
光温条件和土壤湿度对栓皮栎幼苗蒸腾潜热和叶温的影响
2017-04-18 07:09陈景玲王佩舒刘琳奇王谦王旭杨喜田张劲松
中国水土保持科学 2017年1期
陈景玲,王佩舒,刘琳奇,王谦†,王旭,杨喜田,张劲松
(1.河南农业大学林学院,450002,郑州;2.中国林业科学院林业研究所,100091,北京)
光温条件和土壤湿度对栓皮栎幼苗蒸腾潜热和叶温的影响
陈景玲1,王佩舒1,刘琳奇1,王谦1†,王旭1,杨喜田1,张劲松2
(1.河南农业大学林学院,450002,郑州;2.中国林业科学院林业研究所,100091,北京)
研究光温条件和土壤湿度对栓皮栎幼苗叶片蒸腾的影响程度,以及太阳辐射、蒸腾和对流换热对叶温形成的贡献。用盆栽遮雨和称量法,控制土壤干旱胁迫水平(体积含水量)为轻度(12.5%~14.5%)、中度(9.5%~11.5%)和重度(5.5%~7.5%),并分别在自然和人工气候箱(温度控制在25~43℃)的环境下测定蒸腾速率和气象因子;用热量分析方法,定量确定各因子对叶温差的贡献。结果表明:1)轻度干旱下,蒸腾速率与正常土壤水分下相近,重度干旱胁迫下蒸腾速率降到1.5 mmol/(m2·s)以下;2)晴天的蒸腾速率与太阳辐射关系密切,呈正相关,不同土壤水分胁迫下的斜率不同,表明太阳辐射是蒸腾的主导因子;3)多云天时,蒸腾速率与太阳辐射的线性关系明显下降,说明白天蒸腾一旦开始,蒸腾速率不因短时间的太阳辐射下降而降低;4)在3个土壤干旱水平下,气温都不是蒸腾的主导因子。人工气候箱试验条件下,蒸腾速率虽与气温线性关系明显,但蒸腾速率明显小于晴天自然条件下。同时也说明,轻度干旱不影响栓皮栎蒸腾;在静风条件下,太阳辐射是栓皮栎叶温变化的主导因子,可使叶温变化7℃左右,占叶温变化的50%~70%;蒸腾潜热和对流换热项可使叶温变化1~2℃左右,各占叶温变化的10%~20%。本研究为构建栓皮栎的WSI,以及用叶气温差诊断栓皮栎土壤水分提供理论依据。
栓皮栎;辐射;温度;土壤水分;蒸腾速率;叶温
栓皮栎(Quercus variabilis)是干旱半干旱地区的适生乔木树种,蒸腾速率较低[1],作为退化生境的主要建群乔木树种,能维持稳定的乔灌草生态系统,又有保持水土的功能,在我国水土保持植物措施的面积比例高达80%[2],因而栓皮栎的应用价值和潜力较大;但是,过度的土壤干旱胁迫,造成栓皮栎生长势降低,严重的会枯死,使栓皮栎林失去稳定和丧失水土保持功能。栓皮栎实生苗也会因干旱而死亡,造成造林成活率低[3]。土壤水对植物的胁迫程度可在植物叶温上反映出来,叶气温差被用来计算水分胁迫指数(WSI),作为土壤水分诊断指标,这为生态系统健康状态判断提供有效途径;但这些研究集中在农业领域,称为作物水分胁迫指数(CWSI)[4-7],对栓皮栎叶温研究很少,更没有栓皮栎水分诊断研究。而栓皮栎的蒸腾速率远小于农作物,其叶气温差远大于农作物[8],因此,栓皮栎的CWSI也必然与农作物不同。
叶气温差法用于水分诊断的理论基础是冠层温度的形成。影响叶气温差的因子很多,如空气湿度、饱和差、气温、风速、太阳辐射、冠层净辐射和土壤含水量,这些因子大多也影响蒸腾速率,进而影响叶温形成[9,10]。多因子复合影响使单因子分析时的线性关系拟合结果较差[11],主要问题是对叶温形成的影响因子没有区别其重要性,影响叶气温差进行土壤水分诊断。前人综述了冠气温差等多种水分诊断方法,认为均存在一定问题,必须加强基础理论研究[12]。
笔者拟通过自然环境和人工气候箱控制环境因子试验,研究光、温和土壤水对栓皮栎叶片蒸腾速率的影响程度,以及太阳辐射、蒸腾和对流换热对叶温形成的贡献,为构建栓皮栎的WSI,以及用叶气温差诊断栓皮栎土壤水分提供理论依据。
1 试验设计与方法
1.1 试验材料
试验地点选在河南农业大学林学院试验基地(E 112°42',N 34°43')。试验材料为栓皮栎盆栽幼苗,共选择100盆试验用苗。2014年10月播种,每盆播3粒种子,2015年3月出苗,2016年4月开始试验,试验时苗龄13个月,平均苗高30 cm。试验用盆的高度为24 cm、上口径18 cm、盆底直径10 cm,花盆平均质量102.1 g。所用土壤为沙壤褐土,盆中土壤深度18 cm,盆内体积1 748.4 cm3,土壤密度1.22 g/cm3,土壤孔隙度52.9%,土壤田间持水量为30.0%(体积含水率)。
