
河南北部稻蛀茎夜蛾的发生规律研究初报
2017-03-29 03:11黄建荣李国平田彩红封洪强
植物保护 2017年2期
黄建荣, 黄 博, 李国平, 田彩红, 封洪强
(河南省农作物病虫害防治重点实验室,农业部华北南部作物有害生物综合治理重点实验室,河南省农业科学院植物保护研究所,郑州 450002)
河南北部稻蛀茎夜蛾的发生规律研究初报
黄建荣, 黄 博, 李国平, 田彩红, 封洪强*
(河南省农作物病虫害防治重点实验室,农业部华北南部作物有害生物综合治理重点实验室,
河南省农业科学院植物保护研究所,郑州 450002)
随着全球气候变暖,昆虫分布及越冬地域也在发生变化。稻蛀茎夜蛾Sesamiainferens(Walker)的越冬北限在20世纪80年代被认为不过北纬34°线,近年发现该虫在更北的黄淮海夏玉米-小麦轮作种植区新乡市(北纬35°)发生为害,为该种植模式下发生的新害虫。为了准确监测,科学、有效地制定该害虫的防治对策,本研究对河南北部地区稻蛀茎夜蛾的发生为害规律及生活史进行了初步调查。结果表明:稻蛀茎夜蛾可以在小麦-玉米轮作的条件下顺利完成生活史,其中在河南新乡该虫为害穗期玉米并顺利越冬的事实是首次报道。应用灯光监测并结合田间幼虫调查发现,当地稻蛀茎夜蛾一年发生3代。越冬代幼虫化蛹后在4月中下旬开始羽化,羽化的成虫在小麦或早春玉米上产卵,第1代幼虫主要为害春玉米和小麦,6月中旬第1代成虫开始羽化飞往夏玉米田内产卵,幼虫主要为害夏玉米幼苗。第2代幼虫化蛹后于8月初开始羽化形成第2代成虫,产卵孵化后的幼虫主要钻蛀夏玉米穗和茎秆,9月下旬幼虫陆续沿茎秆钻入玉米残桩基部越冬。综合调查结果,绘制了河南北部地区稻蛀茎夜蛾的年生活史表。
稻蛀茎夜蛾; 玉米; 种群动态; 生活史
气候变暖不仅直接影响着昆虫的生长发育和繁殖[1],还影响各类昆虫的地域分布,使得昆虫向高纬度、高海拔等温度适宜地区迁移扩散,害虫的适合生长区域范围发生变化,为害时期和地域也相应发生了变化[2-3]。稻蛀茎夜蛾Sesamiainferens(Walker),又称大螟,隶属于鳞翅目、夜蛾科、蛀茎夜蛾属Sesamia[4]。国外分布于产稻国家,在中国据记载分布北限为陕西周至、河南信阳、安徽合肥、江苏淮安,大致在北纬34°线以南[5-6]。稻蛀茎夜蛾是杂食性昆虫,主要寄主有水稻、麦类、玉米、高粱、谷子、茭白、野茭白、香蒲、芦苇、油菜、稗、向日葵以及禾本科和莎草科杂草。在水稻、茭白、野茭白、玉米上能完成完整的世代[7-9]。随着耕作制度、作物布局及药剂使用上的变化,我国南方许多地方稻蛀茎夜蛾成为主要的水稻害虫之一,为害程度不断加重。稻蛀茎夜蛾也为害玉米,是印度半岛玉米生产中主要的害虫[10],在我国,稻蛀茎夜蛾在南方稻区也为害玉米[11],主要为害春玉米。最新研究显示,稻蛀茎夜蛾在我国更北的黄淮海夏玉米区发生并为害夏玉米苗[12],而该虫在夏玉米种植区是否能顺利完成整个世代,其发生为害规律如何还没有相关报道。本研究通过灯光诱集成虫,田间调查玉米上幼虫的种群动态,拟明确稻蛀茎夜蛾在玉米上的发生为害规律和发生世代,为该虫的监测和防治提供依据。
