
不同环境因子对梭梭种子萌发的影响
2017-04-26 14:41彭梦文刘晓龙郭帅马雨荷王红燕
安徽农学通报 2017年7期
彭梦文+刘晓龙+郭帅+马雨荷+王红燕
摘 要:该文研究了温度、盐分、干旱等环境因子对梭梭种子萌发的影响。结果表明:梭梭种子萌发速率随着温度的降低而减慢,不同温度条件下,梭梭种子的最终萌发率均在90%以上;较高浓度的Nacl溶液,对梭梭种子萌发进程具有抑制作用,当Nacl浓度大于44g/L时,最终萌发率和萌发速率显著降低,盐胁迫复水后的萌发恢复率均在60%以上;梭梭种子只能在渗透势为0~-2.4MPa的条件下萌发,随着干旱胁迫的加强,梭梭种子的萌发率和萌发速率逐渐降低,干旱胁迫复水后的萌发恢复率均在70%以上。通过研究梭梭种子对温度的广泛适应性,以及在盐胁迫和干旱胁迫后种子复水萌发的能力,探索梭梭种子萌发机制,为荒漠区植被恢复和重建提供参考。
关键词:种子萌发;梭梭;温度;盐胁迫;干旱胁迫
中图分类号 S793 文獻标识码 A 文章编号 1007-7731(2017)07-0033-05
Abstract:The influences of temperature,salinity and drought stress on Haloxylon ammodendron seed germination were studied. The results showed that the germination rate reduced when the temperature decreased,its final germination percentage could reach more than 90% in different temperature levels. The germination percentageand germination ratedecreased when the salinity concentration increased,the germination decreased significantly when the concentration of NaCl solution exceeded 44g/L.When the incubated seeds treated in NaCl solution were transferred to distilled,the recovery rate of the germination was more than 60%. H.Ammodendron seed only