
宽叶蓝靛果在不同海拔分布的生物多样性分析
2017-05-17 11:40张巍孟庆彬王洪刚邵明海
森林工程 2017年3期
张巍,孟庆彬,王洪刚,邵明海
(伊春林业科学院,黑龙江 伊春 153000)
宽叶蓝靛果在不同海拔分布的生物多样性分析
张巍,孟庆彬,王洪刚,邵明海
(伊春林业科学院,黑龙江 伊春 153000)
蓝靛果忍冬是我国具有重要经济价值的浆果资源,而宽叶蓝靛果是蓝靛果中较为特殊的品种,具有很高的研究价值。为研究宽叶蓝靛果物种多样性的垂直变化效应,进一步揭示在不同海拔梯度中宽叶蓝靛果群落的结构稳定性,在小兴安岭伊春林区以600、1 000、1 400 m三个海拔梯度为基础,共设临时样地7块,计算种群内重要值及各相关指数。结果表明:①海拔600m群落灌木层多样性指数为各海拔区域数值最高。海拔1 400 m群落多样性指数为各海拔梯度最低。②海拔1 000~1 400 m,丰富度指数总体处于0.11~0.12,没有梯度变化。③群落主要伴生树种以蔷薇科树种居多,多以东北珍珠梅与宽叶蓝靛果伴生形成稳定群落结构。
宽叶蓝靛果;生物多样性;种群分布
0 引言
生物多样性是指一个区域内生命形态的丰富程度,是生物及其与环境有规律地结合所构成稳定的生态综合体以及与此相关的各种生态过程的总和[1]。生物多样性沿环境梯度的变化规律是生物多样性的研究一个重要议题[2-3]。近年来在小兴安岭伊春林区发现宽叶蓝靛果忍冬(Loniceraedulisvar.turczaninowii(Pojark.))的野生资源分布[4-5]。通过对所找到的宽叶蓝靛果多个野生分布区的调查证明,其在伊春林区野生分布的海拔约为550~1 400 m左右,低海拔地区未发现分布。
随着市场需求的增大,小浆果资源受到越来越多的关注。目前我国引进的蓝靛果品种有40个左右[6-7]。这说明,做为一种新型浆果资源,蓝靛果的发展潜力巨大。本文通过对宽叶蓝靛果在不同海拔梯度的分布格局的调查,分析宽叶蓝靛果群落物种多样性的变化规律,研究结果可以丰富宽叶蓝靛果群落生态学的内容,并为宽叶蓝靛果群落生物多样性保护及资源利用提供理论依据。
1 分布区概况
至目前,共确定伊春林区的宽叶蓝靛果野生分布区三处,分别在带岭林业局,南岔林业局和桃山林业局。
南岔林业局位于东经129′28″,北纬47′132″,积温全年10.0 ℃以上在2 200~2 300 ℃左右。年极端最高气温为38.2 ℃,年极端最低气温为-44.2 ℃,年平均降雨量600 mm,日照时数1 236.4 h,无霜期115 d。宽叶蓝靛果主要分布于石头河林业局黄花山,海拔550~670 m处。在黄花山野生分布中,蓝靛果忍冬(LoniceraedulisTurcz)与宽叶蓝靛果忍冬(va.venulosa(Max)Rehd.)混生分布,在其它海拔分布少有发现此类情况,林型较为典型。
带岭林业局位于东经129′02″,北纬47′02″,平均海拔418.6 m,最高海拔为大菁山1 250 m。年均温1.4 ℃,≥10 ℃积温2 663.1 ℃,日照时数2 156.2 h,无霜期115 d,年降水量661 mm。宽叶蓝靛果主要分布于大菁山海拔1 000~1 200 m处,分布广泛。是目前为止伊春林区所找到的分布面积最大的地区。
桃山林业局位于东经128′171″,北纬46′936″,平均海拔300 m,最高海拔为平顶山1 429 m,为小兴安岭最高点。桃山林业局年平均气温为2.8 ℃,≥10 ℃,年积温2 153.2 ℃,日照时数2 318.1 h,无霜期113 d,年平均降水量665 mm。宽叶蓝靛果主要分布于平顶山海拔1 050~1 400 m处,海拔1 300 m分布最为广泛,南北坡均有分布。
2 研究方法
2.1 样地调查方法
依据三个种源地宽叶蓝靛果的野生分布面积,分别在黄花山海拔600 m设临时样地两块。大菁山海拔1 000 m设临时样地3块。平顶山海拔1 400 m设临时样地两块,样方面积统一为30 m×30 m。调查所有乔、灌木。调查内容包括种类、株数、株高、冠幅、盖度等。灌木增加调查株丛分枝数等指标。每个小样方中,设1 m×1 m草本样方,记录种类、株高和盖度等。
2.2 物种多样性统计方法
参照马克平等人的研究方法[8-10],多样性统计方法采用丰富度指数、Shannon-Wiener指数、Simpson指数、Pielou均匀度指数等进行测定。
(1)丰富度指数:物种丰富度指数R用群落中物种的总数S进行计算,公式为:

式中:N为样方中观察到的所有物种的个体总数;S为群落中物种的总数。
(2)物种多样性:物种多样性采用目前应用最为广泛的Shannon-Wiener指数H和Simpson优势度指数D表示,公式为:
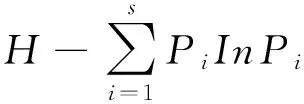

式中:N、S意义同上;ni为第i个物种的个体数。Pi为第i个物种的个体数占总个体数N的比例,Pi=ni/N。
(3)物种均匀度:均匀度是指样地中各个物种多度的均匀程度,即每个物种个体数间的差异(基于Shannon-Wiener指数的Pielou均匀度指数J的计算公式为:

式中:J意义同上;H为Shannon-Wiener指数。
3 结果与分析
3.1 不同海拔梯度群落重要值分析
对宽叶蓝靛果各海拔群落树种进行分类,结果显示,宽叶蓝靛果野生种群伴生植物共有21科15属。其中乔木层5科7属,灌木层3科6属,草本层13科21属。重要值是反映植物种类在群落中地位重要程度的综合指标之一[11]。根据不同海拔,将宽叶蓝靛果野生种群以乔木层、灌木层、草本层进行分类及计算重要值[11-13]。分析可知,宽叶蓝靛果在不同海拔的群落结构均存在一定差异,重要值也有所不同(表1、表2)。
由表1可知,海拔600~1 400 m乔木层平均株高分别为7.28、6.4、3.87 m。在宽叶蓝靛果不同海拔梯度构成中,乔木层主要构成树种为红皮云杉(Picea koraiensis Nakai)、花楷槭(Acer ukurunduense Trautv.et)等。而随着海拔梯度升高,树种株数、盖度等指标也程负相关走势。乔木层还包括大黄柳(Betula platyphylla Suk)、毛赤杨(Alnus sibirica Fisch.ex Turcz)等树种,但多丛生或胸径较小,所以没有纳入重要值计算。
灌木层最典型的伴生树种主要是东北珍珠梅(Spodiopogon cotulifer),由表1可知,东北珍珠梅在所有海拔均有野生分布,植被覆盖度也相对较高,重要值系数大,证明在宽叶蓝靛果群落中占有十分重要的作用,是最主要的伴生树种。由其在海拔达到1 400 m后,群落灌木层主要伴生树种只有东北珍珠梅一种,它与宽叶蓝靛果伴生形成稳定的群落结构。另外在1 000 m以下的低海拔群落中,柳叶绣线菊(Spiraca salicifolia Linn)、大果蔷薇(Rosa albertii Reg.)等在群落中也具有重要地位。

