
Structure and dendroecology of Thuja occidentalis in disjunct stands south ofits contiguous range in the central Appalachian Mountains,USA
2017-04-18 05:15JoshuaKincaid
Forest Ecosystems 2017年1期
Joshua A.Kincaid
Structure and dendroecology of Thuja occidentalis in disjunct stands south ofits contiguous range in the central Appalachian Mountains,USA
Joshua A.Kincaid
Background:Information on foreststructure,growth,and disturbance history is essentialforeffective forest management in a dynamic landscape.Because most ofour research concerning the ecology and growth of Thuja occidentalis comes from sites in northern portions ofits range,highly contextualbiotic and abiotic factors that affect the species in more southern locales may notbe fully accounted for.This research characterized the structural attributes and growth dynamics of Thuja occidentalis in disjunctforeststands south ofits contiguous range margin.
Northern white cedar,Disturbance,Climate,Range,Disjunct,Radialgrowth,Regeneration
Background
Information on stand structure,growth,and disturbance history is essential to our understanding of patterns and processes in forest ecosystems.By examining the structural attributes and radial growth patterns of trees,we can quantitatively characterize conditions that have influenced forest stand development(Lorimer 1980;Frelich and Lorimer 1991;Hart and Grissino-Mayer 2008;Hart et al.2012).The range of variation elucidated by quantitative data facilitates more effective forest management in a landscape affected by both anthropogenic activities and climate change(Kincaid and Parker 2008;Hart et al.2010;Hart et al.2012).Climate change is expected to greatly reduce suitable habitat for many tree species in eastern North America.Climate coupled with factors such as disturbance and competition can affect the physiology and growth of trees,including processes such as regeneration and recruitment (Paul et al.2014).Under various climate change scenarios,one of the most impacted tree species will be northern white cedar(Thuja occidentalis L.)(Iverson et al.2008).
T.occidentalis,also known as arborvitae,is a shadetolerant coniferous tree species of eastern North America. This slowing growing tree commonly reaches heights of 12–15 m and diameters ranging from 30 to 60 cm(Johnston 1990).Although individuals exceeding 1000 years of age have been documented in Canada,the typicallifespan for T.occidentalis is 80–400 years(Larson 2001).The species tolerates a range of substrates,but growth is maximized on moist,well-drained soils derived from calcareous bedrock(Johnston 1990).T.occidentalis occurs in mixed to pure stands throughout its contiguous range, which extends across southeastern Canada and the northeastern United States.Isolated,disjunct stands exist beyond the contiguous range margin as far south as North Carolina and Tennessee(Braun 1950;Caplenor and Speir 1975;Walker 1987),but are rare in the central Appalachian Mountains(Fleming 1999;Fleming and Coulling 2001).
It is a common assumption that environmental conditions experienced by marginal or disjunct populations are more stressful or different than those experienced by core populations(Lesica and Allendorf 1995;Hardie and Hutchings 2010;Hart et al.2010).If marginal populations are adapted to more stressful or different biotic and abiotic conditions,they may have high conservation value as evolutionary legacies or components of biodiversity(Hoffmann and Blows 1994;Allendorf et al. 1997;Hardie and Hutchings 2010).Several studies have examined T.occidentalis forest structure and growth in northern portions of the range,including in disjunct populations(e.g.Kelly et al.1994;Tardif and Stevenson 2001;Kipfmueller et al.2010;Grotte et al.2012;Ruel et al.2014).However,research examining T.occidentalis forests south of their contiguous range margin has been limited to examinations of ecological community characteristics and population genetics(Walker 1987;Young 1996;Fleming 1999;Fleming and Coulling 2001).
The overall goal of this research was to document the structural attributes and radial growth dynamics of T.occidentalis in disjunct stands south of its contiguous range margin.There were three specific objectives for this research:1)to quantify stand structural attributes;2)to reconstruct the history of T.occidentalis growth-release events;and 3)to characterize the relationship between T. occidentals radialgrowth and climate.
Study area
This study was conducted in Rockingham County, Virginia(38.6363 N,−78.8502 W)located in the northern Shenandoah Valley,USA(Fig.1).Rockingham County lies within the Ridge and Valley Physiographic Province,which is characterized by long,linear ridges separated by valleys with a trellis drainage pattern.Bedrock consists of Paleozoic sedimentary rocks(Fenneman 1938).Mean total precipitation in the study area is 852 mm,61%of which occurs during the months of May through October. The mean annualtemperature is 13 °C(SERCC 2016).
Braun(1950)classified the study area as part of the Ridge and Valley Section of the Oak-Chestnut Forest Region,but the area is a transitional zone with elements of the mixed mesophytic and oak-chestnut regions. More recently the area was classified as the Appalachian Oak Section of the Mesophytic Forest Region with indicator species including Quercus alba,Quercus prinus, Liriodendron tulipifera and Pinus virginiana (Dyer 2006).T.occidentalis forest stands in the region are typically small(~0.1 to 1.0 ha),isolated,and extremely rare.In Virginia,T.occidentalis forests tend to be confined to north-facing slopes and cliffs with calcareous bedrock(Fleming 1999;Fleming and Coulling 2001). The disjunct T.occidentalis stands examined in this research are located approximately 440 km south of the contiguous range margin.