1 材料与方法
1.1 成虫种群动态的监测
监测点位于河南省新乡市河南省农科院第二基地(35.02°N, 113.69°E)内。本研究中采用探照灯诱虫器诱集稻蛀茎夜蛾成虫,3月中旬开灯,11月初关灯。灯在日落后开启,每日约在23:00~0:30之间取回前半夜诱集的活虫,次日日出后关灯,取回诱虫器内所诱集的后半夜活虫,活虫取回马上置于冷冻冰箱内冻死,记录稻蛀茎夜蛾成虫的种群数量。
1.2 幼虫的田间调查
玉米品种为‘郑单958’和‘浚单20’。2014年6月2日和2015年6月10日播种,行距约0.85 m,株距约0.3 m。调查试验共设3个小区,连续排列,小区面积约200 m2,翻耕和播种均采用机械化进行,试验地在玉米生育期内的农事操作,如除草、灌溉、化肥等耕作管理和常规田一致,苗期至穗期玉米的长势较健壮,与常规夏玉米长势一致。每隔7天调查1次,按5点取样法进行,每个调查点单行连续调查5株后,采取前进式再调查邻近行内的另外5株,每点共调查10株玉米,每小区内共调查50株。玉米进入小喇叭口期之后采用定株调查,对每株玉米做好标记。调查时若标记好的植株被人为损坏,采用邻近未标记的植株替换。统计每个小区稻蛀茎夜蛾幼虫的总数量。越冬调查在2016年2月18日进行,调查新乡市原阳县师寨镇任庄村(35.12°N, 113.87°E)、亢村镇府庄村(35.06°N, 113.72°E)和亢村镇(35.11°N, 113.69°E)地头残存的玉米秆、桩及附近土层内是否存在活体稻蛀茎夜蛾,记录越冬部位及虫龄。
1.3 种群动态分析
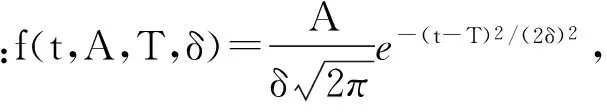
2 结果与分析
2.1 稻蛀茎夜蛾成虫种群动态
连续两年对稻蛀茎夜蛾成虫的监测发现,2015年主要有3个成虫高峰,越冬代成虫始见期为4月26日,羽化高峰日为5月14日,末期为6月7日;第1代成虫始盛期为6月17日,高峰日为7月4日,末期为7月7日;第2代成虫始盛期为8月4日,第2代高峰日为9月8日,末期为10月13日,第1代成虫量较越冬代和第2代虫量低(图1)。2014年主要有3个成虫高峰,越冬代成虫始见期为4月14日,羽化高峰日为4月23日,末期为5月27日;第1代成虫始盛期为6月13日,高峰日为7月1日,末期为7月24日;第2代成虫始盛期为8月3日,高峰日为8月25日,末期为10月22日(图1)。两年成虫诱集总量比较发现,2015年第1~2代成虫量明显较2014年虫量少(图1)。对比前后半夜诱集的成虫量表明,2015年第2代成虫主要是后半夜上灯,诱集的虫量较前半夜多,而2014年成虫前半夜明显较后半夜诱集的成虫量多(图2)。

图1 稻蛀茎夜蛾成虫的种群动态Fig.1 Population dynamics of Sesamia inferens moths