germinated when the solution osmotic potential higher more than -2.4MPa.With the increase of osmotic stress,H.Ammodendron seeds germination percentage decreased significantly. The re-germination percentage could reach more than 70% after osmotic stresstreatments. We concluded that H.Ammodendron had extensive adaptability to temperature conditions and the re-germination ability after salinity and drought stress.
Key words:Seed germination;Haloxylon ammodendron;Temperature;Salt stress;Drought stress
梭梭(Haloxylon ammodendron)属于藜科多叶旱生植物,是分布于中国西北地区的多年生灌木,具有在干旱、盐碱、营养不良、强风、沙土移动和在强光下生长的特性[1]。其抗旱、抗热、抗寒、耐盐碱性较强,茎枝内盐分含量高达15%左右,喜光,不耐庇荫,适应性强,生长迅速,枝条稠密,根系发达,防风固沙能力强,是我国西北和内蒙古干旱荒漠地区固沙造林的优良树种,也可以作为牲畜的饲料。作为地中海植物区系的重要组成植物,梭梭主要广泛分布于准噶尔盆地、塔里木盆地以及一些荒漠地区,是一种典型的沙生植物[2]。
关于各种环境因子对梭梭种子萌发的影响已有大量报道[3-5],其中,温度是决定种子萌发的一个重要因素[6]。一般来说,种子萌发的温度被认为在10℃以上[1,7]。然而一些研究者表明有些物种可以在低温条件下完成萌发。周平等研究表明白梭梭(Halaxylon persicum Bunge)种子在2℃时就可以萌发,当积雪厚度达到22cm时,白梭梭萌发率达到80%以上[4]。韩建欣等在融雪期间开展了高枝假木贼(Anabasis elatior)胎生萌发特性的野外观测,并进行了低温(5/2℃)萌发实验,认为低温只是降低了种子萌发速度而对总萌发率没有影响[8];而Shimono and Kudo发现前胡属多管藁本(Peucedanum multivittatum)种子在低温(0℃)处理期间就可以萌发,并观测到了雪床生境中的种子在融雪期间就已经出苗[9]。