
大宁河浮游植物季节演替与环境的响应关系
2017-03-21 03:12纪道斌刘德富宋林旭吕林鹏
中国农村水利水电 2017年4期
王 雄,纪道斌,刘德富,宋林旭,吕林鹏,李 欣
(1.三峡大学水利与环境学院,湖北 宜昌 443002;2.湖北工业大学 河湖生态修复与藻类利用湖北省重点实验室,武汉 430068)
自2003年三峡水库蓄水后,由于水动力及水环境条件的变化,部分支流多次出现了不同程度水华现象,水华频发[1]。库区支流水华优势种随季节而发生转变,从春季至冬季水华优势种依次为硅甲藻、蓝绿藻、绿藻、再到硅藻[2],具有明显的季节性演替特征。大宁河是三峡水库典型的支流之一,位于三峡库区腹心地带,自2003年6月首次在双龙暴发蓝绿藻水华后[3],2004年至今在河流各支段水华频发[4],已成为三峡水库大宁河库湾重要的水环境问题。
随着水环境问题的日益严重,针对三峡水库水华的研究愈发重要,而库区支流全年藻种演替规律的探索对水华的深入研究具有重要意义。因此,该研究基于2015年大宁河不同季节的水温、光照、营养盐、藻种等监测数据,探讨该河流浮游植物季节演替规律、空间分布的特点及其与环境因子的关系,为揭示三峡水库典型支流库湾水华机理及防控提供技术和理论支撑。
1 材料和方法
1.1 研究区域概况
