMPK/PI3K/Akt/eNOS/NO通路的激活,APN可能成为潜在用于治疗低氧性肺动脉高压的辅助药物。
[1] Cheng KK,Lam KS,Wang Y,etal.Adiponectin-induced endothelial nitric oxide synthase activation and nitric oxide production are mediated by APPL1 in endothelial cells[J].Diabetes,2007,56(5):1387-1394.
[2] Gladwin MT.Cardiovascular complications and risk of death in sickle-cell disease[J].Lancet,2016,387(10037):2565-2574.
[3] Ebrahimi-Mamaeghani M,Mohammadi S,Arefhosseini SR,etal.Adiponectin as a potential biomarker of vascular disease[J].Vasc Health Risk Manag,2015,11:55-70.
[4] Nosalski R,Guzik TJ.Perivascular adipose tissue inflammation in vascular disease[J].Br J Pharmacol,2017:Epub ahead of print.
[5] 张存娟,杨瑞,邢文娟,等.脂联素改善间歇性低氧性肺动脉高压大鼠离体肺动脉舒张功能及其机制[J].心脏杂志,2016,28(3):273-278.
[6] Hoeper MM,Humbert M,Souza R,etal.A global view of pulmonary hypertension[J].Lancet Respir Med,2016,4(4):306-322.
[7] Ma C,Li Y,Ma J,etal.Key role of 15-lipoxygenase/15-hydroxyeicosatetraenoic acid in pulmonary vascular remodeling and vascular angiogenesis associated with hypoxic pulmonary hypertension[J].Hypertension,2011,58(4):679-688.
[8] Markaki A,Psylinakis E,Spyridaki A.Adiponectin and end-stage renal disease[J].Hormones (Athens),2016,15(3):345-354.
[9] 苗兰英,林大勇,白剑,等.低氧条件下大鼠血管内皮细胞的形态观察[J].生物学杂志,2011,28(4):18-20.
[10] Kajimoto K,Hossen MN,Hida K,etal.Isolation and culture of microvascular endothelial cells from murine inguinal and epididymal adipose tissues[J].J Immunol Methods,2010,357(1-2):43-50.
[11] Rafii S,Butler JM,Ding BS.Angiocrine functions of organ-specific endothelial cells[J].Nature,2016,,529(7586):316-325.
[12] Shan L,Wang B,Gao G,etal.L-Arginine supplementation improves antioxidant defenses through L-arginine/nitric oxide pathways in exercised rats[J].J Appl Physiol (1985),2013,115(8):1146-1155.
[13] Fish JE,Yan MS,Matouk CC,etal.Hypoxic repression of endothelial nitric-oxide synthase transcription is coupled with eviction of promoter histones[J].J Biol Chem,2010,285(2):810-826.
[14] Giuliano F.New horizons in erectile and endothelial dysfunction research and therapies[J].Int J Impot Res,2008,20(Suppl 2):S2-S8.
[15] Chalupsky K,Kracun D,Kanchev I,etal.Folic acid promotes recycling of tetrahydrobiopterin and protects against hypoxia-induced pulmonary hypertension by recoupling endothelial nitric oxide synthase[J].Antioxid Redox Signal,2015,23(14):1076-1091.
[16] Chen H,Montagnani M,Funahashi T,etal.Adiponectin stimulates production of nitric oxide in vascular endothelial cells[J].J Biol Chem,2003,278(45):45021-45026.
Effect of diponectinon on hypoxia-induced pulmonary microvascular endothelial cells and its mechanism
XU Haijun1,2,ZHANG Cunjuan1,ZHOU Ningjuan1,SU Hui3,SUN Xin1*
(1DepartmentofPediatrics,XijingHospital,FourthMilitaryMedicalUniversity,Xi’an710032,China;2DepartmentofPediatrics,HospitalofYanglingDemonstrationDistrict;3DepartmentofGeriatrics,XijingHospital,FourthMilitaryMedicalUniversity;*Correspondingauthor,E-mail:sunxin6@fmmu.edu.cn)
ObjectiveTo investigate the production of NO in rat pulmonary microvascular endothelial cells(PMVECs) under hypoxia,and to explore the underlying molecular mechanisms.MethodsThe third passage of primary culture PMVECs from SD rat were identified by immunohistochemical method and divided into four groups.The PMVECs in four groups were cultured respectively under normoxia condition(210 ml/L O2,37 ℃),hypoxia condition(20 ml/L O2),hypoxia condition(20 ml/L O2)+APN (1 μg/ml),hypoxia condition(20 ml/L O2)+APN (1 μg/ml)+L-NAME (1 μg/ml).Supernatant and cell lysate were collected after culture for 12 h.The concentration of NO in supernatant was determined by nitrate reductive enzymatic method,the expression of eNOS mRNA was detected by RT-PCR,and protein levels of AMPK/p-AMPK,PI3K/p-PI3K,Akt/p-Akt and eNOS/p-eNOS were detected by Western blot in each group.ResultsPrimary culture PMVECs were successfully identified.NO production and expression of eNOS mRNA in hypoxia group decreased significantly compared with normoxia group(P<0.01).NO production and eNOS mRNA expression in hypoxia+ANP group were significantly higher than those in hypoxia group(P<0.01).L-NAME deceased the production of NO and the expression of eNOS mRNA(P<0.01).There was no significant difference in total protein levels of AMPK,PI3K,Akt and eNOS among the four groups.Phosphorylated AMPK,PI3K,Akt and eNOS decreased significantly in hypoxia group compared with normoxia group(P<0.05).Phosphorylated AMPK,PI3K,Akt and eNOS increased significantly in hypoxia+ANP group compared with hypoxia group(P<0.05).Phosphorylated AMPK,PI3K and Akt showed no significant difference between hypoxia+ANP+L-NAME group and hypoxia+ANP group(P<0.05).L-NAME deceased the expression of eNOS phosphorylation(P<0.01).ConclusionAPN can promote the production of NO from PMVECs under hypoxic conditions at the cellular level by the activation of AMPK/PI3K/Akt/eNOS/NO signaling pathway.
adiponectin; pulmonary artery endothelial cell; signaling pathway; nitric oxide
国家自然基金资助项目(81270330,30700345,31371151,31271219);陕西省科技厅国际合作课题资助项目(2013KW30-02);陕西省科学技术研究发展计划资助项目(2013Kkjxx-89)
徐海军,男,1980-05生,硕士,E-mail:navy800628@163.com
2017-02-10
R544.1
A
1007-6611(2017)05-0415-05
10.13753/j.issn.1007-6611.2017.05.003
猜你喜欢
实用心脑肺血管病杂志(2021年12期)2021-12-08
天津医科大学学报(2021年3期)2021-07-21
陕西医学杂志(2021年6期)2021-06-18
天津医科大学学报(2019年6期)2019-08-13
国际呼吸杂志(2019年2期)2019-02-14
分析化学(2017年12期)2017-12-25
中国医药生物技术(2016年5期)2016-11-09
国外医药(抗生素分册)(2016年5期)2016-07-12
安徽医科大学学报(2015年9期)2015-12-16
中国病理生理杂志(2015年10期)2015-01-26