蒸腾速率测定采用Li-6400光合仪,太阳辐射强度、空气温度等气象因子用北京雨根公司的小气候监测系统测定,辐射和空气温湿度传感器为美国Avalon公司生产,环境控制采用杭州赛福强冷光源人工气候箱,称重采用感量0.001 g电子天平。
1.2 试验处理
每盆选择一株达平均株高的健壮栓皮栎幼苗,其他多余苗株紧贴盆土剪去,保留其成熟健康叶片5片,剪去其他叶片。
土壤湿度(体积含水量W)的设置参考文献[13],确定3个处理,即轻度干旱(W1)为12.5%~14.5%,中度干旱(W2)为9.5%~11.5%,重度干旱(W3)为5.5%~7.5%。从2016年4月10日开始,对选出的100盆幼苗,分3批浇水处理,时间间隔为3 d。傍晚时分浇水,水层到花盆上口径边缘,花盆放置一夜,待水分渗漏停止,保证土壤水分饱和,第二天早晨称量,取得花盆平均质量为2 950 g。按此平均花盆质量去土或补土,使花盆质量一致。去土或补土后,花盆中的土壤深度仍控制为18 cm,以保证花盆中土壤体积一致。处理后的幼苗搬进试验田防雨棚,依靠自然蒸发,将盆内土壤湿度下降到试验设计要求的范围。
控制土壤湿度采用称量法。于开始试验前,在经处理的100盆苗中,随机选10盆,依次编号,顺序分离出土壤、栓皮栎苗和空花盆,称量每个花盆的幼苗、空花盆质量,得出每株花盆苗质量均值mplant为31.9 g,空花盆质量均值mpot为102.1 g。将分离出的湿土,放进烘箱于105℃烘干,得出每花盆干土质量均值msoil为2 135.0 g。空花盆装水至18 cm高度,称出水质量,为土壤容积对应水质量均值mwater为1748.4 g。计算不同干旱程度下对应的试验花盆质量mtotal的公式为
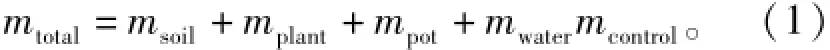
式中:Mcontrol为不同干旱程度的土壤容积含水量。由此得出,不同干旱程度的花盆质量范围(表1)。

表1 不同干旱程度下花盆控制质量Tab.1 Mass of flowerpot under different droughts
试验开始后,每天傍晚称花盆质量,对土壤水分低于各预定干旱水平的花盆,停止用于试验,而从前述3批浇水的花盆中,选择处于该干旱水平的花盆继续试验。对停止用于试验的花盆,重新补水至饱和,再放入防雨棚内自然蒸发,降低至控制土壤湿度时,重复用于试验。考虑到干旱胁迫对幼苗的损伤,中度和重度干旱的花盆不再重复用于试验。补水时,花盆下方放置托盘,收集因补水而流出的土壤,并放回花盆内。
1.3 测定方法
1.3.1 日变化的测定选自然蒸发后土壤含水量降至不同干旱水平范围的幼苗各3盆,用塑料袋包裹盆,只露出苗木,防止土壤蒸发和水分逸散。用Li-6400光合仪测定叶片温度和蒸腾速率,每盆取1片叶,重复测定3次。选典型晴天和多云天,从08:00开始,到18:00结束,1 h测定1次。
1.3.2 气候箱内栓皮栎蒸腾速率测定气候箱控制温度分别为25、27、29、31、33、35、37、39、41和43℃。将气候箱LED光源全打开,实测光强为500 W/m2,该光强为晴天自然光太阳辐射强度(0~1 000 W/m2)的中值,即光强达中等水平。设置空气湿度为40%(参照试验期间室外空气湿度日均值),使人工气候箱试验主要反映温度单一因素对蒸腾速率的影响。
25和43℃预实验表明,栓皮栎幼苗首次放入气候箱中3 h后,蒸腾速率才能达到稳定。选自然蒸发后,土壤水分降至表1中不同干旱水平的幼苗各3盆,用塑料袋包裹盆,只露出苗木。试验前对花盆称其质量,并记录为m0,将其放入设定的气候箱中适应3 h,在蒸腾速率稳定后,用Li-6400光合仪迅速测定栓皮栎幼苗蒸腾速率,每盆取1片叶片,重复测定3次。测定结束后,对花盆再次称其质量记为m1。保证试验前后花盆质量平均值(m0+m1)/2在干旱水平控制花盆质量范围内。然后,调整人工气候箱至下一个温度处理,1 h后进行蒸腾速率测定。如某一温度处理时,因蒸发使花盆质量低于控制质量范围的低值,则更换花盆,重复上述试验过程,以进行不同温度处理。更换的花盆首次放入气候箱,均需要稳定3 h的处理。