大宁河回水段位于重庆市巫山县境内(E108.125°~108.135°,N30.195°~31.12°),距离三峡大坝123 km,河道蜿蜒,部分水域峡谷陡峭高深,而部分水域地势较为平坦,河面相对宽敞,如双龙、大昌河段。三峡水库蓄水后河水流速大大减缓,河面增宽,其回水段水域成为水库发生富营养化的敏感区域,多次暴发藻类水华[5-9]。同时,大宁河也是著名风景区小三峡所在河流,回水区内有大量频繁的旅游行船,强烈的行船扰动会对表层水体尤其是对狭窄河段水体造成剧烈掺混作用。
1.2 样点布置
根据河道形态以及大宁河历年的水华暴发情况,从大宁河下游出河口(长江)至上游回水区末端(大昌镇),分别设置了DN01~DN06共6个断面(见图1)。各样点地理环境比较如表1所示。
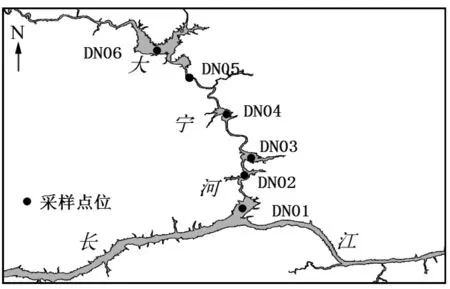
图1 大宁河采样点分布图Fig.1 Location of sampling sites in the Daning River
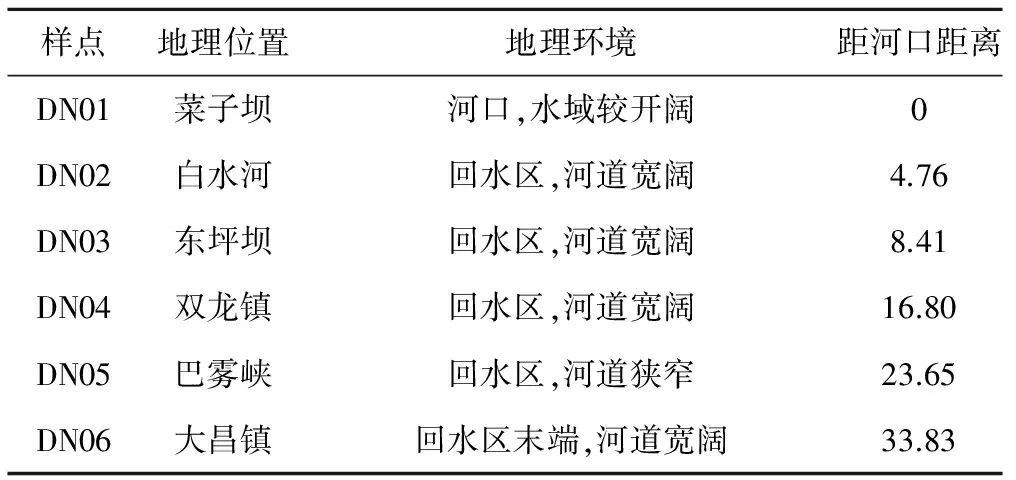
Tab.1 The comparison of geographical environment in different sampling sites
1.3 监测时段
2015年3月30日、8月1日、11月3日和2016年1月11日4个时段。
1.4 监测指标与方法
监测指标及相应方法如表2所示。
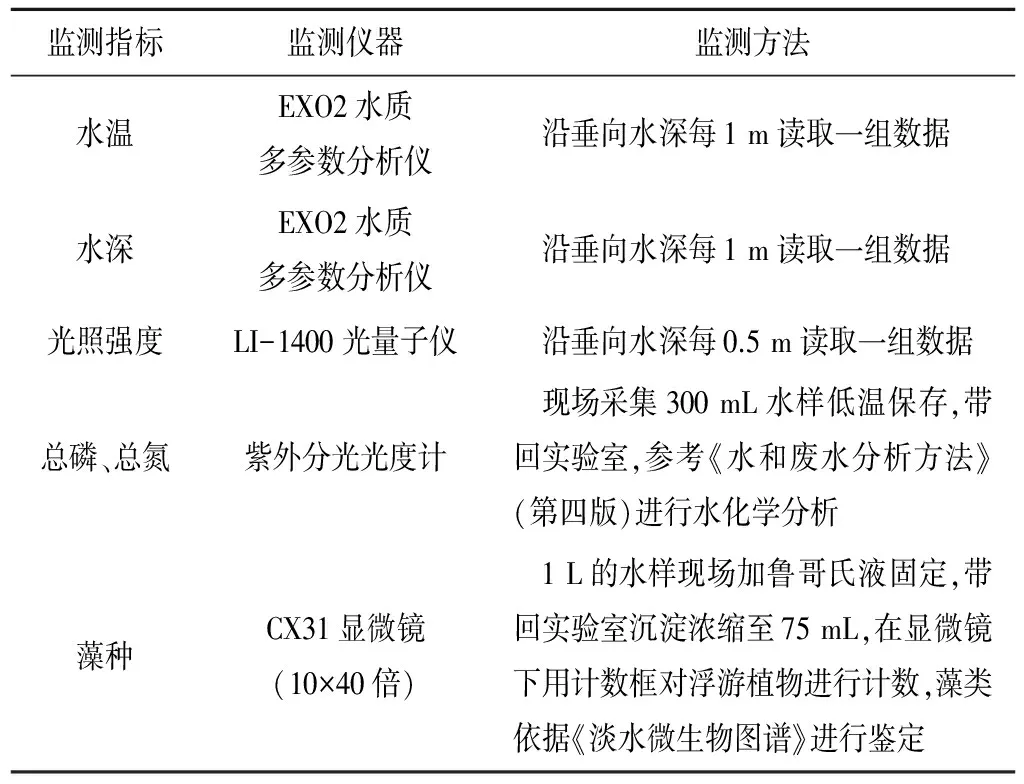