表1 不同海拔梯度乔、灌层树种构成Tab.1 Tree species composition of arbor and shrub layer at different elevation gradients
由表2可以看出,草本层在群落构成较为丰富。但不同海拔梯度草本层构成种类有所差异。海拔600 m群落主要构成品种为柳兰、龙江凤毛菊等。海拔1 000 m主要构成为草乌头、林问荆、白芷等。海拔1 400 m主要由唐松草、单穗升麻等构成。而大叶樟、猴腿蹄盖蕨在各海拔均有分布,且重要值较高,证明在群落中占有重要地位。
总体分析,因为宽叶蓝靛果忍冬群落一般为郁闭度较低,光照充足的林型,所以乔木层在宽叶蓝靛果群落中所占的重要值不高,灌木层和草本层在群落中起到了重要作用。而灌木层和草本层重要值没有随海拔梯度变化而变化,是由于灌木层和草本层的树种对不同海拔的适应能力相差不大造成的。
3.3 不同海拔梯度的物种多样性分析
由表3可知,在不同海拔梯度灌木层物种多样性中,海拔1 000 m以下的宽叶蓝靛果灌木层物种多样性最为丰富(图1)。而在海拔1 000 m以上,多样性指数H,丰富度指数R和优势度指数D较小,且数值相近,处于同一区间。均匀度指数J随海拔升高而降低呈减小趋势,证明高海拔区域对宽叶蓝靛果群落稳定性产生一定影响。
由表1、表3可知,虽然灌木层在海拔1 000 m种群种类最为丰富,但丰富度指数仍然较低,这是因为一般灌木树种分布较少,重要值较低,建群的主要树种就是宽叶蓝靛果和东北珍珠梅组成的群落结构。由此也证明,东北珍珠梅对此生态环境适应且地位稳定,为不同海拔梯度宽叶蓝靛果群落灌木层中的常见伴生种。
海拔1 000 m群落的草本层种类最为丰富,共12科17属,其优势度指数D最高(表3)。海拔1 400 m多样性指数H,均匀度指数J最高。而在海拔1 000~1 400 m丰富度指数处于同一区间,且均高于海拔600 m。分析可知,海拔1 000~1 400 m 是高山草甸过渡地带,宽叶蓝靛果容易在开阔地形成群落,这为草本植物生长提供了良好的生长基础,因而草本植物个体分布均匀,多样性较高[12]。

表2 不同海拔梯度草本层构成Tab.2 Composition of herb layer at different elevation gradients

表3 灌、草层物种多样性Tab.3 The species diversity of shrub and grass layer

图1 海拔600 m处宽叶蓝靛果所在林型Fig.1 The broad leaf Lonicera edulis forest at an altitude of 600 m