目前,对于梭梭种子萌发的温度条件主要集中于5℃以上,关于更低温度对种子萌发的影响鲜有报道,本文通过模拟野外温度环境,在实验室条件下,开展了低温(5/2℃、2℃、2/-3℃)萌发实验,对梭梭种子能否适应低温环境而萌发进行研究。关于盐分和干旱条件对梭梭种子萌发的影响有较多报道,但胁迫处理后对种子进行复水实验,探测种子是否还具有萌发活力的内容还不够全面。因此,本文通过研究梭梭种子对低温的适应性,以及在经历胁迫后种子复水萌发的能力,揭示梭梭种子对于温度、盐分、干旱的适应范围,用于阐明梭梭种子在野外的萌发特性,为荒漠地区植被恢复与重建提供科学依据。
1 材料与方法
1.1 供试材料 梭梭种子于2016年10月采自准噶尔盆地南缘的克拉玛依农业开发区自然种群中,在室温下贮藏。
1.2 研究方法
1.2.1 不同温度处理下种子的萌发 选择籽粒饱满、发育良好的成熟种子,设置实验温度分别为20/15℃、15/10℃、10/5℃、5/2℃、2℃、2/-3℃。将种子放置在具有湿润滤纸的培养皿中,每组40粒、3个重复,以胚根出现为种子萌发标准,定期观察萌发情况并记录其发芽数,计算发芽率。
1.2.2 不同盐胁迫处理下种子的萌发 配置浓度为2g/L、4g/L、6g/L、8g/L、12g/L、16g/L、20g/L、24g/L、30g/L、36g/L、44g/L、52g/L、60g/L、68g/L的NaCl溶液,以蒸馏水为对照,将种子放置在具有湿润滤纸的培养皿中,每组40粒、3个重复,在20℃/10℃(光照/黑暗=12h/12h)条件下进行萌发,每天记录萌发情况。
1.2.3 不同干旱胁迫处理下种子的萌发 在20℃/10℃下,采用模拟干旱的PEG-6000溶液,溶液的渗透势为0,-0.3,-0.6,-0.9,-1.2,-1.5,-1.8,-2.1,-2.4,-2.7和-3.0MPa。将种子放置在具有湿润滤纸的培养皿中,每组40粒、3个重复,在20℃/10℃(光照/黑暗=12h/12h)条件下进行萌发,每天记录萌发情况。
1.2.4 复水实验 胁迫实验进行到第8d,将培养皿中未萌发的种子取出,用蒸馏水进行冲洗过后,放置在加入蒸馏水的湿润培养皿中,每个培养皿垫有两层滤纸。以胚根出现为种子萌发标准,定期观察萌发情况并统计其萌发数,计算萌发率。萌发率计算公式:最终萌发率(GF)=Σn/N。
1.3 数据处理 本试验数据分析采用Excel 2007和SPSS 18.0软件,绘图采用ORIGIN 7.5软件。
2 结果与分析