1.4 数据分析
气候学中,下垫面的热量收支平衡方程为
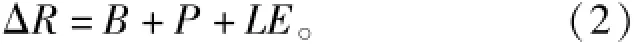
式中:ΔR为净辐射;B是土壤热通量;P是湍流热交换;L为水的蒸发潜热,L=2 498.9-2.33Tα,J/g;E为蒸腾速率,mmol/(m2·s)。
叶表面的净辐射为

式中:Qa为叶片吸收太阳辐射,W/m2;εl为叶片的辐射出射率;εe为天空的辐射出射率;σ为斯忒芬-波尔兹曼常数;Tl为叶温;Te为天空温度,Te= 1.06Ta-21;Ta为气温。Qa=R(1-α),R为太阳辐射强度,W/m2;α是叶表面对太阳辐射的反射率。
对叶片,其下面接触的是空气,不是土壤,即没土壤热通量B,而是仅与空气进行湍流热交换。在静风条件下,叶片上下表面的湍流热交换可近似看作相等,湍流热交换为

式中:hc为对流换热系数。静风时hc可参考D.M. Gates[14]和刘静等[15]的对流换热系数研究,引用公式hc=2.38(Tl-Ta)0.25,则各热量收支项对叶温的影响分析,用叶表面热量平衡公式计算:
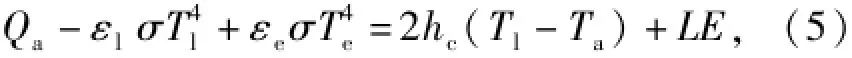
则净辐射对叶温变化的贡献为
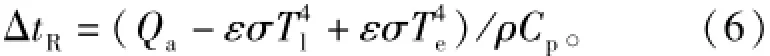
对流产生的叶温变化贡献为

潜热量产生的叶温变化贡献为

式中:ρ为栓皮栎叶片的表面密度,实测为0.015 85 kg/m2;Cp为叶片热容量,据文献[16]取3 590.1 J/(kg·℃)。
2 结果与分析
2.1 不同土壤干旱下的蒸腾速率
2.1.1 蒸腾速率与太阳辐射的关系在4月29日晴天条件下,蒸腾速率与太阳辐射的关系见图1。由图1a可知,蒸腾速率和太阳辐射的时间变化,3种土壤水分处理都是单峰型变化,峰值出现时间一致,且与太阳辐射峰值时间接近,仅略晚于太阳辐射峰值出现时间。在4月30日多云天(图略),太阳辐射因在13时出现低值,但蒸腾速率并不表现为随之降低。由图1b可知,不同土壤水分处理的蒸腾速率都随太阳辐射增加而增加,但从V1到V3,蒸腾速率随太阳辐射增加的幅度(线性关系式的斜率),及线性关系式的确定系数R2均在减小。
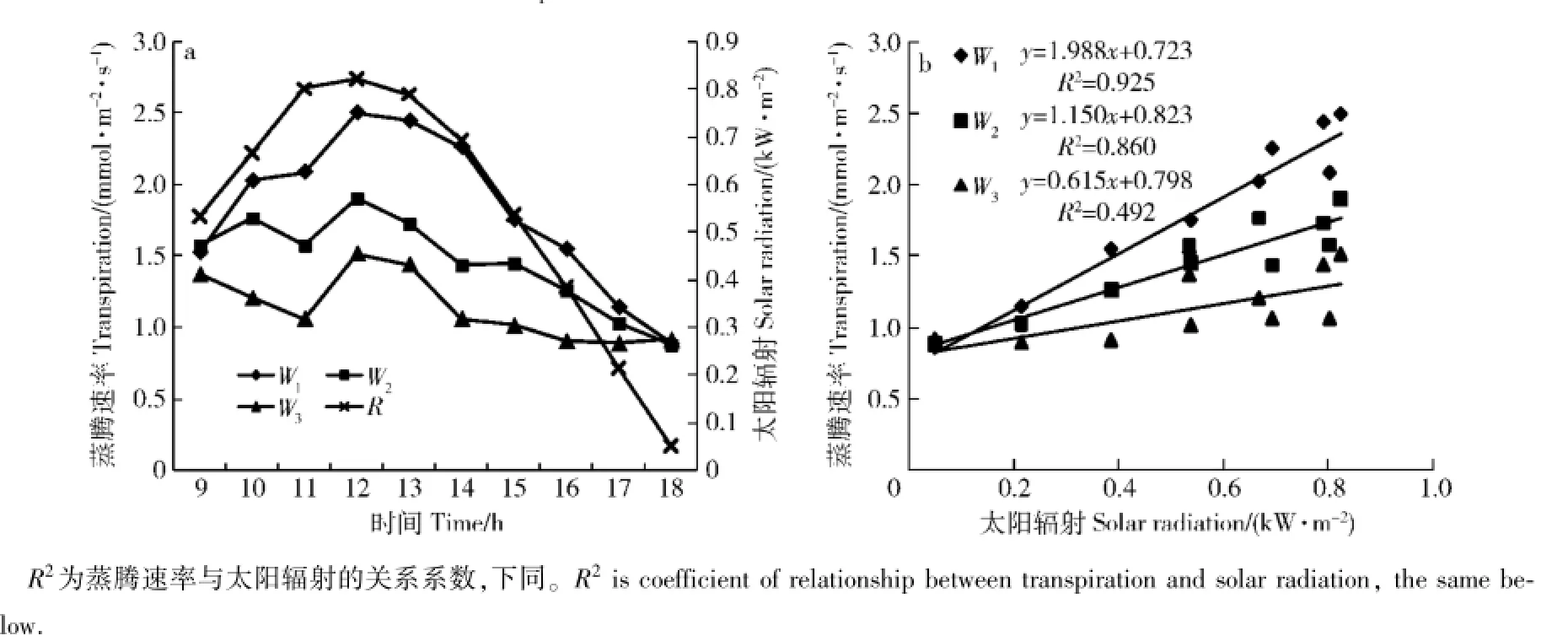
图1 蒸腾速率与太阳辐射关系(晴天)Fig.1 Relationship between transpiration(Tr)and solar radiation(R)of Quercus variabilis(In clear day)
对蒸腾速率与太阳辐射做回归分析显著性检验,结果见表2。其中:晴天轻度干旱时的确定系数为0.925 2,即太阳辐射因子能解释蒸腾速率变差的92.5%;确定系数随干旱胁迫增加而变小,中度干旱时为0.860 4,重度干旱时为0.492 0。在多云天,轻、中和重度干旱时的太阳辐射能解释蒸腾速率变化的46.3%、34.7%和20.3%。对比晴天与多云,其相同点是蒸腾速率都与太阳辐射正相关,说明太阳辐射是蒸腾速率的主导因子;不同点是多云天的相关显著性降低,主要原因是多云天的太阳辐射,在中午因多云而出现降低,但栓皮栎的蒸腾速率并没有随太阳辐射的短时间降低而降低;蒸腾速率与太阳辐射的升降有一定的节律一致,但蒸腾速率的波动比太阳辐射的波动小,且滞后。
2.1.2 蒸腾速率与气温的关系蒸腾与气温的关系均表现为不密切,晴天情况示于图2(多云天略)。从日变化图上看,在午后气温尚维持较高水平时,蒸腾速率已有大幅度下降,二者峰值的位相明显不同。线性回归关系式的确定系数均很低。说明气温与太阳辐射共同影响蒸腾速率时,气温不是主导因子。
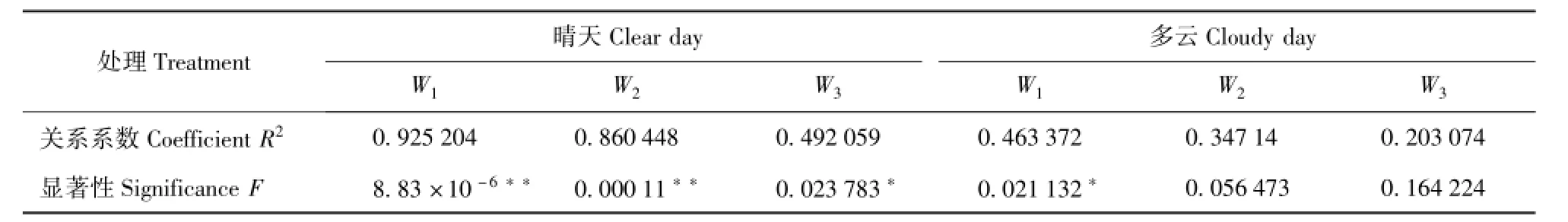
