
含碎石土壤酶活性对植被建植模式的响应
2017-05-23 11:17薛海龙许文年刘大翔
江苏农业科学 2017年7期
薛海龙+许文年+刘大翔


摘要:以含碎石土壤为研究对象,对不同建植模式土壤蔗糖酶、脲酶、碱性磷酸酶活性的变化进行研究,探讨不同植被恢复模式对土壤酶活性的影响机制。结果表明,三大类14种建植模式中,除了单植草本A2模式外,其余13种建植模式的土壤容重都有一定程度的增大,其中单植灌木模式下容重增大幅度最为明显;土壤含水率变化均明显;总孔隙度在三大类14种建植模式中出现降低现象,其中单植灌木模式下降低幅度明显且最大,草灌混植模式次之,单植草本模式没有明显变化;pH值没有明显变化。不同建植模式下土壤酶活性的垂直分布均表现为0~10 cm土层高于 10~20 cm土層;不同建植模式间土壤酶活性亦存在明显差异,其中以蔗糖酶活性差异最为显著,在B1模式(多花木兰栽植密度20%)下酶活性最高,A6模式(狗牙根栽植密度为100%)下最低,C模式(狗牙根多花木兰混植)下较为稳定;磷酸酶活性在C3模式(狗牙根栽植密度60%+多花木兰栽植密度40%)下最高;脲酶活性变化范围较小,且在整个土壤剖面中受建植模式影响不明显。相关分析表明,三大类建植模式下,土壤酶活性之间存在正相关关系,且土壤基本理化性质与土壤酶活性间也存在着一定相关性。
关键词:含碎石土壤;建植模式;酶活性;相关性分析
中图分类号: X171.4文献标志码: A文章编号:1002-1302(2017)07-0248-04
含碎石土壤是理化性质恶劣、生物生境丧失的一种特殊立地类型,也是植被恢复的困难立地类型之一[1]。含碎石土壤形成的原因多种多样:采矿过程中产生的尾矿废弃物、开山过程中产生的碎石废料、岩石风化、剥落等情况都能形成含碎石废弃地,不仅占用了大量土地资源,而且水土流失状况较为严重,影响了当地的生态环境[2-3]。有研究表明,在含碎石土壤中引入耐贫瘠、具有固氮作用的植物种类,形成稳定的人工林群落,可以形成相对适宜的群落环境,为更多植物种类的进入创造条件,使含碎石土壤中植被的物种多样性得以提高[4-6]。在含碎石土壤废弃地引种耐贫瘠植物,能够在一定程度上提高土壤质量,进而推动重建生态系统的恢复演替[7-8]。还有研究表明,植被混合型建植模式下,植被多样型建植模式恢复效果要高于植被单一型建植模式[9-10]。
然而,此前对于不同建植模式产生的效果差异研究仍然不够深入,本研究根据植物与土壤互作原理,认为土壤质量改善是植被立地的根本,因此以土壤质量为测定指标。为了研究不同建植模式对土壤质量的改良效果,以含碎石土壤为研究对象,以草灌植物搭配反映建植模式,以土壤酶活性反映土壤质量状况。其中土壤酶是土壤重要的组成成分之一,数量少但作用大,是土壤中的生物催化剂,在土壤营养物质循环和能量转化过程中起着重要的作用,与土壤肥力具有非常密切的联系,因此其活性可以反映土壤生物化学过程的强度和方向。本研究通过聚氯乙烯(PVC)管植物种植试验,探讨不同建植模式对土壤酶活性的影响,为含碎石土壤生态环境改良与恢复提供依据,本研究也对类似土壤的植被恢复具有积极作用。
1材料与方法
1.1试验材料
1.1.1含碎石土壤用直径16 cm、高20 cm(0~10 cm、10~20 cm 2层式设置,由胶带黏合密封)的PVC管装填含碎石土壤。碎石选取的是粒径≤1 cm的石灰岩颗粒,土壤则选择的是过1 mm筛的黄棕壤,碎石与土壤质量配比为 30 ∶100,装填前先将碎石与土壤按比例充分掺混均匀,运用分层和少量多次的方式装填含碎石土壤,装满至PVC管口为止,即形成20 cm种植土柱。
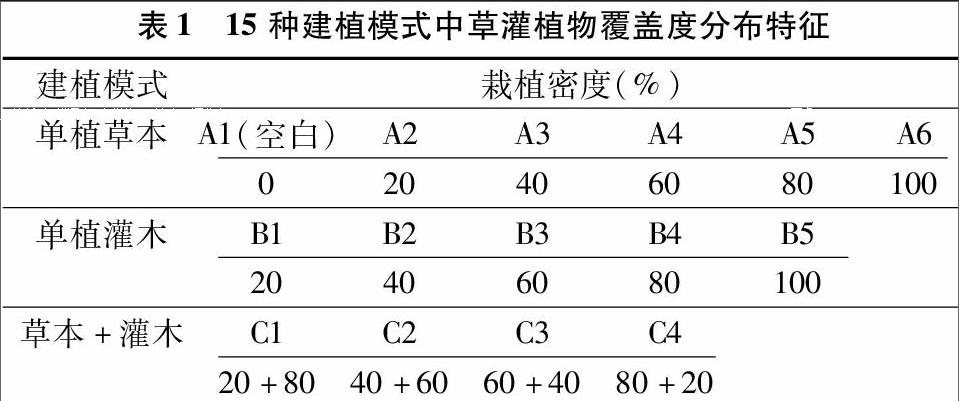