Methods
Structural attributes were obtained from 10 0.01 ha circular plots in each of two T.occidentalis forest stands(n=20 plots).Plot centroids were established every 10 m along a randomly placed transect in each stand.Each transect was situated at the mid-slope position and parallel to slope contours to avoid forest and environmental transitions.Two transects were adequate for data collection because both stands were oblong in extent and located on steep slopes.In each plot,species, diameter at breast height(dbh=1.4 m)of all stems ≥2.5 cm dbh,and snag abundance were recorded.Thepoint-quarter method was used at plot centroids to systematically select canopy trees for the collection of increment cores(Cottam and Curtis 1956).In some cases steep terrain restricted sampling to trees that could be safely cored at breast height.A total of 42 trees were examined in this research.
Fig.1 Map showing the range of Thuja occidentalis and study area(see Additionalfile 1 for high resolution)
In the laboratory,all increment cores were air-dried, glued to wood mounts,and sanded with progressively finer grits of sand paper(Phipps 1985;Stokes and Smiley 1996). The growth rings on all cores were visually inspected and crossdated using the list method(Yamaguchi 1991).Ringwidths were measured to the nearest 0.001 mm using a Velmex measuring stage and Measure J2X software.The visually crossdated measurement series were statistically verified using the computer program COFECHA(Holmes 1983;Grissino-Mayer 2001).
I used standard dendroecological techniques to reconstruct the history of T.occidentalis growth-release events in each stand(Lorimer and Frelich 1989;Nowacki and Abrams 1997;Rubino and McCarthy 2004).Raw tree-ring measurements were analyzed using a 10-year running median and percent-increase equation(Nowackiand Abrams 1997;Rubino and McCarthy 2004).Minor release events were defined as median growth increases between 25% and 49.9%.Major releases were defined as median growth increases of≥ 50%.The temporal distribution of growthrelease events detected using the ≥ 50%growth threshold was not significantly different(t=1.36;p=0.20)than the distribution observed using the absolute increase threshold(≥0.41 mm)of Fraver and White(2005).Only growth-release events sustained for a minimum of 3 years were recorded in this research(Rubino and McCarthy 2004).The growth-release data were used to construct a decadal disturbance chronology detailing the history of T. occidentalis growth-release events in each stand.
In preparation for the analysis of T.occidentalis growth-climate interactions,Iused the computer program ARSTAN to standardize the raw tree-ring measurements with a 30-year cubic smoothing spline.Standardization removes the effects of tree age and forest disturbance and allows for the comparison of trees with different growth rates(Cook 1985).Pearson correlation analysis was used to examine the relationship between climate and the T.occidentalis standard chronology.Climate data for Virginia Climate Division 5 were retrieved from the National Climate Data Center(NCDC 2015).Division 5 data were interpolated from stations using a 5 km grid resolution to ensure adequate spatial sampling.The climate variables examined in this research included monthly mean temperature,monthly total precipitation,and monthly Palmer Drought Severity Index(PDSI)for the current and previous year.The PDSI is commonly used in climate-tree growth research because it integrates measures of soil water,precipitation,and temperature,and more closely reflects the conditions required for tree growth than a single variable alone(Henderson and Grissino-Mayer 2009).The T.occidentalis radial growthclimate relationship was analyzed using a 17-month window extending from 1895–2010.
Results
Forest structuralattributes
The T.occidentalis forest stands contained a total of 13 tree species with a mean stand density of 915 ± 219.2 stems/ha(Table 1).Total stand basal area averaged 54.3 ± 12.4 m2/ha with T.occidentalis contributing substantially to the basal area of the sites.The importance values for T.occidentalis at CS1 and CS2 were 82.2% and 52.5%,respectively.Tilia americana comprised a distinct second tier at CS2 where the relative dominanceof the species was 21.7%.Species of tertiary importance at CS1 and CS2 included Liriodendron tulipifera, Fraxinus americana,Ailanthus altissima and Pinus strobus.

Table 1 Compositionalcharacteristics by forest stand
The size distribution of stems within the stands varied from unimodal or bimodal to multimodal structures (Fig.2).At CS1 the size distribution approximated a unimodal structure with the number of T.occidentalis stems peaking in the 20–25 cm size class.No T.occidentalis stems were present in the smallest size class.L.tulipifera, P.strobus,and the “other hardwood”group exhibited a bimodal structure of stems at CS1.F.americana occupied the intermediate size classes from 10–30 cm at both sites. The size distribution of T.americana was distinctly multimodal,while A.altissima exhibited a bimodal distribution of stems at CS2.T.occidentalis peaked in the largest size class at CS2,but was absent from the smallest class.The number of “other hardwood”stems was greatest in the smallest size classes atboth sites.
The absence of T.occidentalis stems in the smallest size class at both sites was reflected in the age distribution of the species.The number of T.occidentalis stems peaked in the 90–110 year age-classes,while stems less than 70 years old were absent from the stands.Aside from sapling snags,there was no evidence of recent T. occidentalis establishment and recruitment in the understories of CS1 or CS2(Fig.3).