表2 不同土壤水处理下蒸腾速率与太阳辐射回归显著性检验Tab.2 Significence test of the relationship of transpiration and solar radiation under different soil moistrures
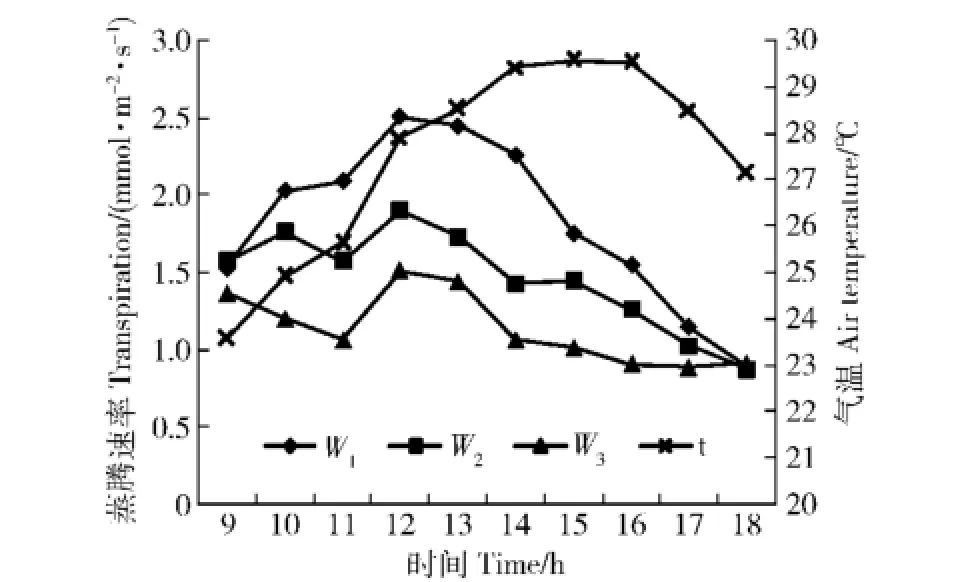
图2 蒸腾速率与气温关系(晴天)Fig.2 Relationship between transpiration and air temperature ofQuercus variabilis(In clear day)
2.2 人工气候箱控制温度对蒸腾速率的影响
为表现温度单因子对蒸腾速率的影响,在人工气候箱控制不同温度,对栓皮栎幼苗进行处理,结果见图3。可以看出:随温度增加,轻度、中度和重度干旱处理的蒸腾速率均逐渐增加;但蒸腾速率的变化率明显低于晴天自然环境下的数值,在轻度干旱条件下,只有0.6~1.4 mmol/(m2·s),最大值不超过1.5 mmol/(m2·s)。表现出温度单因子对蒸腾速率的作用较小,这与前述气温不是蒸腾主导因子的结论并不矛盾。
在干旱条件下,蒸腾速率总体很低,虽然重度干旱条件下也有随温度增加而增加的线性规律,且回归关系式可通过0.01信度的显著性检验(表3);但它是低蒸腾速率水平下的规律,最大值仅为0.6 mmol/(m2·s)。

表3 人工气候箱不同土壤水蒸腾速率与温度回归显著性检验Tab.3 Significence test of the relationship of transpiration and air temperature under different soil moistrures in manu-al climate box
2.3 蒸腾对叶温的影响
以轻度干旱处理W1的蒸腾数据,计算能量收支各分量,并进行能量分配比例和对叶温的贡献分析,按式(5)计算对流换热、净辐射和蒸腾量LE,得到各热量收支的分配。其随时间的变化见图4,净辐射ΔR随时间有明显的峰值特征,对流换热Φc和潜热交换量LE则数据较小。三者之和与吸收太阳辐射量Qa比较,时间变化曲线接近,中午和午前几乎重合,下午3个分量之和低于吸收太阳辐射量。二者重合说明各分量计算准确,下午二者有差异,说明存在计算误差;但由于差异不大,可以用这些分量分析各因子的作用规律。

图3 气候箱内栓皮栎蒸腾速率与气温关系Fig.3 Relationship between transpiration and air temperature of Quercus variabilis in artificial chamber
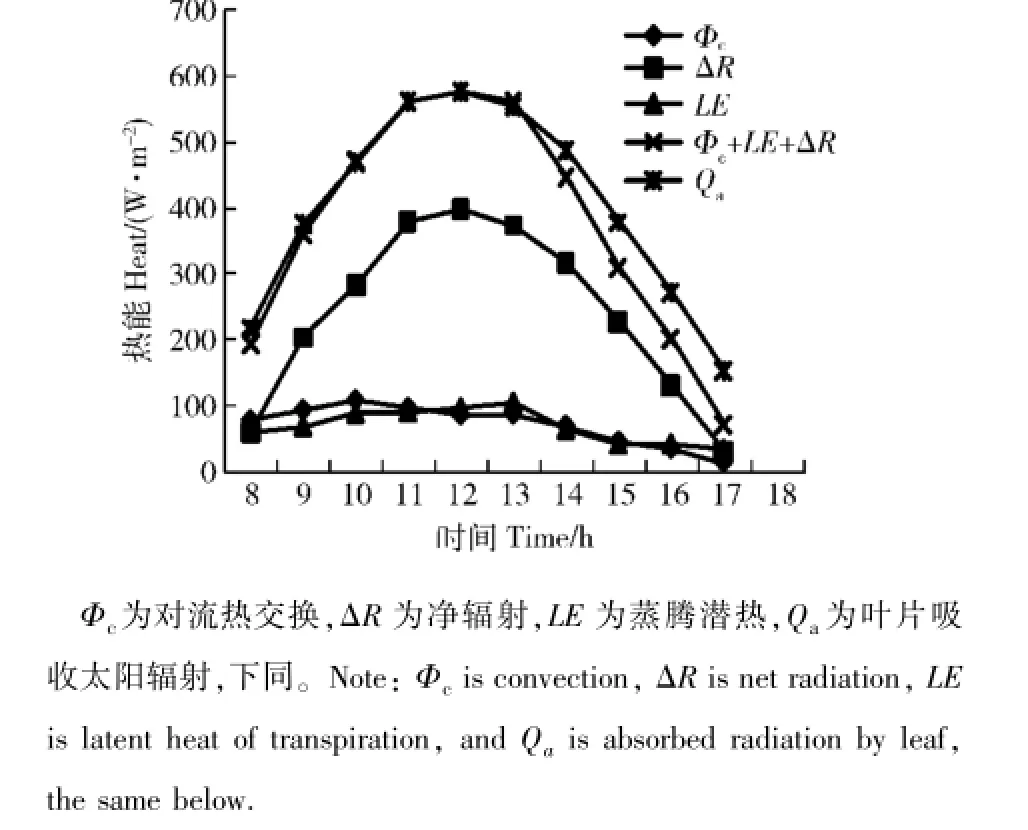
图4 叶表面各能量分支的比较Fig.4 Contrast of every energy item on leaf surface
分析能量收支各分量与吸收太阳辐射量Qa的比值可以看出,净辐射占比例最大,除去08:00和17:00外,整个白天所占比例均较高,在50%~70%之间。对流和潜热占比例相当,占10%~20%之间,说明能量支出所占比值较小。产生的叶温变化量见图5,其净辐射最大可使叶温变化7℃,而对流换热和潜热交换可使叶温变化在1℃左右。所以,可认为叶气温差形成的主要原因是太阳辐射的能量输入。
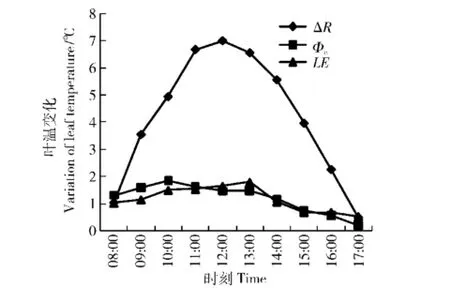
图5 各能量分支对叶温的贡献Fig.5 Temperature variation of each energy item
3 讨论
3.1 土壤干旱降低栓皮栎蒸腾速率