2.1 不同温度对梭梭种子萌发的影响 通过10d的培养发现,随着时间变化,梭梭种子的萌发率总体呈增长趋势(图1)。随着温度的降低,梭梭种子萌发率的增长变得缓慢。温度为5/2℃、2℃、2/-3℃时的萌发率增长曲线明显比20/15℃、15/10℃、10/5℃温度下的增长曲线平缓,在20/15℃温度条件下培养的梭梭种子,实验开始的24h内就出现萌发现象,并在实验的2~3d发芽达到高峰,第4d完成萌发,最终萌发率达到98.8%。在15/10℃温度条件下的梭梭种子,实验的2~4d发芽达到高峰,第6d基本完成萌发,最终萌发率为98.4%。当温度为10/5℃时,梭梭种子在实验的1~4d内萌发率逐渐增加,实验的第4~7d,萌发率增长迅速,随后萌发率达到最大值97.2%不再变化。当温度为5/2℃时,梭梭种子的萌发速率明显减弱,尤其在实验的1~4d萌发率较低,实验的第9d萌发率达到最大值95.6%。在温度为2℃、2/-3℃的低温条件下,种子萌发缓慢,完成萌发的时间较长,但最终萌发率均达到90%以上(图1,表1)。由此可知,梭梭种子在各个温度梯度下均能萌发,且萌发率差异不显著,低温仅仅减缓了种子萌发速率,延迟萌发时间,但是对最终的萌发率没有影响(表1)。
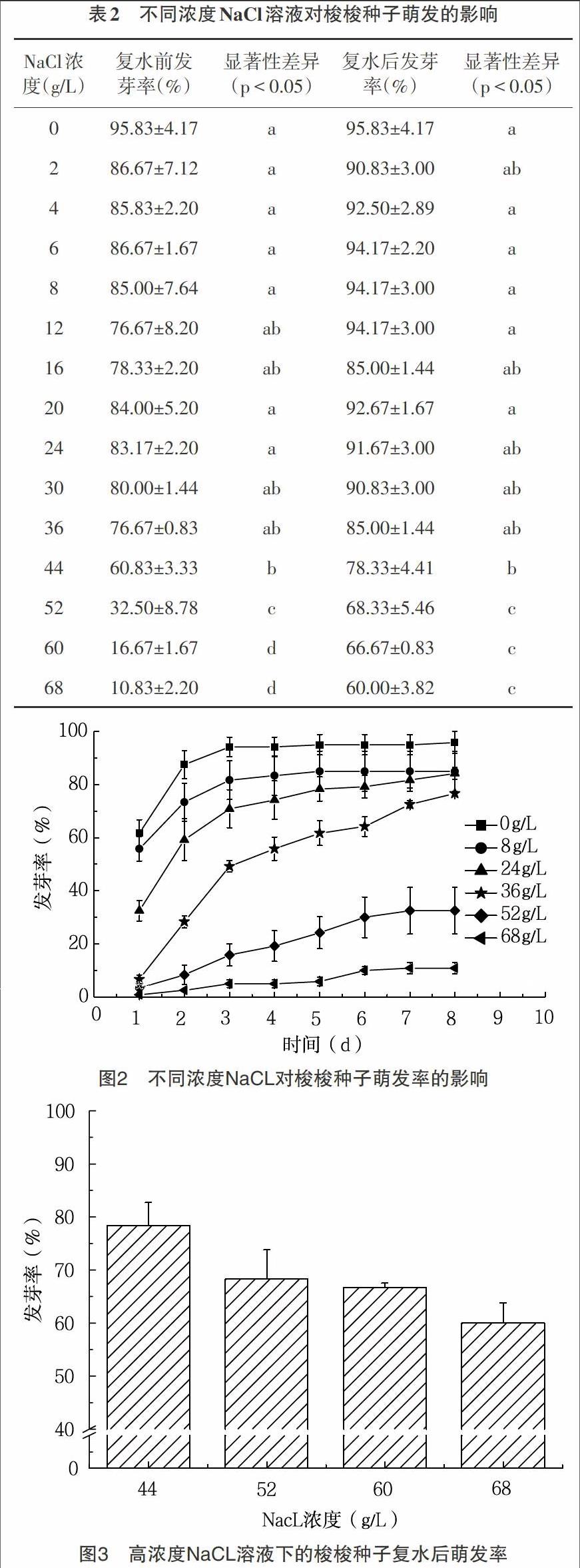
2.2 不同浓度NaCl溶液处理对梭梭种子萌发的影响 梭梭种子的萌发率与NaCl浓度整体呈反比,即随着NaCl浓度的增加,梭梭种子的萌发率逐渐降低(图2,表2)。当NaCl浓度低于36g/L时,梭梭种子的萌发率较高,基本处于80%以上,且萌发速度较快。当NaCl浓度高于36g/L时,梭梭种子的萌发率逐渐呈下降趋势。当NaCl浓度高于44g/L时,梭梭种子萌发速率显著下降。实验至第8d,将未萌发的梭梭种子进行复水实验,种子萌发迅速且萌发恢复率较高,将NaCl处理浓度低于36g/L时的梭梭种子进行复水,最终发芽率均达到90%以上,将高浓度(44g/L、52g/L、60g/L、68g/L)的NaCl处理下的种子进行复水,萌发率增加至60%以上(图3,表2)。浓度高于44g/L的NaCl溶液中的梭梭种子萌发率和低于44g/L的NaCl溶液中的梭梭种子萌发率之间具有显著性差异,说明高浓度的NaCl盐溶液抑制了种子萌发(表2)。
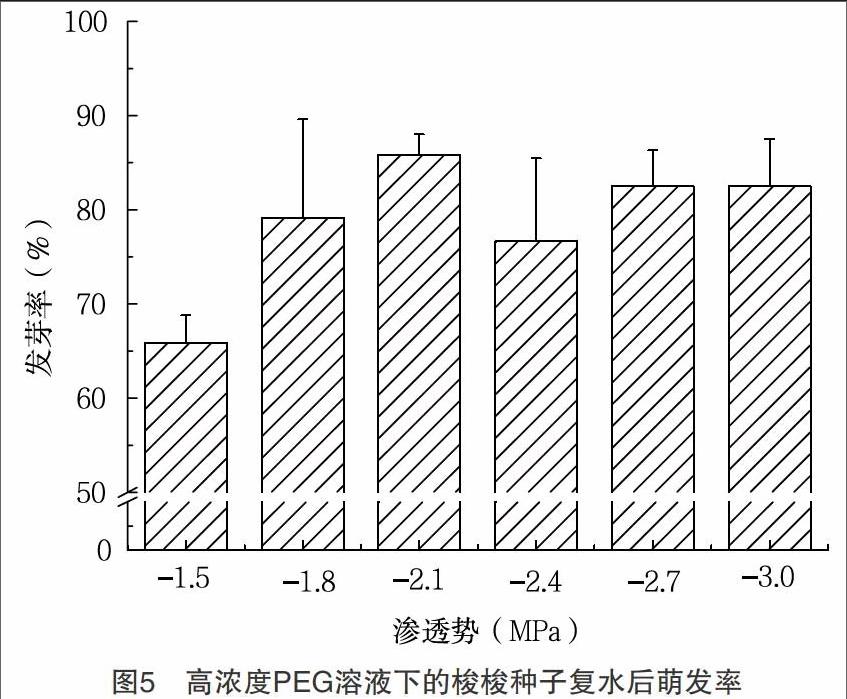
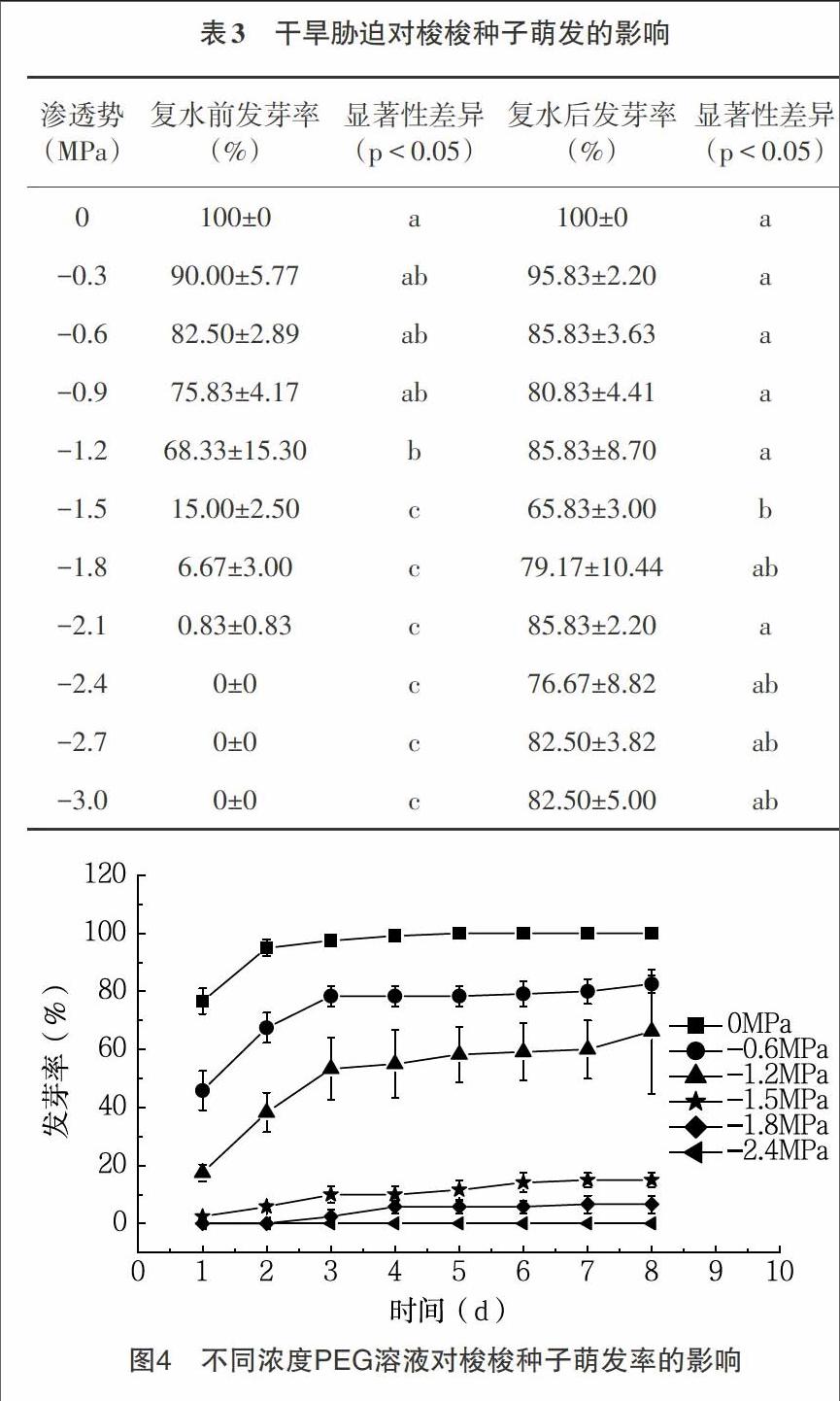