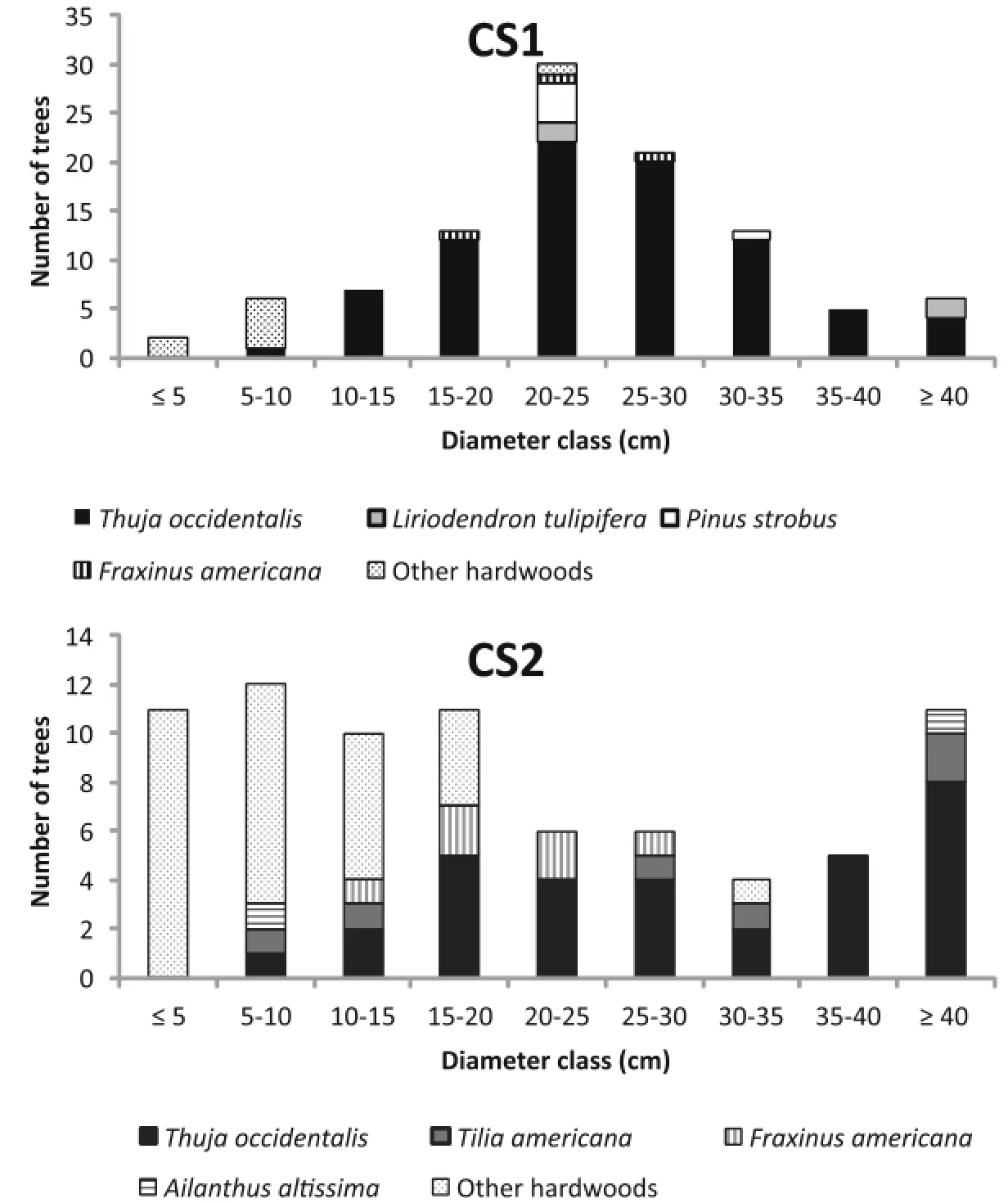
Fig.2 Diameter class distributions fortree species in each forest stand.Note that y-scale varies(see Additionalfile 1 for high resolution)
A total of 57 snags representing four different species were identified in this research(Table 2;Fig.4).Seven percent of the snags(n=4)were identified only as“hardwood.”Mean snag dbh for all stems was 11.3 ± 4.0 cm with a maximum of 36.5 cm for a single Tsuga canadensis stem.Total snag density averaged 280 ± 212.1 stems/ha within stands.The basal area of all snags was similar(~3.0 m2/ha)for both stands.T.occidentalis represented 71.9%of all snags,but T.canadensis snags were dominant(76.9%ofsnags)at CS2.
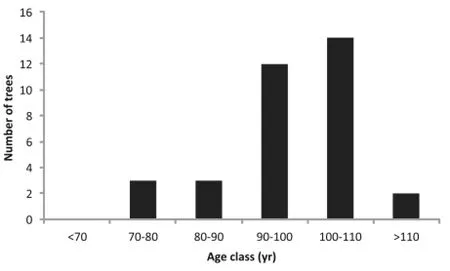
Fig.3 Age class distribution of Thuja occidentalis in the study area (see Additionalfile 1 for high resolution)
Radial growth-release events
Of the 42 T.occidentalis individuals examined in this research,59.5%(n=25)experienced growth release events (Fig.5).All25 of these trees exhibited at least one growth increase between 25%and 49.9%over the 10-year running median.Major events resulting in a ≥ 50%growth increase were detected in 28.6%(n=12)of the trees.One T.occidentalis individual experienced four minor releaseevents,while an additional five individuals experienced 2 to 3 minor growth release events.Only three individuals experienced a growth increase ≥ 100%over the 10-year running median,with the increase in one tree peaking at 165.3%.Growth release events were generally distributed across the chronology,but there was a conspicuous increase in major events during the 1950s and 1960s.The mean release duration was 7.1 ± 1.0 years.

Table 2 Snag characteristics by forest stand.Columns refer todensity(D,stems/ha)and basalarea(BA,m2/ha)

Fig.4 Diameter class distributions for snags in each forest stand(see Additionalfile 1 for high resolution)
Radial growth-climate interactions
The 42 T.occidentalis measurement series composing the chronology possessed an interseries correlation of 0.62(p < 0.01)and mean sensitivity of 0.25.Mean sensitivity is a measure of the year-to-year variability in tree-ring widths and is a standard COFECHA metric(Speer 2010). Sustained periods of above mean radial growth occurred during the 1940s and late 1960s to mid-1970s.Notable peaks in mean growth occurred in 1906 and 1995.Prolonged periods of below mean radial growth occurred in the late 1910s to mid-1920s and late 1940s to late 1950s.