图2 前半夜与后半夜稻蛀茎夜蛾的上灯虫量Fig.2 Number of Sesamia inferens moths caught by light trap before and after mid-night
2.2 稻蛀茎夜蛾幼虫种群动态
调查发现6月初春玉米上有稻蛀茎夜蛾高龄幼虫,依据越冬代成虫羽化高峰日为5月14日(2015)和4月23日(2014),调查到的应为第1代幼虫。在6月中下旬发现有该虫幼虫(龄期2~5龄)钻蛀夏玉米苗心叶,导致幼苗死亡,造成单株产量100%损失。7月中旬至8月中下旬也调查到幼虫钻蛀心叶,排出大量虫粪,根据第1代成虫高峰日在7月4日(2015年)和7月1日(2014年),应为第2代幼虫。9月份之后,一直能见到幼虫为害玉米秆或玉米穗,虫口密度较喇叭口期更大,在9月下旬仍发现有2~3龄幼虫在玉米秆和穗上为害,该代幼虫直接在玉米桩内越冬,为越冬代的幼虫(图3)。

图3 稻蛀茎夜蛾幼虫在两个玉米品种上的种群动态Fig.3 Larval population dynamics of Sesamia inferens in two varieties of corn
2.3 稻蛀茎夜蛾为害穗期玉米
前期研究报道了河南北部夏玉米苗期受稻蛀茎夜蛾的为害[12],经过两年的田间调查表明,稻蛀茎夜蛾除在苗期为害玉米苗外,还在穗期为害玉米穗。幼虫咬断花丝,从雌穗顶端钻入苞叶内咬食嫩玉米籽粒直至玉米成熟(图4a~d),随着玉米的成熟,幼虫可钻入穗轴内为害,被咬食的籽粒发生霉变,不但降低玉米产量而且降低玉米的品质(图4e~h)。

图4 被稻蛀茎夜蛾幼虫为害的夏玉米穗Fig.4 Corn ears damaged by larvae of Sesamia inferens in summer corn field
2.4 稻蛀茎夜蛾在夏玉米上的越冬
春后调查发现,稻蛀茎夜蛾幼虫在玉米残桩内越冬,越冬部位为土面下方2cm左右处的茎秆基部或玉米茎秆内(图5)。同时也发现,在地下2cm左右的玉米桩内越冬的幼虫存活率高,越冬虫态以3~4龄幼虫为主。而在玉米残桩地上部分越冬的幼虫全部死亡,死亡虫龄以4~5龄为主(图6)。
2.5 稻蛀茎夜蛾在河南北部的生活史
根据稻蛀茎夜蛾幼虫的田间调查结果,结合成虫种群动态,制定了稻蛀茎夜蛾年生活史表(表1)。稻蛀茎夜蛾在河南北部一年发生3代,越冬幼虫化蛹后在4月中下旬开始羽化,羽化的成虫可以在小麦或早春玉米上产卵,第1代幼虫主要为害春玉米和小麦。6月中旬第1代成虫开始羽化产卵,第2代幼虫主要为害夏玉米幼苗。2代成虫于8月初开始羽化产卵,幼虫主要钻蛀夏玉米穗和茎秆为害,幼虫直接在9月下旬开始滞育在玉米残桩及土表层越冬。

图5 夏玉米上稻蛀茎夜蛾的越冬部位Fig.5 Overwintering site of Sesamia inferens insummer corn field
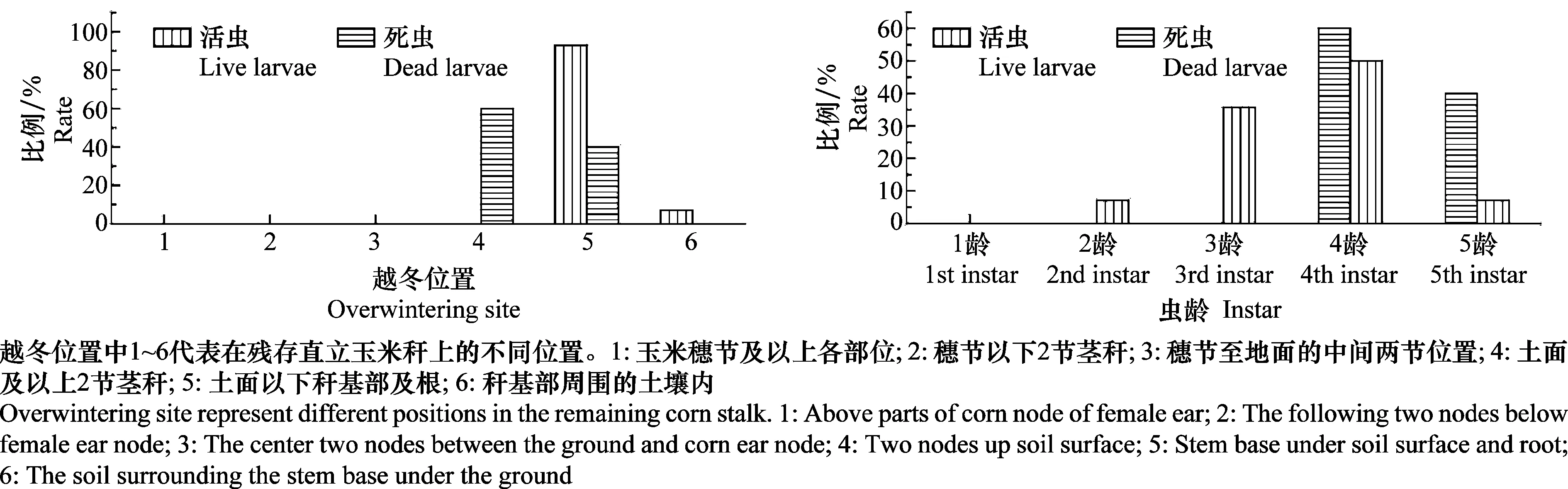