2.3 干旱胁迫对梭梭种子萌发的影响 梭梭种子在不同渗透势的PEG溶液中具有不同的萌发率,并且随着干旱胁迫强度的增强,萌发率显著降低(表3)。在PEG溶液渗透势为0MPa、-0.3MPa、-0.6MPa、-0.9MPa、-1.2MPa 时,梭梭种子在1~4d内萌发较快,随后种子萌发率缓慢增长最终达到稳定值不再变化。PEG溶液渗透势为-1.5MPa、-1.8MPa、-2.1MPa时,梭梭种子萌发速率显著下降,且萌发率低。渗透势为-2.4MPa、-2.7MPa、-3.0MPa时的萌发率为0(表3,图4)。实验至第8d,将未萌发的种子进行复水试验,各个渗透势的PEG溶液下的梭梭种子,绝大部分完成萌发,萌发率显著增加,并且基本在80%左右(表3)。在PEG溶液浓度较高的条件下(渗透势为-1.5MPa、-1.8MPa、-2.1MPa、-2.4MPa、-2.7MPa、-3.0MPa)培养的种子,经复水试验后,最终也达到较高的萌发率(图5)。在复水试验前,各个渗透势的PEG溶液下种子萌发率不同,PEG溶液渗透势大于-1.5MPa时梭梭种子萌发率与渗透势小于-1.5MPa时的种子萌发率之间差异显著,滲透势在-1.5~-3.0MPa间梭梭种子萌发率之间差异不显著,且均为0。解除PEG胁迫后,各个渗透势处理的梭梭种子的萌发率差异显著,且渗透势在-1.5~-3.0MPa梭梭种子都出现迅速萌发现象,说明干旱胁迫会抑制梭梭种子的萌发(表3)。
3 讨论与结论
3.1 讨论 种子萌发是植物生活史中的重要阶段,但是此阶段在时间和环境条件上却存在很大的不可预测性[10],气候变化间接影响植物变化[11-12]。适宜温度是荒漠植物种子萌发的先决条件,喜温的荒漠植物种子要求的温度通常是15~25℃[13],不同的荒漠植物种子萌发都有其最低、最适和最高温度。适温时,荒漠植物种子萌发最快,萌发率也最高;温度较高时,种子萌发速度与适温时相差不大,但发芽率则随温度升高而降低[14],本研究结果表明,梭梭种子的最适萌发温度为20/15℃和15/10℃,随着温度的降低,梭梭种子的萌发速率减慢,但各个温度下的最终萌发率没有差异,均表现出高萌发率,说明梭梭种子对于温度的适应范围较广,低温条件仅仅减弱了种子萌发速率,推迟了种子完成萌发的时间,但最终萌发率与最适温度下的最终萌发率无显著差异,梭梭在低温环境下的萌发特性也许是利用了早春积雪融化时期良好的土壤条件,从而在经历春季干燥期之前促进幼苗的生长和木质化的形成。在低温条件下完成萌发,表现出梭梭种子能够适应荒漠区恶劣环境,从而促进种子建苗的萌发机制,这对植被在荒漠区成功建群具有很大的意义。