图2 海拔1 000 m处宽叶蓝靛果所在林型Fig.2 The broad leaf Lonicera edulis forest at an altitude of 1 000 m
4 结论及讨论
(1)不同区域,不同海拔宽叶蓝靛果种群多样性在一定区间内相对稳定。主要伴生植物差异不大,但其它伴生树种有所差异,树种在群落中的重要值也有所差异。
(2)做为草甸过渡带,虽然海拔1 000 m区间物种最为丰富,但多样性、丰富度和优势度等指数仍然显示出由低海拔向高海拔递减趋势,这证明海拔梯度对宽叶蓝靛果群落各层次影响明显,呈负相关走势。
(3)宽叶蓝靛果群落在不同海拔的主要伴生树种均为东北珍珠梅,证明这一林型在小兴安岭宽叶蓝靛果野生分布中具有稳定的群落结构,并与宽叶蓝靛果伴生成为该群落的建群树种。
物种多样性是反映群落结构和功能特征的有效指标,是生态系统稳定性的量度[12-13]。而海拔的变化是决定本区生境差异的主导因子,继而制约着本区植物群落的空间分布格局[14-18]。上述分析表明,宽叶蓝靛果种群多样性随海拔梯度而改变,证明不同海拔梯度的改变可以对宽叶蓝靛果群落结构产生直接影响。另外,海拔1 000 m以上的宽叶蓝靛果群落乔、灌木树种组成较少,林份构成较为单一,这也为宽叶蓝靛果种群的形成提供了条件,但这种单一的林份结构也容易受到极端环境因子变化的影响。
[1]陈灵芝.中国的生物多样性——现状及其保护对策.[M]北京:科学出版社,1993.
[2]Kitayama K.An altitudinal transect study of the vetetation of Mount Kinabatu[J],Borneo vegetation,1992,102:149-171.
[3]郭英荣,雷平,晏雨鸿,等.江西武夷山黄岗山西北坡植物物种多样性沿海拔梯度的变化.[J].生态学杂志,2015,34(11):3002-3008.
[4]周以良,董世林,聂绍荃.黑龙江树木志[M].哈尔滨:黑龙江科学技术出版社,1986.
[5]张巍,王洪学,王洪刚,等.伊春林区蓝靛果种质资源调查研究[J].森林工程,2014,30(1):46-48.
[6]古丽江·许库尔汗.俄罗斯的蓝靛果忍冬品种[J].落叶果树,2012,44(1):62-65.
[7]古丽江·许库尔汗.俄罗斯的蓝靛果忍冬品种续[J].落叶果树,2012,44(2):60-64.
[8]赵志模,郭依泉.群落生态学原理与方法.[M].重庆:科学技术文献出版社重庆分社,1990:213-235.
[9]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究[J].生态学报,1995,15(3):269.
[10]马克平,刘灿然,刘玉明.生物多样性的测度方法.Ⅱ.多样性指数的测度方法[J].生物多样性,1995,3(1):38-39.
[11]王义弘,李俊清,王政权.森林生态学实验实习方法.[M].哈尔滨:东北林业大学出版社,1990.
[12]丁圣彦.浙江天童常绿阔叶林演替系列栲树和木荷成为优势种的原因[J].河南大学学报:自然科学版,2001 31(1):79-81.
[13]Pishancourt J B,Firn J,Chades I,et al Growing biodiverse carbonrich forests.[J].Globrl Change Biology,2014,20(2):382~393.
[14]Brown K A,Gurevitch J.Long-term impacts of logging on forest diversity in Madagascar[J].proceedings of the National Academy of Sciences 2004,101(16):6045-6049.
[15]贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[16]Ding Y Zang R,Letcher S G,et al.Disturbance regime change s the trait distribution phyloyenetic structure and commlcnity assembly of tropical rain foreste.[J].Oires,2012,121(8):1263-1270.[17]Letcher S G Chazdon R L,Rapid recovery forest chronosegrence in northeastern Costa Rica.[J].Biotropica,2009,41(5):608-617.
[18]毛波,董希斌.大兴安岭低质山杨林改造效果的综合评价[J].东北林业大学学报,2016,44(8):7-12.
Analysis of Biodiversity of the Broad LeafLoniceraedulisat Different Elevations
Zhang Wei,Meng Qingbin,Wang Honggang,Shao Minghai
(Yichun Academy of Forestry sciences,Yichun 153000)
Lonicera edulis is a berry species in China and has significant economic value.The broad leaf Lonicera edulis is a special species of berry of high research value.In order to study the vertical variation of the species diversity of broad leaf Lonicera edulis and further reveal the structural stability of its community at different elevations,seven temporary sample plots were established in Yichun Forest Region in Xiao Hinggan Mountains at 600 m,1 000 m,and 1 400 m respectively to calculate importance values and relative indexes within the community.The results showed that community diversity index at 600m shrub layer is the highest within all altitude regions,while the one at 1 400 m is the lowest;the richness index is generally between 0.11~0.12 with no gradient change in the altitude range of from 1 000 m to 1 400 m.The main species of the community is Rosaceae and most of the northeast Sorbaria sorbifolia associated with broad leafLoniceraedulisto form a stable community structure.
broad leaf ofLoniceraedulis;biodiversity;species distribution
2016-10-24
黑龙江省森工总局科技支撑项目(sgzjY2014024)
张巍,硕士, 工程师。研究方向:森林特产和森林经营。E-mail:zw81025259@163.com
张巍,孟庆彬,王洪刚,等.宽叶蓝靛果在不同海拔分布的生物多样性分析[J].森林工程,2017,33(3):28-32.
S 718
A
1001-005X(2017)03-0028-05
猜你喜欢
中国地市报人(2022年7期)2022-07-21
环境保护与循环经济(2021年12期)2021-03-16
矿产勘查(2020年5期)2020-12-25
当代贵州(2018年28期)2018-09-19
森林工程(2018年3期)2018-06-26
乡村科技(2018年8期)2018-02-13
绿洲(2017年5期)2017-10-10
农家科技下旬刊(2017年6期)2017-07-05
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