Examination of the standard chronology revealed significant correlations(p < 0.05)between T.occidentalis radial growth and climate variables(Fig.6).The relationship between mean temperature and T.occidentalis growth was the weakest among allclimate variables examined in this research.Only previous August(+)and current July(−)mean temperature were significantly correlated with T.occidentalis growth.The relationship between T.occidentalis growth and precipitation was predominantly positive,with previous October,current June and August-September precipitation being significantly correlated with the chronology.The positive correlation between T.occidentalis growth and PDSI was generally stronger and more persistent than the association between growth and temperature or precipitation alone.
Discussion
Forest structures and dynamics
T.occidentalis was the leading canopy dominant at CS1 and CS2,but was not well-represented in the smallest size classes.The age distribution of T.occidentalis indicated a wave of establishment between 1900 and 1920, with successful recruitment lacking after 1940.Heitzman et al.(1997)documented a comparable timeline of T.occidentalis establishment and recruitment in Michigan, which they ascribed to stand initiating disturbances between 1870 and 1935,followed by stem exclusion in the1940s.The abundance of snags,indicative of self-thinning in the past,and the presence of hardwood species in the smallest size classes suggests CS1 and CS2 are in the early phases of the understory reinitiation stage during which herbs,shrubs,and advance regeneration invade the forest (Oliver and Larson 1996).As the understory reinitiation stage progresses,stand densities should decline to levels at which the regeneration of T.occidentalis is more likely to occur(Lamy et al.1999).
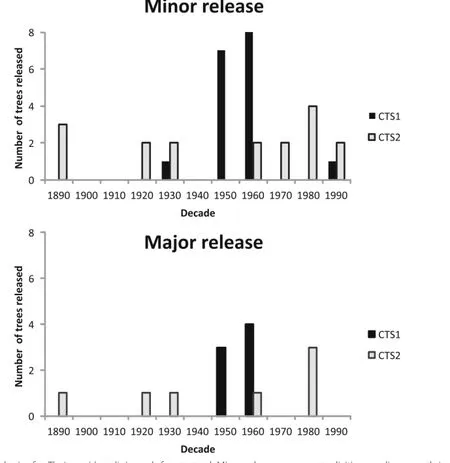
Fig.5 Disturbance chronologies for Thuja occidentalis in each forest stand.Minor releases are events eliciting median growth increases between 25 and 49.9%.Major releases are events eliciting median growth increases of≥ 50%(see Additionalfile 1 for high resolution)
The presence of A.altissima in the understory and overstory at CS2 is cause for concern.A.altissima is an introduced species from China that frequently spreads into forests by aggressively colonizing canopy gaps (Knapp and Canham 2000;Espenschied-Reilly and Runkle 2008;Martin et al.2010).In addition to outcompeting native species in canopy gaps,A.altissima is associated with changes in ecosystem processes and soil nutrient cycling,which may change the relative abundance of native trees to favor other,more competitive species(Gomez-Aparicio and Canham 2008).Using herbicides to control A.altissima is probably the most effective approach at this time,although landowners with limited budgets can remove the species with physical approaches such as pulling,cutting,digging,or girdling(Virginia 2009).
Disturbance and radialgrowth
T.occidentalis growth-release events were distributed across the disturbance chronology and generally affected a small number of trees,which is characteristic of forest gap dynamics.Canopy gap-scale processes involving the death of a tree or group of trees are often necessary for the persistence of shade tolerant tree species in forests (Runkle and Yetter 1987;Runkle 1998;Frelich 2002; Kincaid 2012).Increased light availability has been reported to enhance T.occidentalis seedling survival and abundance in northern portions of the range(Cornett et al.2000;Rooney et al.2002).Moreover,Scott and Murphy(1987)observed most T.occidentalis regeneration in their study area in Michigan was associated with a large patch of windthrown trees,not small,single-tree gaps. Despite evidence of ongoing canopy gap-scale processes within the forest stands,both structural attributes and field observations suggested a lack of successful T.occidentalis establishment during the last 70 years. This research was limited to the examination of radial growth-release events and stand structures,but I speculate single tree gaps are insufficient for T.occidentalis regeneration because adjacent canopy trees rapidly expand to occupy the space(Oliver and Larson 1996).