土壤水分供应不足势必影响植物蒸腾,中度至重度干旱下,栓皮栎的蒸腾速率与谢会成等[17]研究结论一致。研究表明,栓皮栎幼苗受土壤水分胁迫(田间持水量的40%和30%,相当于体积比12%和9%)时,蒸腾速率为0.841~1.485 mmol/(m2·s)。而轻度干旱时,高于文献[17]的蒸腾速率,说明轻度干旱对蒸腾速率影响较小,受土壤水分以外的其他因子影响,不同的试验可能有不同的蒸腾速率结果。在土壤水分为重度干旱(体积含水量5%~10%)时,蒸腾速率保持低水平,低于许多乔木树种蒸腾速率[18-20]。虽然耗水量减少,但不致死亡,表明栓皮栎是较好的耐旱和水土保持林种。
3.2 太阳辐射对栓皮栎蒸腾速率的主导作用
在3种土壤干旱胁迫条件下,栓皮栎叶蒸腾速率与太阳辐射的关系均呈直线正相关,表明太阳辐射是蒸腾的主导因子。桑玉强等[21]研究表明,蒸腾速率与太阳辐射正相关,且相关密切;与空气温度在不同季节的相关性不同,且小于太阳辐射,这与笔者结论相互印证。佟长福等[22]研究紫花苜蓿叶水势和蒸腾速率,也得出蒸腾速率与净辐射密切相关,在一定范围内呈直线关系(相关系数0.787 1,显著水平F>F0.01),说明太阳辐射也是紫花苜蓿蒸腾的主导因子。于金凤等[23]用TDP法测定苹果树蒸腾速率日变化特征,白天单峰型,夜间接近于0;蒸腾速率与太阳辐射的峰值出现时间相近,二者呈正相关,总辐射是影响果树蒸腾速率变化的直接因子;蒸腾速率与气温的峰值出现时间不同,略早于气温,这与笔者研究结果一致。罗永忠等[24]研究水分胁迫对紫花苜蓿蒸腾速率的影响时发现,蒸腾随胁迫增加而下降,这与笔者有相似之处。
3.3 气温不是蒸腾速率变化的主导因子
气温与蒸腾速率的关系在自然条件下,晴天时不同土壤湿度均表现为不密切,证明温度不是主导因子。人工气候箱试验结果表明,蒸腾速率与气温均呈现直线相关,蒸腾速率基本上是随温度升高而线性升高,但蒸腾速率明显低于晴天自然环境下,轻度干旱条件下最大值只有1.4 mmol/(m2·s),重度干旱下只有0.6 mmol/(m2·s)。也进一步说明与太阳辐射的作用相比,气温不是主导因子。这与桑玉强等[21]研究认为蒸腾速率与空气温度在不同季节的相关性不同,且小于太阳辐射的结论一致。佟长福等[22]通过蒸腾速率的日变化与气温的关系得出,蒸腾速率与温度的关系是先随温度升高而升高,到20.3℃后,随温度增加而降低,呈二次抛物线关系。笔者人工气候箱试验结果证明并非如此,只从日变化中看,随温度升高蒸腾是升高的,因为太阳辐射也在升高,午后太阳辐射下降,蒸腾速率主要受太阳辐射控制,即使温度在上升,蒸腾速率也要下降。所以得出的二次抛物线是不确切的,因为它没有排除太阳辐射的影响。前述桑玉强、于金凤等也得出气温对蒸腾的相关性小于太阳辐射的结论,印证了笔者的观点。
3.4 空气湿度对植物蒸腾的作用
空气湿度有绝对湿度(水汽压)、相对湿度和饱和差(VPD)等多个量来表示。由于空气温度的日变化,饱和水汽压随之变化,使空气相对湿度和饱和差在一天内有较大变化。而空气湿度对植物蒸腾的影响,主要用VPD来分析。VPD越大,越有利于植物蒸腾,从而降低叶温;但VPD与植物蒸腾和叶气温差的关系较难确定,这在CWSI研究的“非水分亏缺基线”确定研究中,表现很明显。如有人用14:00时冠气温差和VPD实测结果确定[25],还有人用冠气温差和VPD在10:00—14:00时有线性关系的时段回归确定[26]。未来应该在人工气候室内,控制其他因子为适宜条件下,设置不同VPD条件,测定植物蒸腾,确定其关系。
3.5 叶温对植物蒸腾的作用
叶温在一天中不同时间与气温有不同的差异,表现为越接近中午,叶气温差越大。王佩舒等[8]研究栓皮栎叶片的气候空间列线表明:太阳辐射越强,叶温越显著高于气温;但叶温对植物蒸腾的作用像气温的影响规律一样,如果没有太阳辐射(如夜间),较高的叶温也没有植物的蒸腾作用,所以,叶温不是植物蒸腾的主导因子。
3.6 叶温变化决定于叶表面净辐射
叶表面热量收支平衡分析表明,叶表面净辐射是叶温变化的主要控制因子,因为其在叶表面吸收的太阳辐射中,白天大部分时间占50%~70%的份额,对流换热量和潜热交换量仅占10%~20%的份额。各能量收支项对叶温变化贡献的量化计算表明,净辐射使叶温变化7℃左右,而对流换热和蒸腾潜热交换使叶温变化1℃左右。这些都表明太阳辐射是叶温升高的主导因子。许多研究将多种气象因子在蒸腾中的作用进行分析,认为均相关。如陈歆等[27]研究土壤水分对槟榔蒸腾速率的影响,认为槟榔蒸腾速率与气孔导度、胞间CO2浓度、空气温度、湿度及饱和差均达到显著性相关。事实上,各因子的作用是不同的,笔者研究认为,太阳辐射是主导因子,气温的影响小于太阳辐射。