1.1.2植被为了实现快速建植,缩短研究周期,本试验选取2种耐性较强且生长期相对较短的常用先锋植物(草本植物:狗牙根,灌木植物:多花木兰),设置3类15种建植模式,具体设置见表1。每种模式设置3个平行处理,每个处理均依据表1中所设置的盖度比例移栽事先培育的相同生长时期(幼苗期)的植物幼苗,植株比例与植物数量相对应(例如:当草本+灌木=20%+80%时,即代表当下分别移栽了2株狗牙根和8株多花木兰幼苗)。
1.2样品采集方法与分析
采集植被生长4个月后的土壤进行试验。采集土样前,先将土柱表面干土与凋落物轻轻铲除,表面要铲平,尽量不要给下部土壤施加压力,以免土壤被压实。首先用10 cm口径环刀分层(0~10 cm、10~20 cm)取土,以测定土壤容重、含水量、总孔隙度以及pH值4个指标。然后再采用分层多点取样法,取20 cm(0~10 cm、10~20 cm)土壤,每层采样量1 kg左右,采样时将土钻垂直压入PVC管中,至钻头深度达到刻线20 cm处时停止,这时土钻中充满20 cm的土壤。将20 cm土壤分2次(0~10 cm、10~20 cm)取出,分别装入不同布袋或塑料袋中,贴上标签,做好记录,在实验室内反复按四分法弃取,留下所需的土样进行酶活性的测定。
1.3分析项目与方法[11]
土壤容重采用环刀法测定;土壤含水率采用铝盒烘干称质量法测定;土壤总孔隙度依据容重算得;土壤pH值采用电位法测定;土壤蔗糖酶活性采用二硝基水杨酸比色法测定;脲酶活性采用比色法测定;磷酸酶活性采用苯磷酸二钠比色法测定;数据采用SPSS 19.0以及Excel软件处理。
2结果与分析
2.1土壤基本理化性质分析
土壤容重、含水率、总孔隙度以及pH值都是可以反映土壤质量的基础指标。由图1可知,与A1(对照组)相比,除了A2建植模式下土壤容重发生了一定程度的减小以外,其余13种建植模式的土壤容重都有一定程度的增大,其中单植灌木模式下容重增大幅度最为明显。原因可能是土壤容重是由土壤孔隙度和土壤固体的数量决定的,可作为判断土壤肥力状况的指标之一;灌木根系较为发达,有利于透水、通气、保持土壤疏松,土壤孔隙和固体数量均增多,因此土壤容重增大最明显。卞正富等在毛乌素沙漠区进行的研究表明,植被的根系深度是决定不同深度内土壤含水率高低的主要因素[12]。本研究所用植物均为较短生长周期植物,根系较浅,受植被覆盖度的影响较小,因此土壤含水率变化均不明显。总孔隙度在14种建植模式中均出现降低现象,其中单植草本模式下降低幅度不明显,单植灌木模式下降低幅度明显且最大,草灌混植模式降低幅度次之。pH值在三大类模式下均没有明显变化。
2.2土壤酶活性垂直剖面分布特征
3类建植模式下蔗糖酶活性(图2-a)、脲酶活性(图2-b)均显著高于对照组,其中蔗糖酶活性增长幅度在B模式时最大,平均达到 91.28%,A模式时最小,但平均也达到了5526%,脲酶活性在A模式时平均增长幅度达到50%,碱性磷酸酶活性与对照组差异不显著(图2-c)。不同植被恢复模式间土壤酶活性都有一定程度的差异,其中蔗糖酶活性差异最明显。与对照组相比,0~10 cm土层蔗糖酶活性增加50.98%~117.64%,10~20 cm 土层酶活性增加55.26%~121.05%,其中以单植灌木模式对土壤蔗糖酶活性的改善效果最好。