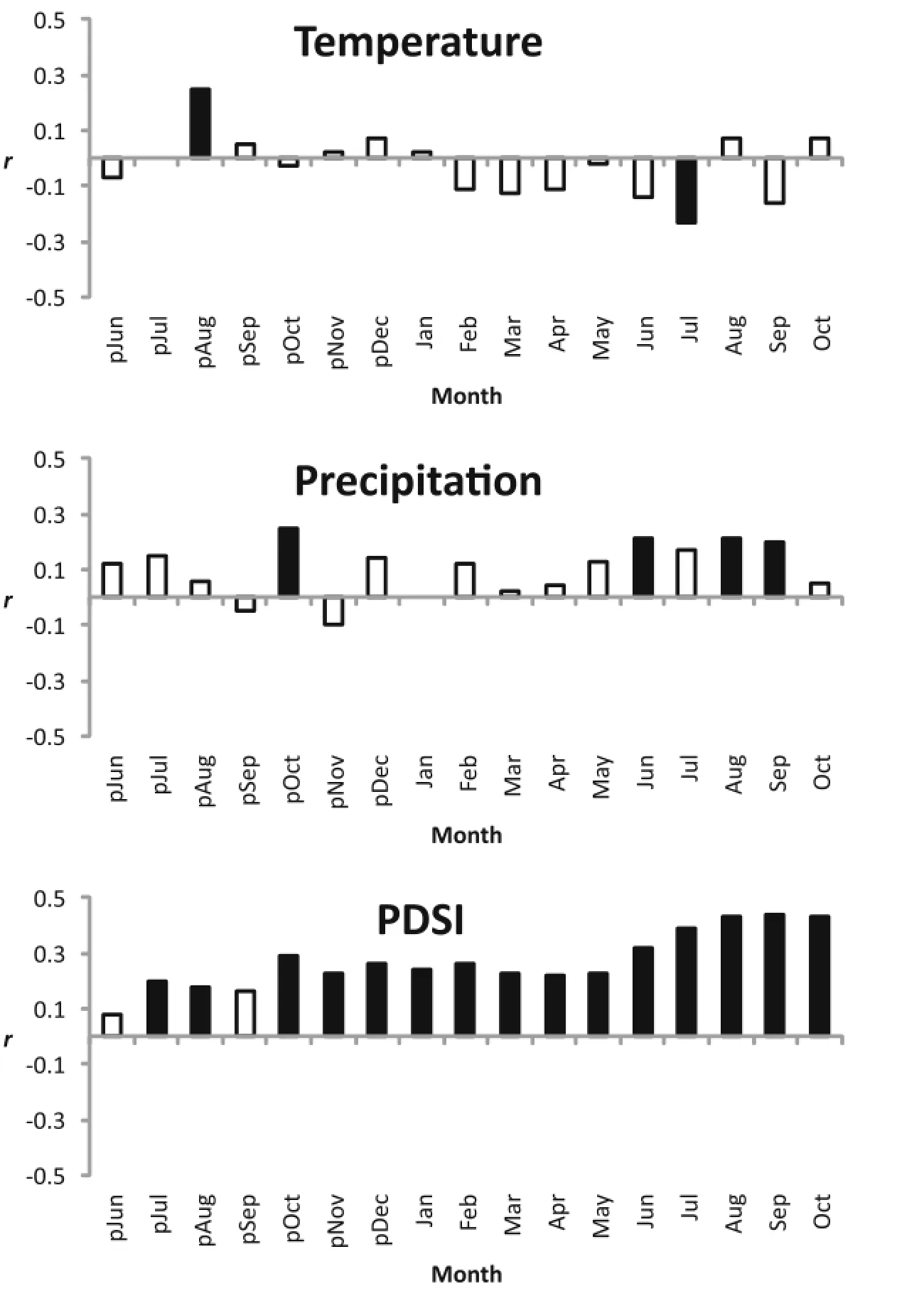
Fig.6 Correlation between the standardized tree-ring chronology and climate variables.Solid bars indicate significantvalues(p < 0.05)(see Additional file 1 forhigh resolution)
Climate and radialgrowth
The interseries correlation for all 42 T.occidentalis measurement series was statistically significant and compared well with the coefficient(0.66 versus 0.62,p < 0.01) reported by Kipfmueller et al.(2010)for a chronology developed in Minnesota,USA.Mean sensitivity of the T. occidentalis chronology also was within the range of variation reported for other chronologies developed in northern portions of the range(Kelly et al.1994;Tardif and Bergeron 1997;Tardifand Stevenson 2001;Kipfmueller et al.2010).The mean sensitivity of the chronology was nearly identical(0.25 versus 0.24)to the sensitivity of another disjunct shade tolerant conifer at its southern range margin in the Appalachian Mountains,USA(Hart et al. 2010).Although the sensitivity of T.occidentalis to climate is similar throughout its range,there appears to be geographicalvariation in the growth response of the species to climate factors such as temperature and precipitation,a phenomenon also observed in other tree species(Cook and Cole 1991;Hart et al.2010).
In northern portions of the range,T.occidentalis radial growth is inversely related to previous summer temperatures(Kelly et al.1994;Tardif and Bergeron 1997;Tardif and Stevenson 2001;Kipfmueller et al. 2010).The current study found a positive association between previous August mean temperature and T. occidentalis growth.The positive relationship between previous summer temperatures and growth has been observed in other conifers,including Chamaecyparis thyoides L.(Atlantic white cedar)(Hopton and Pederson 2003;Henderson and Grissino-Mayer 2009).Moderate temperatures and abundant precipitation late in the growing season may be favorable for T.occidentalis growth in the central Appalachian Mountains;however,the association between growth and temperature is not constant throughout the year.The temperature-growth association turns negative during the spring and summer of the current year,which is a period of phloem expansion in T. occidentalis(Bannan 1955).Phloem tissue is essential for the translocation of photosynthates from leaves to other parts of the tree.An inverse growth response to summer temperatures is common in trees because higher temperatures tend to facilitate internalwater deficits and increased respiration over carbon fixation(Edwards and Dixon 1995;Henderson and Grissino-Mayer 2009).Although some researchers have suggested T.occidentalis growth is not limited by climate,particularly by temperature in northern portions of the range,climate could influence the regeneration and growth of the species(Tardif and Stevenson 2001;Paul et al.2014)in disjunct stands south ofthe contiguous range margin.