表2 监测指标及相应方法Tab.2 Monitoring index and relevant method
2 结果与分析
2.1 大宁河浮游植物细胞密度
2015-2016年大宁河浮游植物细胞密度呈现显著的时空异质性(见表3),总细胞密度在0.664×105~54.848×105cell/L之间变化,最大值出现在8月份的DN04,最小值出现在11月份的DN05。浮游植物总细胞密度年内分配不均,11月份和1月份库湾平均细胞密度分别为0.959×105cell/L和4.764×105cell/L,整体处于较低水平,4月份和8月份库湾总细胞密度整体较高且波动较大,空间上水面开阔的DN03、DN04藻细胞密度相比于其他断面高。
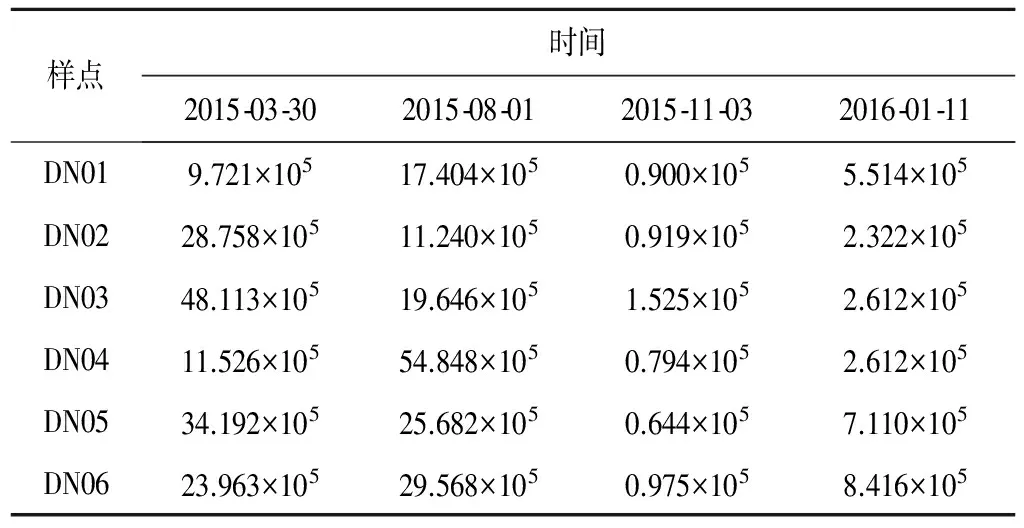
表3 大宁河浮游植物细胞密度的时空变化 cell/L
2.2 大宁河浮游藻类优势藻种及季节演替
本研究中硅藻细胞总密度在春冬两季较高,其数量远超过蓝绿藻(见图2)。在空间分布上,河道狭窄的DN05样点硅藻细胞密度高于其他样点,春季硅藻细胞密度占总细胞密度的89.7%,冬季为76.7%,蓝绿藻则在夏秋两季大量繁殖,蓝藻在夏季和秋季的百分比分别为47.7%和15.7%,绿藻在夏季和秋季的百分比分别为24.5%和50.2%。因此,在本研究中,硅藻为优势藻种时,蓝藻和绿藻变化规律相似,而当蓝藻或绿藻大量繁殖时,蓝绿藻会出现交替占优,说明蓝藻和绿藻之间会出现相互竞争现象。