图6 夏玉米田内稻蛀茎夜蛾的越冬部位及龄期Fig.6 Overwintering site and instar of Sesamia inferens in summer corn field

世代Generation虫态 Stage4月April上中下5月May上中下6月June上中下7月July上中下8月August上中下9月September上中下10月October上中下11-3月November-March上中下越冬代Overwinteringgeneration(-)(-)(-)⊕⊕⊕++++第1代Firstgeneration···------⊕⊕⊕⊕+++++第2代Secondgeneration···-------⊕⊕⊕⊕⊕⊕++++++++第3代(越冬代)Thirdgeneration(overwinteringgeneration)······------(-)(-)(-)
1) 作者未调查卵和蛹,表中的·和⊕是依据幼虫和成虫出现时间推测的。·卵;- 幼虫;(-) 越冬幼虫;⊕蛹;+成虫。 The authors did not investigate the egg and pupa. ·and⊕ in the table were speculated by the appearing time of larvae and adult. ·Egg; - Larva; (-) Overwintering larva; ⊕ Pupa;+Adult.
3 讨论
气候变暖将导致昆虫向高海拔高纬度地区扩展[14-18],为害区域的北扩,给高纬度地区带来严重的经济损失[19-20]。1965年,章士美[21]提出稻蛀茎夜蛾在我国的发生不过淮河。1985年顾海南[6]在研究稻蛀茎夜蛾的越冬参数后认为该虫的越冬北界在北纬34°以南。经过调查,我们首次报道了稻蛀茎夜蛾在黄淮海夏玉米区的新乡地区为害夏玉米苗,又经过两年的持续监测,发现稻蛀茎夜蛾不仅一直为害夏玉米苗,还对玉米穗造成危害,直接导致产量的损失,稻蛀茎夜蛾已成为河南北部夏玉米上重要的新害虫。调查还发现在河南新乡稻蛀茎夜蛾以幼虫在夏玉米残桩内越冬,这与其在南方水稻桩内的越冬方式一致,但该虫在地下2 cm左右的玉米桩基部以3~4龄幼虫越冬。本研究调查的越冬北界较北纬34°的结果北扩了近1°多,这可能与气候变暖有关,而该虫是否还在更北的区域发生和为害,还需要更进一步的调查研究。
1965年,宁波专区农业科学研究所植保组[22]的试验结果表明,稻蛀茎夜蛾在宁波地区每年发生3~4代,第3代螟蛾产的卵,在8月20日以前孵化的幼虫都要变为第4代,8月20日以后孵化的幼虫多以老熟幼虫越冬。李洪山等[23]2002年的研究结果表明,在更北的江苏盐城,稻蛀茎夜蛾每年发生3~4代,第4代为不完全世代。