In contrast to the relationship between T.occidentalis radial growth and temperature,the relationship between radial growth and moisture variables was more consistently positive.The positive associations between spring and summer precipitation of the current year and T.occidentalis growth are consistent with relationships reported in northern portions ofthe range,including the northwestern range margin(Tardif and Stevenson 2001;Kipfmueller et al.2010).The significant positive association between previous October precipitation and T.occidentalis growth is unique to this study.Hart et al.(2010)found a positive association between precipitation in October of the previous year and the growth of T.canadensis in disjunct stands near its southern range margin in the Appalachian Mountains.The authors speculated that October precipitation influences autumn phenology.It also is possible precipitation during the fall of the previous year facilitates ongoing carbohydrate storage,which is necessary for renewed growth in the following spring (Henderson and Grissino-Mayer 2009).The consistently positive association between T.occidentalis growth and PDSI has been observed in other southern conifers and is at least partly the result of autocorrelation in the index because values from previous months are used to calculate current values(Grissino-Mayer and Butler 1993;Henderson and Grissino-Mayer 2009;Hart et al.2010). Despite the autocorrelation in the index values,the positive relationship between PDSI and T.occidentalis growth may reflect the sensitivity of the species to moisture conditions in disjunct stands south of its contiguous range margin.The positive association between growth and PDSI is particularly strong from late summer to fall of the current year,which indicates that moisture conditions during this period are important to the growth of T.occidentalis in the study area.
Conclusion
This research has characterized the structural attributes and radial growth dynamics of disjunct T.occidentalis forests in the central Appalachian Mountains,USA. Structural attributes indicate the forest stands are in the understory reinitiation stage of forest development(Oliver and Larson 1996).Silvicultural manipulations such as thinning may be necessary to stimulate the understory and promote T.occidentalis establishment.It also may be necessary to utilize chemical and physical approaches to reduce A.altissima in order to prevent the species from colonizing canopy gaps and outcompeting native species such as T.occidentalis(Knapp and Canham 2000;Espenschied-Reilly and Runkle 2008;Virginia 2009;Martin et al.2010).
The sensitivity of T.occidentalis radial growth to climate appears similar throughout its range,including at disjunct sites beyond its contiguous range margin.However,geographical variation in the growth response of the species to climate factors is apparent(Cook and Cole 1991;Hart et al.2010).The response of T.occidentalis growth to moisture conditions is predominately positive throughout the year,while the relationship between growth and temperature is much more irregular.
The current study has documented geographical variation in the growth of T.occidentalis in disjunct forest stands south of its contiguous range margin.Most studies examining T.occidentalis ecology and growth come from northern portions of the range,so more research is necessary to increase the geographical and ecological scope of our knowledge,particularly at more southern and disjunct sites.Indeed,current research could be overlooking important local or regional biotic and abiotic factors that can significantly affect T.occidentalis ecology and growth.
Additionalfile
Additional file 1:Allfigures in high resolution.(ZIP 3.57 mb)
Acknowledgements
Iwould like to thank Gary Fleming and Wendy Hochstedlerfor information regarding the white cedarforests in this study.Ialso thank Angela Felicio, Cory Miller,and Kirby Talbertforassistance in the field and laboratory.This manuscript was improved by comments from Doug Enders and two anonymous reviewers.
Author’s contribution
Joshua Kincaid is the sole author ofthis manuscript.
Author’s information
Joshua Kincaid,Ph.D.Associate Professor of EnvironmentalStudies and Geography,Shenandoah University.Background:Ecologicalbiogeography, dendroecology,and human-environment relationships
Competing interests
There are no financialornon-financialcompeting interests with regard to this manuscript.
Received:12 August 2016 Accepted:1 November 2016
Allendorf FW,Bayles D,Bottom DL,Currens KP,Frissell CA,Hankin D,Lichatowich JA,Nehlsen W,Trotter PC,Williams TH(1997)Prioritizing pacific salmon stocks forconservation.Conserv Biol11:140–152
Bannan MW(1955)The vascular cambium and radialgrowth in Thuja occidentalis L.Can JBot 33:113–138
Braun EL(1950)Deciduous forests ofeastern North America.The Blakiston Company,Philadelphia
Caplenor D,Speir H(1975)Thuja occidentalis L.on the Eastern Highland Rim in Tennessee.J Tenn Acad Sci50:74–75
Cook ER(1985)A time-series analysis approach to tree-ring standardization.PhD Dissertation,University ofArizona
Cook ER,Cole J(1991)On predicting the response of forests in eastern North America to future climate change.Clim Change 19:271–282
Cornett MW,Frelich LE,Puettmann KJ,Reich PB(2000)Conservation implications ofbrowsing by Odocoileusvirginianus in remnantupland Thuja occidentalis forests.Biol Conserv 93:359–369
Cottam G,Curtis JT(1956)The use ofdistance measures in phytosociological sampling.Ecology 37:451–460