同一种酶活性在垂直空间上存在变化,土壤上层酶活性普遍高于土壤下层。从图2可以看出,在不同植被建植模式中均表现为下层土壤酶活性低于上层,其中蔗糖酶、脲酶、碱性磷酸酶活性的空间分布特征基本一致,都随剖面深度的增加酶活性呈下降趋势,但降低幅度不同,分别降低了 0.001 3~0.002 7 mg/g,0.000 0~0.000 1 mg/g,0.000 7~0000 9 mg/g。可见土壤酶活性在垂直分布上均表现出随土层深度的增加酶活性减弱的趋势,这与以往陈立明等研究结果[13-14]一致,也符合毛志刚等关于土壤酶活性的垂直分布特征及规律的研究结论[15-16]。究其原因可能是土壤酶主要来源于微生物、动植物残体及植物根系,0~10 cm土层植物残体及微生物分布最多,根系分布密集,根系能向土壤释放酶,植物残体可通过自身的腐烂分解释放酶进入土壤,也可通过对土壤动物和微生物区系的作用而间接影响土壤酶活性,土壤表层积累了较多的枯枝落叶和腐殖质,这为微生物的生长提供了营养来源。同时,微生物的活动增强,加速了物质的转化,提高了土壤有机物质含量。0~10 cm土层水热条件和通气状况好,微生物代谢增强,使表层的土壤酶活性较高。随着土层深度的增加,土壤中水、肥、气、热状况越来越差,土壤熟化程度、肥力水平及营养元素状况都不利于微生物的活动与繁殖,导致酶活性较低[17]。土壤酶活性的这种空间分布特征表明,土壤有机质分解、土壤营养元素循环与土壤剖面结构息息相关。
2.3土壤酶活性对不同建植模式的响应
2.3.1土壤蔗糖酶活性对建植模式的响应由图3可以看出,同一建植模式下不同土层土壤蔗糖酶活性变化明显,A1~A5 模式下酶活性随着狗牙根栽植密度的增加呈现直线上升趋势,A6模式时突然下降,原因可能是狗牙根为草本植物,根系旺盛但扎入土层较浅,当其覆盖度为100%时,根系对土壤内部营养的吸收利用较强,导致土壤生物活性相对低下。B模式下酶活性呈现先下降后上升趋势, 原因是多花木兰为小灌木,根系发达,影响土壤内部生长环境,造成酶活性降低[18];随着栽植密度的增加,枯枝落叶覆盖层对土壤环境进行改良,酶活性上升(圖3)。C模式下酶活性变化趋势不显著(图3)。
不同建植模式下同一土层中土壤蔗糖酶活性均呈极明显差异(图3),活性变化范围为0.003 3~0.014 3 mg/g。与对照组相比,蔗糖酶活性在0~10 cm、10~20 cm土层中大部分有较为明显的增加,但A6模式却与整体趋势相悖,表现为降低,原因可能是草本植被覆盖度100%时,地表因照不到阳光而出现低温现象,从而使得蔗糖酶活性急剧减弱,其中B1模式下蔗糖酶活性最高,变化也最明显,原因可能是多花木兰属于豆科植物,当栽植密度为20%时,根部根瘤菌对土壤内部环境的改良效果相对最好,土壤动物及微生物也最多,对蔗糖酶活性的影响最强。B1模式下酶活性在上、下土层的差别也最明显,原因是多花木兰属于小型灌木,根部发达而且扎入土层较深,根部上端侧根多于下端,对土壤酶活性的有益影响表现为上端>下端,因此酶活性在B1模式下上层显著强于下层。
2.3.2土壤碱性磷酸酶活性对建植模式的响应如图4所示,同一建植模式下不同土层土壤碱性磷酸酶活性变化十分明显,三大类建植模式土壤酶活性均表现为上层>下层,而且变化幅度有较大差异,A模式中酶活性变化幅度为0.000 6~0.000 9 mg/g,B模式中酶活性变化幅度为 0.000 5~0.001 2 mg/g,C模式中酶活性变化幅度为0.000 8~0.001 4 mg/g。
各建植模式0~10 cm土层土壤碱性磷酸酶活性差异明显,变化范围为0.000 6~0.002 6 mg/g,C3模式下碱性磷酸酶活性表现为最强,且与其他建植模式区别最大(图4),这主要是由于C3模式中多花木兰栽植密度为60%,狗牙根栽植密度为40%,分布比例较为平均,能很好地发挥豆科植物根系对土壤环境的改良作用,同时保证狗牙根根系数量对土层较弱的影响,这既能积累充分的营养源以利于微生物旺盛生长,又促进了土壤生物的正常活动,影响了土壤生物的代谢产酶能力,使土壤磷酸酶活性呈现上述趋势。土壤碱性磷酸酶活性在 10~20 cm 土层中,3类建植模式下酶活性变化均不明显,原因可能为植被建植时间较短,土壤中有机磷的积累较少,导致生物对磷的分解转化较少,有效性较低。