本研究结合田间夏玉米上幼虫的调查,可以确定该虫在河南北部每年可以发生完整的3代,第3代高龄幼虫在新乡可以顺利越冬,但10月下旬田间幼虫调查时发现有低龄幼虫(2龄),春后越冬调查也发现有2龄幼虫,由于本文拟合的第2代成虫期从8月至10月,是否存在第2、第3代成虫世代重叠现象,田间调查到的低龄幼虫是否是第4代幼虫,第4代幼虫是否可以春后化蛹并顺利羽化,还有待更为细致的研究,但目前新乡稻蛀茎夜蛾发生世代数与50年前宁波(纬度相差5°)的调查研究结果相同,说明该害虫在南方地区有可能随着气候的变暖发生世代数增加。本文为常规的田间调查结果,田间查到的卵和蛹非常复杂,无法根据作者经验确定是否是稻蛀茎夜蛾的卵或者蛹,只能根据幼虫和成虫发生时期推测出卵和蛹的发生时期,并组建河南北部地区稻蛀茎夜蛾的生活史。稻蛀茎夜蛾在玉米植株上的产卵是否存在选择性还需要更多的对比研究。前人一直认为稻蛀茎夜蛾的趋光性不强,采用灯光监测不能反映成虫的消长状况[24-26],探照灯诱虫器采用的1 000 W金属卤化物灯泡有别于黑光灯,光柱朝向空中,对高空至少500 m处的昆虫种群具有诱集效应[27],本研究两年的连续监测结果发现稻蛀茎夜蛾成虫发生有明显的消长节律,能清楚地反映该虫成虫的发生数量和发生高峰。同时两年的监测结果显示各代成虫量存在较大的差异,这是否与两年之间气候变动或者其他原因有关,还需更多年份或者更多监测点的调查数据才能说明。本研究中稻蛀茎夜蛾幼虫在玉米上发生3代,第1代为害春玉米和小麦,第2代为害夏玉米苗,第3代为害穗期玉米,给玉米产量造成损失,两年的调查均发现第3代幼虫为害穗期玉米后在玉米残桩内顺利完成越冬,说明稻蛀茎夜蛾在该地区能完成生活史,存在本地虫源。害虫的迁飞导致农作物被害区域更大,在我国,迁飞性害虫每年春季的北迁导致北方农作物受害严重。据报道,稻蛀茎夜蛾的有效飞行日龄为6 d,测试其成虫在晚上的飞行能力表明,成虫累计飞行距离<5 km的个体占75.5%[28],如果持续吊飞该虫,雌、雄虫可分别达30与50 km以上[29],但在水稻害虫测报和防治中,把稻蛀茎夜蛾归为非迁飞性害虫,不考虑该虫的远距离迁飞。一般认为迁飞性害虫的本地虫源大部分在前半夜起飞,后半夜收集到的成虫更可能为外地虫源[30],本研究对第1~2代成虫的监测发现,2014年前半夜成虫量多于后半夜,监测到第1~2代成虫种群数量较大,2015年后半夜成虫量大于前半夜,而第1代成虫量少,第2代成虫量较大,是否存在第2代成虫迁入的可能还需要对该虫的起飞节律和虫源性质进行详细的研究。
[1] Bale J S, Masters G J, Hodkinson I D, et al. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores [J]. Global Change Biology, 2002, 8(1): 1-16.[2] 陈瑜, 马春森. 气候变暖对昆虫影响研究进展[J]. 生态学报, 2010, 30(8): 2159-2172.
[3] 张花龙, 杨念婉, 李有志, 等. 气候变暖对农业害虫及其天敌的影响[J]. 植物保护, 2015, 41(2): 5-15.