Dyer JM(2006)Revisiting the deciduous forests ofeastern North America. Bioscience 56:341–352
Edwards DR,Dixon MA(1995)Mechanisms ofdrought response in Thuja occidentalis L.I.Water stress conditioning and osmotic adjustment.Tree Physiol15:121–127
Espenschied-Reilly AL,Runkle JR(2008)Distribution and changes in abundance of Ailanthus altissima(Miller)Swingle in a southwest Ohio woodlot.Ohio J Sci108:16–22
Fenneman NM(1938)Physiography ofeastern United States.McGraw-HillBook Company,New York
Fleming GP(1999)Plant communities of limestone,dolomite,and other calcareous substrates in the George Washington and Jefferson national forests.Virginia DepartmentofConservation and Recreation,Division of NaturalHeritage,NaturalHeritage Technical Report 99–4,Virginia
Fleming GP,Coulling PP(2001)Ecologicalcommunities ofthe George Washington and Jefferson nationalforests.Preliminary classification and description ofvegetation types.Virginia Departmentof Conservation and Recreation,Division of NaturalHeritage,Natural Heritage Technicalreport 01–1,Virginia
Fraver S,White AS(2005)Identifying growth releases in dendrochronological studiesofforestdisturbance.Can JFor Res 35:1648–1656
Frelich LE,Lorimer CG(1991)Naturaldisturbance regimes in hemlock-hardwood forests of the upper Great Lakes region.Ecol Monogr 61:145–164
Frelich LE(2002)Forest dynamics and disturbance regimes.Cambridge University Press,Cambridge
Gomez-Aparicio L,Canham CD(2008)Neighborhood analyses ofthe allelopathic effects of the invasive tree Ailanthus altissima in temperate forests.JEcol 96:447–458
Grissino-Mayer HD(2001)Evaluating crossdating accuracy:a manualand tutorial for the computer program COFECHA.Tree-Ring Res 57:205–221
Grissino-Mayer HD,Butler DR(1993)Effects ofclimate on growth ofshortleaf pine(Pinus echinata Mill.)in northern Georgia:A dendroclimatic study. Southeast Geogr 33:65–81
Grotte KL,Heinrichs DK,Tardif JC(2012)Old-growth characteristics of disjunct Thuja occidentalis standsat their northwestern distribution limit,central Canada.NatArea J32:270–282
Hardie DC,Hutchings JA(2010)Evolutionary ecology at the extremes of species’ranges.Environ Rev 18:1–20
Hart JL,Grissino-Mayer HD(2008)Vegetation patterns and dendroecology of a mixed hardwood foreston the Cumberland Plateau:Implicationsforstand development.For Ecol Manag 255:1960–1975
Hart JL,van de GevelSL,Sakulich J,Grissino-Mayer HD(2010)Influence of climate and disturbance on the growth of Tsuga canadensis at its southern limit in eastern North America.Trees 24:621–633
Hart JL,Clark SL,Torreano SJ,Buchanan ML(2012)Composition,structure,and dendroecology ofan old growth Quercus forest on the tablelands ofthe Cumberland Plateau,USA.For Ecol Manag 266:11–24
Heitzman E,Pregitzer KS,Miller RO(1997)Origin and early development ofnorthern white-cedarstands in northern Michigan.Can JFor Res 27:1953–1961
Henderson JP,Grissino-Mayer HD(2009)Climate-tree growth relationships of longleafpine(Pinus palustris Mill.)in the southeastern Coastal Plain,USA. Dendrochronologia 27:31–43
Hoffmann AA,Blows MW(1994)Species borders:Ecologicaland evolutionary perspectives.Trends EcolEvol9:223–227
Holmes RL(1983)A computer-assisted quality controlin tree-ring dating and measurement.Tree-Ring Bull43:69–78
Hopton HM,Pederson N(2003)Climate sensitivity of Atlantic White Cedarat its northern range limit.In:Burke MK,Sheridan P(eds)Atlantic White Cedar: ecology,restoration,and management.Proceedings ofthe Arlington Echo Symposium,Millersville
Iverson LR,Prasad AM,Matthews SN,Peters M(2008)Estimating potentialhabitat for 134 eastern UStree species undersix climate scenarios.For EcolManage 254:390–406
Johnston WF(1990)Thuja occidentalis L.Northern White Cedar.In:Burns RM, Honkala BH(eds)Silvics of North America:Volume 1.AgriculturalHandbook 654.USDA Forest Service,Washington DC
Kelly PE,Cook ER,Larson DW(1994)A 1397-year tree-ring chronology of Thuja occidentalis from clifffaces of the Niagara Escarpment,southern Ontario, Canada.Can JFor Res 24:1049–1057
Kincaid JA(2012)Regeneration dynamics of Tsuga canadensis in the southern Appalachian Mountains.USA 26:1045–1056
Kincaid JA,ParkerAJ(2008)Structuralcharacteristics and canopy dynamics of Tsuga canadensis in forests ofthe southern Appalachian Mountains,USA. Plant Ecol199:265–280
Kipfmueller KF,Elliott GP,Larson ER,Salzer MW(2010)An assessment ofthe dendroclimatic potentialofthree coniferspecies in northern Minnesota. Tree-Ring Res 66:113–126
Knapp LB,Canham CD(2000)Invasion of an old-growth forest in New York by Ailanthus altissima:sapling growth and recruitmentin canopy gaps.JTorrey Bot Soc 127:307–315
Lamy S,Bouchard A,Simon JP(1999)Genetic structure,variability,and mating system in eastern white cedar(Thuja occidentalis)populations ofrecentorigin in an agriculturallandscape in southern Quebec.Can JFor Res 29:1383–1392
Larson DW(2001)The paradox ofgreatlongevity in a short-lived tree species. Exp Gerontol36:651–673
Lesica P,Allendorf FW(1995)When are peripheralpopulations valuable for conservation?Conserv Biol9:753–760
Lorimer CG(1980)Age structure and disturbance history ofa southern Appalachian virgin forest.Ecology 61:1169–1184
Lorimer CG,Frelich LE(1989)A methodology forestimating canopy disturbance frequency and intensity in dense temperate forests.Can J For Res 19:651–663
Martin P,Canham CD,Kobe RK(2010)Divergence from the growth-survival trade-offand extreme high growth rates drive patterns of exotic tree invasions in closed-canopy forests.JEcol98:778–789