2.3.3土壤脲酶活性对建植模式的响应从图5可以看出,同一建植模式下不同土层土壤脲酶活性,除了A6及B1模式变化较小外,其他各建植模式不同土层变化幅度均比较一致,约为0.000 1 mg/g。
土壤脲酶活性测定值变化范围为0.000 1~0.000 4 mg/g,变化范围较小,而且变化趋势不明显,无论是同一建植模式下不同土层间,还是同一土层不同建植模式下,其活性在整个土壤剖面中受建植模式影响均不明显,虽然对于脲酶活性空间分布的研究报道已有很多,但是建植模式是否影响脲酶活性尚没有确切的定论。土壤脲酶对促进土壤氮素循环和提高土壤氮素的利用率有重要意义[19],脲酶活性与土壤呼吸强度、土壤微生物活动均相关,在一定程度上反映了土壤微生物活动的强度,而微生物在复杂生态系统中分布较多,由此推断脲酶活性应该表现出与其他酶活性相类似的分布特征,但本试验分析表明,脲酶活性被植被影响的程度相对于蔗糖酶、碱性磷酸酶活性不明显,原因可能是多花木兰虽然为豆科植物,根部根瘤菌有一定的固氮作用,但由于灌木植株较少,对土壤供氮能力的影响较弱,因此土壤脲酶活性变化程度均不明显。
2.3.4土壤基本性质与酶活性的相关关系通过相关分析发现,三大类建植模式土壤酶活性之间,及土壤酶活性与土壤基本理化性质之间均存在一定的相关性(表2),其中土壤蔗糖酶活性与脲酶活性均呈现出极显著的正相关关系,与磷酸酶活性以及pH值呈正相关,其中在B、C建植模式下存在的相关关系均达到了极显著、显著水平;与总孔隙度间的相关性均为负相关,在A模式下相关性极显著,B模式下相关性显著。土壤脲酶活性在三大类建植模式下,与磷酸酶活性均有正相关关系,其中在A、C模式下相关性达到了极显著水平;与pH值之间的正相关关系在B模式下极显著;与总孔隙度为负相关关系,且都达到显著水平。土壤碱性磷酸酶活性与pH值和含水率的正相关关系在三大类建植模式下均不显著;与总孔隙度均呈负相关关系,在A模式下相关性极显著,C模式下相关性显著。
3结论与讨论
(1)三大类14种建植模式中,除了单植草本A2模式下发生了一定程度的减小以外,其余13种建植模式的土壤容重都有一定程度的增大,其中单植灌木模式下容重增大幅度最为明显;土壤含水率变化均不显著。总孔隙度在三大类14种建植模式中出现降低现象,其中单植草本模式下降低幅度不明显,单植灌木模式下降低幅度明显且最大,草灌混植模式降低幅度次之。pH值在3类模式下均没有明显变化。
(2)三大类14种建植模式下,土壤蔗糖酶、脲酶、碱性磷酸酶活性的空间分布特征基本一致,都随剖面深度的增加呈下降趋势,但减小幅度不同。同一建植模式不同土层中,蔗糖酶和碱性磷酸酶活性随土层深度增加下降,0~10 cm土层酶活性明显高于10~20 cm土层,脲酶活性变化未达到显著水平。
(3)不同建植模式同一土层中,土壤蔗糖酶、碱性磷酸酶活性均有较明显的差异,其中当单植多花木兰栽植密度为20%时蔗糖酶活性最高,单植狗牙根栽植密度为100%时蔗糖酶活性最低,狗牙根与多花木兰混植模式下蔗糖酶活性最稳定;各建植模式同一土层土壤碱性磷酸酶活性差异显著,碱性磷酸酶活性大小在狗牙根栽植密度60%+多花木兰栽植密度40%时表现为最强;土壤脲酶活性在各建植模式不同土层变化幅度均比较一致,变化范围均较小,无论是同一建植模式下不同土层,还是不同建植模式下同一土层,其活性在整个土壤剖面中受建植模式影响均不明显(从数量级看)。
(4)三大类建植模式下,土壤酶活性之间表现出不同的差异性,并且不同建植模式土壤酶活性之间都存在一定的显著或极显著正相关关系。土壤酶活性均与pH值呈现正相关关系,与总孔隙度呈负相关关系,但相关程度存在较大差异;土壤碱性磷酸酶活性与含水率呈现正相关关系,但不显著。