根据栓皮栎叶气温差的变化特征,未来在建立CWSI时,确定“无蒸腾基线”和“非水分亏缺基线”时,应该以太阳辐射为主导因子,构建基于栓皮栎叶气温差的CWSI。
4 结论
在本次试验的土壤类型和气候类型条件下,通过分析不同土壤水分的自然和人工气候箱环境的蒸腾速率特征,结合叶片热量收支和叶片热物性的叶温形成分析,得出以下结论:
1)轻度干旱下,栓皮栎幼苗叶片的蒸腾速率为2 mmol/(m2·s),最大为2.5 mmol/(m2·s),轻度干旱胁迫不显著降低蒸腾速率,重度干旱下,蒸腾速率降低到1.5 mmol/(m2·s)以下,但不死亡,表现出较强耐旱性。
2)太阳辐射是影响栓皮栎蒸腾速率的主导因子,二者正相关。土壤水分和多云天气可影响这种相关性,二者的原因不同。前者是因为严重干旱条件下,水分供应不足时,太阳辐射增加也不能提高蒸腾速率;后者是因为多云短时间降低太阳辐射,但蒸腾速率不随之同步下降。
3)在自然环境中,气温不是栓皮栎蒸腾速率变化的主导因子,因为它与主导因子太阳辐射和其他因子对蒸腾产生复合影响。人工气候箱控制太阳辐射500 W/m2、空气湿度40%时,蒸腾速率与气温显著相关;但在轻度干旱下,温度从25℃增加到43℃,蒸腾速率从0.6增加到1.5 mmol/(m2·s),变化幅度较小,表现出温度对蒸腾的作用较小。
4)太阳辐射通过叶片净辐射输入叶片能量,而主导叶温形成。叶表面的净辐射对叶温形成的贡献达到50%~70%,能使叶温变化7℃左右;而对流换热和蒸腾潜热交换对叶温变化的贡献分别仅占10%~20%,各使叶温变化1℃左右:所以,太阳辐射是叶温形成的主导因子。
[1] 谢会成.栓皮栎光合生理生态的研究[D].南京:南京林业大学,2002:23. XIE Huicheng.A study on the photosynthetic physioecology of oriental oak[D].Nanjing:Nanjing Forestry University,2002:23.
[2] 刘震.我国水土保持情况普查及成果运用[J].中国水土保持科学,2013,11(2):1 LIU Zhen.Soil and water conservation survey in China and its application[J].Science of Soil and Water Conservation,2013,11(2):1.
[3] 马莉薇,张文辉,薛瑶芹,等.秦岭北坡不同生境栓皮栎实生苗生长及其影响因素[J].生态学报,2010,30 (23):6512. MA Liwei,ZHANG Wenhui,XUE Yiaoqin,et al. Growth characteristics and influencing factors of Quercus variabilisseedlingsonthenorthslopeofQinling Mountains[J].Acta Ecologica Sinica,2010,30(23): 6512.
[4] 张劲松,孟平,高峻,等.基于冠层叶气温差的苹果园土壤水分预报模型[J].农业工程学报,2007,23(6): 17. ZHANG Jinsong,MENG Ping,GAO Jun,et al.Model for predicting soil water status based on the canopy-air temperature differential in apple orchard[J].Transactions of the Chinese Society of Agricultural Engineering,2007,23(6):17.