NCDC(2015)National Climatic Data Center.US Department of Commerce, National Oceanic and Atmospheric Administration,Asheville,http://www. ncdc.noaa.gov.Accessed 15 Jun 2015
NowackiGJ,Abrams MD(1997)Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks.Ecol Monogr67:225–249
Oliver CD,Larson BC(1996)Forest stand dynamics.John Wiley&Sons,New York
Paul V,Bergeron Y,Tremblay F(2014)Does climate controlthe northern range limit ofeastern whitecedar(Thuja occidentalis L.)?Plant Ecol215:181–194
Phipps RL(1985)Collecting,preparing,crossdating,and measuring tree increment cores.US GeologicalSurvey Water Resources Investigations Report85–4148
Rooney TP,Solheim SL,Waller DM(2002)Factors affecting the regeneration of northern white cedarin lowland of forests ofthe Upper Great Lakes region, USA.ForEcolManag 163:119–130
Rubino DL,McCarthy BC(2004)Comparative analysis of dendroecological methods used to assess disturbance events.Dendrochronologia 21:97–115
RuelJC,Lussier JM,Morissette S,Ricodeau N(2014)Growth response ofnorthern white cedar(Thuja occidentalis)to naturaldisturbances and partialcuts in mixedwood stands in Quebec,Canada.Forests 5:1194–1211
Runkle JR(1998)Changes in southern Appalachian canopy trees sampled thrice. Ecology 79:1768–1780
Runkle JR,Yetter TC(1987)Treefalls revisited:gap dynamics in the southern Appalachians.Ecology 68:417–424
Scott ML,Murphy PG(1987)Regeneration patterns ofnorthern white-cedar,an old-growth forest dominant.Am MidlNat 117:10–16
SERCC(2016)The Southeast RegionalClimate Center.The University ofNorth Carolina,Chapel Hill,http://www.sercc.com/climate.Accessed 10 Jul2016
Speer JH(2010)Fundamentals oftree-ring research.University ofArizona Press,Tuscon
Stokes MA,Smiley TL(1996)An introduction to tree-ring dating.University of Arizona Press,Tucson
Tardif J,Bergeron Y(1997)Comparative dendroclimatologicalanalysis oftwo black ash and two white cedar populations from contrasting sites in the Lake Duparquet region,northwestern Quebec.Can J For Res 27:108–116
Tardif J,Stevenson D(2001)Radialgrowth-climate association of Thuja occidentalis L.at the northwestern limit of its distribution,Manitoba,Canada. Dendrochronologia 19:179–187
Virginia DOF(2009)Controland utilization oftree-of-heaven.Virginia Department of Forestry,Charlottesville,Available via:www.dof.virginia.gov/infopubs/ Control-And-Utilization-of-TOH-2009-05_pub.pdf
Walker GL(1987)Ecology and population biology of Thuja occidentalis in its southern disjunct range.PhD Dissertation,University of Tennessee
YamaguchiDK(1991)A simple method forcross-dating incrementcores from living trees.Can J For Res 21:414–416
Young J(1996)The cliffecology and genetic structure ofnorthern white cedar (Thuja occidentalis L.)in its southern disjunct range.MS Thesis,University of Tennessee
Correspondence:jkincaid@su.edu
Shenandoah University,1460 University Drive,Winchester,VA 22601,USA
Methods:The Thuja occidentalis forests examined in this research were located in the centralAppalachian Mountains, USA,approximately 440 km south ofthe contiguous range margin ofthe species.Foreststructuralattributes were characterized in two Thuja occidentalis forest stands,which are rare in the region.Tree-ring chronologies were used to examine the influences ofdisturbance and climate on the growth of Thuja occidentalis.
Results:The forests contained a totalof13 tree species with Thuja occidentalis contributing substantially to the basal area ofthe sites.Thuja occidentalis stems were absent in the smallest size class,while hardwood species were abundantin the smallest classes.Thuja occidentalis stems also were absentfrom the < 70 years age class. By contrast,Thuja occidentalis snags were abundant within stands.Growth-release events were distributed across the disturbance chronology and generally affected a smallnumberoftrees.The Thuja occidentalis tree-ring chronology possessed an interseries correlation of0.62 and mean sensitivity of0.25.The correlation between mean temperature and Thuja occidentalis growth was weak and variable.Growth and moisture variables were more strongly correlated,and this relationship was predominantly positive.
Conclusions:Structuralattributes indicate the forests are in the understory reinitiation stage offorest development. Silviculturalmanipulations may be necessary to promote Thuja occidentalis establishment.The sensitivity of Thuja occidentalis to climate appears similarthroughout its range,butgeographicalvariation in the growth response to climate factors is apparent.More research is necessary to expand the geographicaland ecologicalscope ofour knowledge concerning Thuja occidentalis,particularly at more southern and disjunct sites.

- Forest Ecosystems的其它文章
- Romanian legalmanagement rules limit wood production in Norway spruce and beech forests
- Life cycle cost and economic assessment of biochar-based bioenergy production and biochar land application in Northwestern Ontario,Canada
- Adaptation of forest management to climate change as perceived by forest owners and managers in Belgium
- Patterns offorest composition and their long term environmentaldrivers in the tropicaldry forest transition zone of southern Africa
- Drought sensitivity ofbeech on a shallow chalk soilin northeastern Germany – a comparative study
- Development and evaluation ofan individualtree growth and yield modelfor the mixed species forest ofthe Adirondacks Region of New York,USA