我国西北干旱荒漠区降水稀少,蒸发强烈,导致盐分聚留在土壤表层,形成大量盐渍化生境,盐渍化土壤使植物的生长发育受到严重胁迫,并直接影响着荒漠植物种子的活力和萌发[15-16]。较低的盐分对种子萌发影响不大,甚至有一定的促进作用,究其原因,可能是低盐条件能够促进细胞膜的渗透调节作用,也可能与微量的无机离子(如Na+)激活了某些酶有关[17,18]。本研究表明梭梭种子在盐分较高时,发芽率显著降低,复水后,在高盐分处理下的梭梭种子萌发恢复率显著提高,说明盐分抑制了梭梭种子萌发,但大部分梭梭种子仍具有萌发活力,当盐分被稀释时,即可完成萌发,表现出梭梭种子具有一定的耐盐性。
水分是荒漠地区植物种子萌发的限制条件,不同土壤含水量会导致荒漠木本植物种子萌发率和萌发速度有显著差异[19-21],如内蒙古吉兰泰地区梭梭在萌发期降水量减少的情况下,其种子萌发率显著降低,甚至不萌发[22]。本研究结果也是如此,在干旱胁迫达到一定程度时,梭梭种子表现出萌发速率缓慢,萌发率低甚至不萌发的现象,经过复水后,萌发率显著提高。这与盐胁迫萌发特征一致,不仅体现出梭梭种子具有一定的耐盐、耐旱性,也体现了荒漠植物在长期的进化中形成了其独特的种子萌发适应对策。当梭梭种子处于高盐分,较干旱的环境条件下时,绝大部分种子不萌发且仍具有萌发活力,当盐分被稀释、土壤含水量适宜时,种子即可完成萌发。这种适应策略,不仅提高了荒漠种子的高效利用率,而且种子萌发时间的不一致性,避免了因外界环境巨变造成种群灭绝的危险,从而使物种能够在恶劣环境中成功建群,这对实现治理荒漠化具有重要意义。
3.2 结论 萌发速率随萌发时温度的降低而减缓,各个温度下的梭梭种子萌发率均达到90%以上,其中适宜萌发温周期为20/15℃和15/10℃,低温仅仅抑制了梭梭种子的萌发速率,但并不影响种子的最终萌发率。(2)梭梭种子萌发率随着NaCl浓度的升高而降低,当盐浓度大于44g/L时,种子萌发率显著降低。经复水后,各个盐浓度下的种子萌发恢复率均达到60%以上,说明较高盐条件抑制了种子的萌发,但解除胁迫后,大部分梭梭种子仍具有萌发活力。(3)梭梭种子萌发率随着PEG浓度的升高而降低,当渗透势为-1.5MPa时,梭梭种子的萌发率显著降低至15%。渗透势达到-2.4MPa时,种子不萌发,复水后,种子的萌发率显著提高至70%以上,表明绝大部分梭梭种子经历干旱胁迫后,仍具有较高的萌发活力。
参考文献
[1]HuangZY,ZhangXS,ZhengGH,et al.Influence of light,temperature,salinity and storage on seed germination of Haloxylon ammodendron[J].Journal of Arid Environments,2003,55:453-464.