[5] PAN Luan,ADAMCHUK V I,FERGUSON R B,et al. Analysis of water stress prediction quality as influenced by the number and placement of temporal soil-water monitoring sites[J].Journal of Water Resource and Protection, 2014,6:961.
[6] YANG Yongmin,SU Hongbo,ZHANG Renhua.A new evapo-transpiration model accounting for advection and its validation during SMEX02[J].Advances in Meteorology, Volume 2013,2013:1.
[7] JAHAN M S,NORDIN M N B,LAH M K B C,et al. Effects of water stress on rice production:bioavailability of potassium in soil[J].Journal of Stress Physiology& Biochemistry,2013,9(2):97.
[8] 王佩舒,王威红,陈景玲,等.栓皮栎幼苗叶气温差随太阳辐射和风速变化的列线研究[J].中国农业气象, 2016,37(2):213. WANG Peshu,WANG Weihong,CHEN Jingling,et al. Series lines on change of leaf temperature difference of Quercus variabilis with solar radiation and wind speed [J].Journal of Chinese Agricultural Meteorology,2016, 37(2):213.
[9] 严菊芳,张嵩午.不同温型小麦灌浆结实期农田热量平衡及其气象效应[J].西北农林科技大学学报, 2007,35(9):49. YAN Jufang,ZHANG Songwu.Study on the heat balance and the meteorological effect during milk-filling and burliness stage of the different type wheat[J].Journal of Northwest A&F University,2007,35(9):49.
[10] 刘云,宇振荣,孙丹峰,等.冬小麦冠气温差及其相关影响因素关系研究[J].灌溉排水学报,2004,23(1): 30. LIU Yun,YU Zhenrong,SUN Dan.Difference of canopy-air temperature and its affecting factors[J].Journal of Irrigation and Drainage,2004,23(1):30.
[11] 刘婧然,武金坤,王喆,等.棉花叶温与气象条件的关系研究[J].节水灌溉,2013(2):1. LIU Jingran,WU Jinkun,WANG Zhe,et al.Relation between cotton leaf temperature and meteorological conditions[J].Water Saving Irrigation,2013(2):1.
[12] 张寄阳,段爱旺,孙景生等.作物水分状况自动监测与诊断的研究进展[J].农业工程学报,2006,22(1): 174. ZHANG Jiyang,DUAN Aiwang,SUN Jingsheng,et al. Advances in automated monitoring and diagnosis of crop water status[J].Transactions of the Chinese Society of Agricultural Engineering,2006,22(1):174.
[13] 崔玉川,张文辉,王校峰.栓皮栎幼苗对土壤干旱胁迫的生理响应[J].西北植物学报,2013,33(2): 0364. CUI Yuchuan,ZHANG Wenhui,WANG Xiaofeng. Physiological responses of Quercus variabilis seedlings to soil drought stress[J].Acta Botanica Boreali-Occidentalia Sinica,2013,33(2):0364.
[14] GATES D M.Biophysical ecology[M].New York: Springer-verleg,1980:15.
[15] 刘静,王存诚.生物传热学[M].北京:科学出版社, 1997:36. LIU Jing,WANG Chuncheng.Biological heat transfer [M].Beijing:Science Press,1997:36.
[16] 陈景玲,王谦,朱秀红,等.番茄叶片和果实热容量的热平衡测定方法[J].农业工程学报,2012,28(2): 279. CHEN Jingling,WANG Qian,ZHU Xiuhong,et al. Specific heat of tomato leaf and fruit with heat balance method[J].Transactions of the Chinese Society of Agricultural Engineering,2012,28(2):279.
[17] 谢会成,朱西存.水分胁迫对栓皮栎幼苗生理特性及生长的影响[J].山东林业科技,2004.4:6. XIE Huicheng,ZHU Xicun.The effect of water stress on the photosynthetic characteristics and growth yield of oriental oak seedlings[J].Shandong Forestry Science and Technology,2004.4:6.
[18] 杨建伟,韩蕊莲,刘淑明,等.不同土壤水分下杨树的蒸腾变化及抗旱适应性研究[J].西北林学院学报, 2004,9(3):7. YANG Jianwei,HAN Leilian,LIU Shuming,et at. Transpiration and drought resistance of poplar under different soil drought[J].Journal of Northwest Forestry U-niversity,2004,9(3):7.
[19] 廖行,王百田,武晶,等.不同水分条件下核桃蒸腾速率与光合速率的研究[J].水土保持研究,2007,14 (4):30. LIAO Hang,WANG Baitian,WU Jing,et al.Study on transpiration rate and photosynthesis rate of Juglan sregia under different soil water contents[J].Research of Soil and Water Conservation,2007,14(4):30.
[20] 王旭军,吴际友,廖德志,等.响叶杨光合蒸腾和水分利用效率对光强及CO2浓度升高的响应[J].南京林业大学学报(自然科学版),2009,33(2):55. WANG Xujun,WU Jiyou,LIAO Dezhi,et al.Response of photosynthesis rate,transpiration rate and water using efficiency of Populus adenopoda to light intensity and elevated CO2concentration[J].Journal of Nanjing Forestry University(Natural Science Edition), 2009,33(2):55.
[21] 桑玉强,郭芳,张劲松,等.毛乌素沙地新疆杨蒸腾变化规律及其影响因素[J].林业科学,2009,45(9): 66. SANG Yuqiang,GUO Fang,ZHANG Jinsong,et al. Variation in transpiration of Populus bolleana and the influence factors in Maowusu sandy area[J].林业科学, 2009,45(9):66.
[22] 佟长福,郭克贞,史海滨,等.环境因素对紫花苜蓿叶水势与蒸腾速率影响的初步研究[J].农业工程学报,2005,21(12):152. TONG Changfu,GUO Kezhen,SHI Haibin,et al.Preliminary study on the effect of environmental factor on leaf potential and transpiration rate of alfalfa[J].Transactions of the Chinese Society of Agricultural Engineering,2005,21(12):152.