(5)含碎石土壤不同建植模式下,土壤酶活性随着建植植被种类及复杂程度的变化而变化,植被类型不仅影响土壤质量,而且对土壤酶活性也具有较大的影响。与单植草本模式的土壤相比,单植灌木以及草灌混植模式土壤酶活性均维持在较高的水平,其中草灌混植模式下酶活性不仅较高而且较为稳定。这也说明通过复杂种类植被恢复措施,能够逐渐促进含碎石土壤生态环境的改善。本研究还存在很多不足,期望在此基础上,进一步加强土壤生态环境的跟踪监测,可以将土壤微生物含量和土壤酶活性、土壤理化性质结合起来作为评价含碎石土壤植被建植模式改良效果的综合指标。
参考文献:
[1]陈影,张利,董加强,等. 废弃矿山边坡生态修复中植物群落配置设计——以太行山北段为例[J]. 水土保持研究,2014(4):154-157.
[2]王永生. 矿山地质环境治理存在问题及对策[J]. 南方国土资源,2009(12):36-37.
[3]武丽花. 植被修复对湘潭锰矿矿区废弃地土壤环境效应的影响[D]. 长沙:中南林业科技大学,2008.
[4]王宗永. 杉木人工林林下固氮植被的筛选及其生态经营研究[D]. 长沙:中南林业科技大学,2011.
[5]孙红. 贵州高原禾草+白三叶草地养分及植被异质性形成研究[D]. 蘭州:兰州大学,2014.
[6]王世雄,王孝安,李国庆,等. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释[J]. 生态学报,2010,30(6):1638-1647.
[7]何斌,黄承标,秦武明,等. 不同植被恢复类型对土壤性质和水源涵养功能的影响[J]. 水土保持学报,2009,23(2):71-74.
[8]宋娟丽,吴发启,姚军,等. 弃耕地植被恢复过程中土壤理化性质演变趋势研究[J]. 干旱地区农业研究,2009,27(3):168-173.
[9]李妮,陈其兵,谭昌明. 高速公路边坡乡土灌木建植模式水土保持效益[J]. 水土保持学报,2012,26(1):67-71.
[10]孙铁军,肖春利,滕文军. 不同草地建植模式对荒坡地土壤风蚀及理化性质的影响[J]. 水土保持学报,2011,25(3):44-48.
[11]关松荫. 土壤酶及研究方法[M]. 北京:农业出版社,1986.
[12]卞正富,雷少刚,常鲁群. 基于遥感影像的荒漠化矿区土壤含水率的影响因素分析[J]. 煤炭学报,2009,34(4):520-523.
[13]陈立明,满秀玲. 云冷杉林土壤酶活性与土壤养分的研究[J]. 中国水土保持科学,2009,7(4):94-99.
[14]张仕艳,原海红,陆梅,等. 滇西北不同利用类型土壤酶活性及其理化性质与微生物的关系[J]. 亚热带水土保持,2010,22(2):13-16.
[15]毛志刚,谷孝鸿,刘金娥,等. 盐城海滨湿地盐沼植被及农作物下土壤酶活性特征[J]. 生态学报,2010,30(18):5043-5049.
[16]杨成德,陈秀蓉,龙瑞军,等. 东祁连山高寒草地牧草返青期土壤酶活性特征[J]. 草地学报,2010,18(3):308-313.
[17]司登宇,张金池,陈莉莎,等. 浙江省凤阳山不同林分类型土壤酶活性研究[J]. 水土保持通报,2013,33(6):258-263.
[18]夏雪,刘善江,陈桂梅,等. 土壤酶的研究进展[J]. 中国农学通报,2011,27(21):1-7.
[19]范君华,刘明,高疆生,等. 塔里木河上游不同林地土壤养分和微生物以及酶活性的变化初探[J]. 中国农学通报,2005,21(1):184-188.