[4] 朱弘复, 方承莱, 王林瑶. 中国经济昆虫志第七册·夜蛾科(三)[M]. 北京: 科学出版社, 1963: 120.
[5] 丁锦华, 苏建亚. 农业昆虫学[M]. 北京: 中国农业出版社, 2002: 154.
[6] 顾海南. 大螟越冬特性的初步研究[J]. 生态学报, 1985, 5(1): 64-70.
[7] 金翠霞, 吴亚. 大螟与寄主植物关系的研究[J]. 植物保护学报, 1986, 13(4): 259-265.
[8] 欧克芳, 董立坤, 熊育剑, 等. 香蒲新害虫稻蛀茎夜蛾生物学特性研究[J]. 湖北农业科学, 2013, 52(2): 319-320.
[9] 徐红莲, 严兆龙, 仇广灿, 等. 大螟的转移为害规律及其测报技术探讨[J]. 植保技术与推广, 2001,21(5): 5-7.
[10]Santosh H B, Sekhar J C, Rakshit S, et al. Detection of epistatic interaction for susceptibility towards pink stem borer (SesamiainferensWalker) in maize (ZeamaysL.)[J]. Indian Journal of Genetics and Plant Breeding, 2012, 72(3): 284-289.
[11]谢大赉, 蔡如希, 赵福臻, 等. 大螟为害玉米的生物学特性与药剂防治试验[J]. 四川农学院学报, 1985, 3(5): 33-40.
[12]黄建荣, 封洪强. 河南北部稻蛀茎夜蛾为害夏玉米苗初报[J]. 植物保护, 2015, 41(2): 231-233.
[13]Wu Kongming, Lu Yanhui, Feng Hongqiang, et al. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton [J]. Science, 2008, 321(5896): 1676-1678.
[14]Parmesan C N, Ryrholm C, Steganescu C, et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming [J]. Nature, 1999, 399(6736): 579-583.
[15]Hickling R, Roy D B, Hill J K, et al. A northward shift of range margins in British Odonata [J]. Global Change Biology, 2005, 11(3): 502-506.
[16]Böhme W, Geissler P, Wagner P. A remarkable record ofPhaneropterafalcata(Poda, 1761) (Saltatoria: Phaneropteridae) from north-eastern Poland [J]. Bonn Zoological Bulletin, 2011, 60(1): 109-111.
[17]Musolin D L. Insects in a warmer world: ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change [J]. Global Change Biology, 2007, 13(8): 1565-1585.
[18]Mantyka-pringle C S, Martin T G, Rhodes J R. Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis [J]. Global Change Biology, 2012, 18(4): 1239-1252.
[19]Parmesan C. Ecological and evolutionary responses to recent climate change [J]. Annual Review of Ecology, Evolution, and Systematics, 2006, 37(1): 637-669.
[20]Sánchez-Guillén R A, Córdoba-Aguilar A, Hansson B, et al. Evolutionary consequences of climate-induced range shifts in insects [J]. Biological Reviews, 2016,91(4):1050-1064.
[21]章士美. 从某些农业昆虫的分布来讨论古北、东洋两地区在我国秦岭以东的分界线问题[J]. 昆虫学报, 1965, 14(4): 411-419.
[22]宁波专区农业科学研究所植保组. 宁波地区水稻大螟生活史观察和防治试验初报[J]. 浙江农业科学, 1965(5): 243-248.
[23]李洪山, 李慈厚, 李红阳, 等. 苏北稻区水稻大螟种群消长特点及在寄主间的转换规律[J]. 植保技术与推广, 2002, 22(10): 13-16.
[24]陈龙稳, 欧阳享泱, 吕锡麟. 大螟性诱剂应用于虫情测报[J]. 昆虫知识, 1987, 24(1): 8-10.