[23] 于金凤,刘文兆,甘卓亭,等.黄土塬区苹果树蒸腾速率变化特征及其影响因子[J].干旱地区农业研究, 2010,28(4):59. YU Jinfeng,LIU Wenzhao,GAN Zhuoting,et al.The characteristics of apple tree transpiration rate and its influencing factors on the loess tableland region[J].Agricultural Research in the Arid Areas,2010,28(4):59.
[24] 罗永忠,成自勇.水分胁迫对紫花苜蓿叶水势、蒸腾速率和气孔导度的影响[J].草地学报,2011,19(2): 215. LUO Yongzhong,CHENG Ziyong.Impact of water stress on leaf water potential,transpiration rate and stomatal conductance of alfalfa[J].Acta Agrestia Sinica,2011,19(2):215.
[25] 张喜英,裴冬,陈素英.用冠气温差指导冬小麦灌溉的指标研究[J].中国生态农业学报,2002,10(2): 102. ZHANG Xiying,PEI Dong,CHEN Suying.Use of crop water stress index as indicators for scheduling irrigation in winter wheat[J].Chinese Journal of Eco-Agriculture,2002,10(2):102.
[26] SALEH T,JOSE L C,NEIL C H.Infrared thermometry to estimate crop water stress index and water use of irrigated maize in northeastern Colorado[J].Remote Sens. 2012,4,3619.
[27] 陈歆,刘贝贝,彭黎旭.土壤水分对槟榔幼苗净光合速率和蒸腾速率的影响[J].热带作物学报,2015,36 (11):2034. CHEN Xin,LIU Beibei,PENG Lixu.Effects of different soil moisture on photosynthetic rate and transpiration rate of Areca catechu L.seedling[J].Chinese Journal of Tropical Crops,2015,36(11):2034.
Impacts of radiation,temperature and soil moisture on hidden heat of transpiration and leaf temperature of Quercus variabilis seedlings
CHEN Jingling1,WANG Peishu1,LIU Linqi1,WANG Qian1,WANG Xu1,YANG Xitian1,ZHANG Jinsong2
(1.College of Forestry,Henan Agricultural University,450002,Zhengzhou,China; 2.Research Institute of Forestry,Chinese Academy of Forestry,100091,Beijing,China)
[Background]Drought threats plants'survival and in the same time it causes variation of their leaf temperature(TL),which provides people an efficient path to diagnose soil moisture.Many scholars have been using crop leaf-air temperature deficit(LATD)to calculate crop water stress index (CWSI).However,there are few studies on leaf-air temperature ofQuercus variabilis,and no works on diagnose of soil moisture.The transpiration(Tr)ofQ.variabilisis less than that of crops,and its LATD is larger than agricultural crop,thus the water stress index(WSI)ofQ.variabilisshould be different to CWSI.The key that uses LATD to evaluate water stress is to analyze the energy factors of leaf temperature.[Methods]Watering the pots then soil moisture decreased through natural evaporation,we controlled soil moisture in 3 levels:light drought,12.5%-14.5%(W1);moderate drought,9.5%-11.5%(W2),and severe drought,5.5%-7.5%(W3).Selecting clear and cloudy days,from 8:00 to 18:00,we measuredTrandTLas well as air temperature(t)and solar radiation(R).We also measuredTrin artificial chamber controlling temperature at 25-43℃,at every 2℃interval of treating temperature.According to the ratio of net radiation ΔR,convectionΦc,latent heat of transpirationLE, and leaf area densityρ,heat capacityCp:ΔR/ρCp,Φc/ρCp,LE/ρCp,we calculated the contribution of each energy item toTL.[Results]1)TheTrofW1was close to of normal soil moisture,the maximumTrwas 2.5 mmol/(m2·s).TheTrofW2andW3reduced to be<1.5 mmol/(m2·s).2)In clear day,Trpositively related to solar radiation,which indicated that solar radiation was main factor ofTr.The slopes of the regression line betweenTrand solar radiation were different with different soil moistures,the dryer the soil was,and the greater the slope was.3)In cloudy day,the relation coefficient betweenTrand solar radiation decreased clearly,which meantTrdid not decline with short time interval of radiation reducing,once transpiration started in the morning.4)On any case of soil moisture,air temperature was not main factor ofTr.Though in artificial chamber the linear relation ofTrand temperature was clear,Trwas lower than natural clear day.[Conclusions]1)Light drought did not impactTrofQ.variabilis.2) When wind speed was zero,solar radiation was main factor of leaf temperature variation.Net radiation accounted for 50%-70%of energy budget,and provided approximately 7℃of leaf temperature variation.3)The latent heat ofTrand convection heat exchange had an equal effect on energy budget, which accounted for 10%-20%respectively.Each of two factors caused approximately 1-2℃of leaf temperature variation respectively.
Quercus variabilis;radiation;temperature;soil moisture;transpiration;leaf temperature
S167
:A
:2096-2673(2017)01-0062-09
10.16843/j.sswc.2017.01.008
2016- 08- 04
2016- 08- 20
项目名称:国家自然科学基金“基于气候空间上限列线的栓皮栎幼苗高温叠加干旱胁迫机制研究”(31370621),“番茄叶片低温胁迫气象指标的气候空间列线研究”(31071321)
陈景玲(1964—),女,副教授,硕士生导师。主要研究方向:农林气象。E-mail:chenjingling5@163.com
†通信作者简介:王谦(1963—),男,教授,硕士生导师。主要研究方向:农林气象和林业生态。E-mail:wangqianhau@163.com
猜你喜欢
气候与环境研究(2022年4期)2022-08-15
农业工程学报(2022年5期)2022-06-22
干旱区地理(汉文版)(2022年3期)2022-05-30
水土保持学报(2022年3期)2022-05-26
现代农村科技(2021年11期)2021-11-05
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
中国农业科技导报(2020年12期)2020-03-15
湖南大学学报·自然科学版(2015年10期)2015-11-30
建筑科学与工程学报(2015年3期)2015-07-25