[2]Wang M,Zhang SX,and Chu GM.Point pattern analysis of different life stages of Haloxylon ammodendronin Desert-oasis ecotone of south Junggar basin[J].Polish Journal of Environmental Studies,2014,23(6):367-373.
[3]薛建国.水分、盐分和温度对几种荒漠植物种子萌发的影响[D].兰州:甘肃农业大学,2008.
[4]周平.温度、盐分及干旱胁迫对白梭梭种子萌发的影响[D].乌鲁木齐:新疆农业大学,2012.
[5]吕朝燕,张希明,吕薇,等.梭梭种子萌发对干旱和盐分胁迫的响应[J].北方园艺,2016(01):55-60.
[6]Khan MA,UngarIA.Germination of the salt tolerant shrub Suaeda fruticosa from Pakistan:salinity and temperature responses[J].Seed Science and Technology,1998,26:657-667.
[7]Tobe K,Li XM,and Omasa K.Effects of sodium chloride on seed germination and growth of two Chinese desert shrubs,Haloxylon ammodendron and H.persicum[J].Australian Journal of Botany,2000,48:455-460.
[8]韓建欣,魏岩,严成,等.高枝假木贼的胎生萌发特性及其生态适应[J].生态学报,2011,31(10):2662-2668.
[9]Shimono Y,and Kudo G.Comparisons of germination traits of alpine plants between fellfield and snowbed habitats[J].Ecological Research,2005,20:189-197.
[10]TLIG,TAHAR,GORAI.Germination responses of Diplotaxis harra to temperature and salinity[J].FLORA,2008,203(5):421-428.
[11]Jeltsch F,Moloney K A,Schurr F M,et al.The state of plant population modelling in light of environmental change[J].Perspectives in Plant Ecology,Evolution and Systematics,2008,9(3):171-189.
[12]Shevtsova A,Graae B J,Jochum T,et al.Critical periods for impact of climate warming on early seedling establishment in subarctic tundra[J].Global Change Biology,2009,15(11):2662-2680.
[13]赵晓英,任继周,王彦荣,等.3种锦鸡儿种子萌发对温度和水分的响应[J].西北植物学报,2005,25(2):211-217.
[14]黄振英,张新时,YITZCHAK,等.光照、温度和盐分对梭梭种子萌发的影响[J].植物生理学报,2001,27(3):275-280.
[15]安守芹,于卓,孔丽娟,等.花棒等四种豆科植物种子萌发及苗期耐盐性的研究[J].中国草地,1995(06):29-32.
[16]刘果厚,王树森,任侠.三种濒危植物种子萌发期抗盐性、抗旱性研究[J].内蒙古林学院学报,1999(01):33-38.
[17]王刚,梁学功,冯波.沙漠植物的更新生态位Ⅰ.油篙、柠条、花棒的种子萌发条件的研究[J].西北植物学报,1995(05):102-105.
[18]邢福,郭继勋,王艳红.狼毒种子萌发特性与种群更新机制的研究[J].应用生态学报,2003(11):1851-1854.
[19]Ren J,Tao L,Liu X M.Effect of water supply on seed germination of soil seed-bank in desert vegetation[J].Acta Botanica Sinica,2002,44(1):124-126.
[20]曾彦军,王彦荣,萨仁,等.几种旱生灌木种子萌发对干旱胁迫的响应[J].应用生态学报,2002(08):953-956.
[21]赵晓英,任继周,李延梅.黄土高原3种锦鸡儿种子萌发对温度的响应[J].应用与环境生物学报,2004(03):292-294.
[22]杨美霞,邹受益,赵学勇.吉兰泰地区梭梭林天然更新研究[J].內蒙古林学院学报,1995(02):74-86.
(责编:张宏民)
猜你喜欢
山东农业科学(2016年12期)2017-01-21
现代农业科技(2016年20期)2016-12-20
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年17期)2016-10-17
现代园艺(2016年2期)2016-03-15