[25]徐丽娜, 李昌春, 胡本进, 等. 中国大螟研究历史、现状与展望[J]. 中国农学通报, 2011,27(24): 244-248.
[26]陈晓娟, 卢代华. 多食性害虫大螟发生与防治研究进展[J]. 中国农学通报, 2015, 31(25): 171-175.
[27]Feng H Q, Wu K M, Cheng D F, et al.Radar observations of the autumn migration of the beet armywormSpodopteraexigua( Lepidoptera: Noctuidae)and other moths in northern China [J].Bulletin of Entomological Research, 2003, 93(2): 115-124.[28]韩兰芝, 彭于发, 吴孔明. 大螟幼虫田间扩散及成虫飞行能力研究[J]. 植物保护, 2012, 38(4): 9-13.
[29]孙建中, 张建新, 沈雪生. 三化螟、二化螟及大螟成虫的飞翔能力[J]. 昆虫学报, 1993, 36(3): 315-322.
[30]齐会会, 张云慧, 王健, 等. 稻纵卷叶螟在探照灯下的扑灯节律[J]. 中国农业科学, 2014, 47(22): 4436-4444.
(责任编辑:杨明丽)
Preliminary study on the occurrence of pink stem borer,Sesamiainferens(Walker) (Lepidoptera: Noctuidae) in northern Henan Province
Huang Jianrong, Huang Bo, Li Guoping, Tian Caihong, Feng Hongqiang
(HenanKeyLaboratoryofCropPestControl,KeyLaboratoryofIntegratedPestManagementonCropsinSouthernPartofNorthernChina,MinistryofAgriculture;InstituteofPlantProtection,HenanAcademyofAgriculturalSciences,Zhengzhou450002,China)
Global warming and climate change are likely to trigger changes in geographical distribution and overwintering region of insect pests. The northern boundary of overwintering for pink stem borer,Sesamiainferens(Walker) (Lepidoptera: Noctuidae) was at 34° north latitude in 1980s. Recently, occurrence and damage ofS.inferenson summer corn seedlings has been first observed in wheat-maize rotation region of Xinxiang (35° north latitude), northern Henan Province. In order to develop a sound forecasting and management program in summer corn region of China, population dynamics and life history ofS.inferenswere investigated in Xinxiang from 2014 to 2015. The results indicated thatS.inferenscould finish its life history and damage on ears of summer corn in Xinxiang, which was the first report about its northern boundary in China. Light traps and field surveys showed there were three generations per year forS.inferensin this area. The overwintering larvae pupated and emerged in mid to late April, and then female adults laid eggs on winter wheat and spring corn. Larvae of the first generation damaged winter wheat and spring corn in late May and early June and mature larvae of this generation pupated and emerged after mid-June. The moths of the first generation transferred into summer corn field and oviposited in corn seedling for continuous life cycle. Larvae of the second generation damaged seedling after mid-July.S.inferenslarvae of the third generation (overwintering generation) damaged the stem and ear in summer corn field and drilled into the root of corn stalk for overwintering in late September. An annual life history table for theS.inferensin northern Henan Province was drawn based on these observed data.
Sesamiainferens; corn; population dynamics; life history
2016-03-28
2016-05-09
国家自然科学基金 (31401731);河南省超级产粮大省奖励资金扶持粮油良种培育项目(201513116);河南省农业科学院优秀青年基金(2016YQ15)
S 435.13
A
10.3969/j.issn.0529-1542.2017.02.027
* 通信作者 E-mail:Feng_hq@163.com
猜你喜欢
当代水产(2022年7期)2022-09-20
作物学报(2022年8期)2022-05-29
今日农业(2021年21期)2021-11-26
云南农业(2021年11期)2021-11-12
当代水产(2021年4期)2021-07-20
今日农业(2020年13期)2020-12-15
今日农业(2020年23期)2020-12-15
农民致富之友(2020年17期)2020-06-19
今日农业(2019年11期)2019-08-13
今日农业(2019年13期)2019-08-12