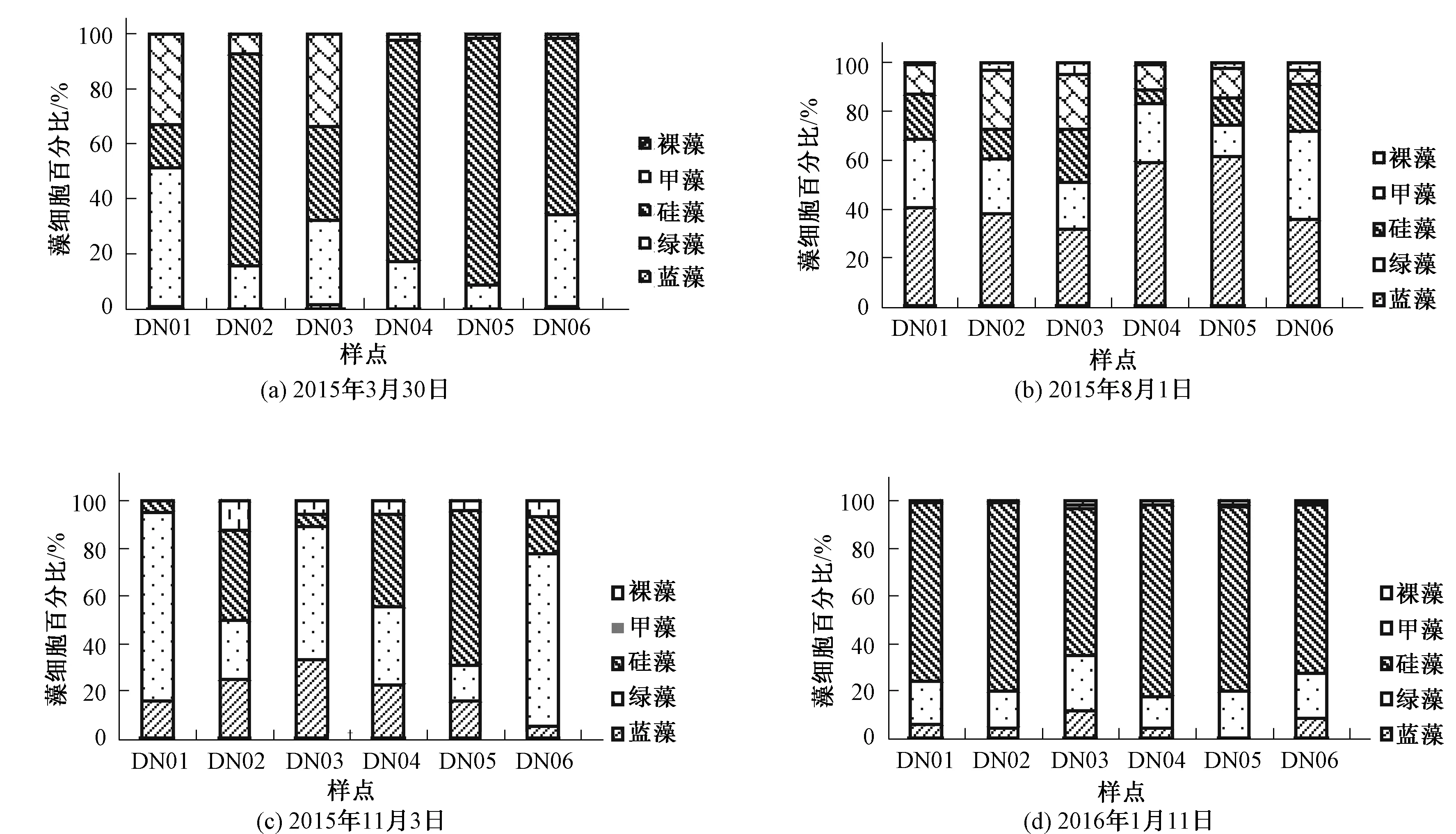
图2 大宁河各藻种细胞密度时空变化Fig.2 Alage cell density changes of different space-time in Daning River
综合大宁河4次采样结果,大宁河水华暴发呈多种复合藻种同时大量增殖态势而少有单一藻种长时间占优现象,大宁河浮游植物优势藻种呈硅藻(小环藻、直链藻)→蓝藻(鱼腥藻、卵囊藻)、绿藻(小球藻)→硅藻(小环藻)的演替规律。
全面从严治党,是以习近平同志为核心的党中央作出的重大战略部署,旨在通过全面增强党要管党、从严治党的自觉,不断提高党的建设质量,不断提高我们党的执政能力和领导水平,持续增强我们党自我净化、自我完善、自我革新、自我提高的能力,把党建设成为始终走在时代前列、人民衷心拥护、勇于自我革命、经得起各种风浪考验、朝气蓬勃的马克思主义执政党。要提升对全面从严治党重要性的认识,必须从战略高度和长远角度重视和推进全面从严治党。
3 讨 论
3.1 大宁河浮游植物季节演替与环境因子的关系
3.1.1 水温的变化
大宁河2015-2016年各采样点表层水温变化如图3所示。各样点表层水温随季节变化规律明显,夏季8月份水温最高,可达31 ℃,其次是秋季11月份,平均温度为21 ℃,春季3月份水温为17 ℃,冬季1月份水温最低,为16 ℃。
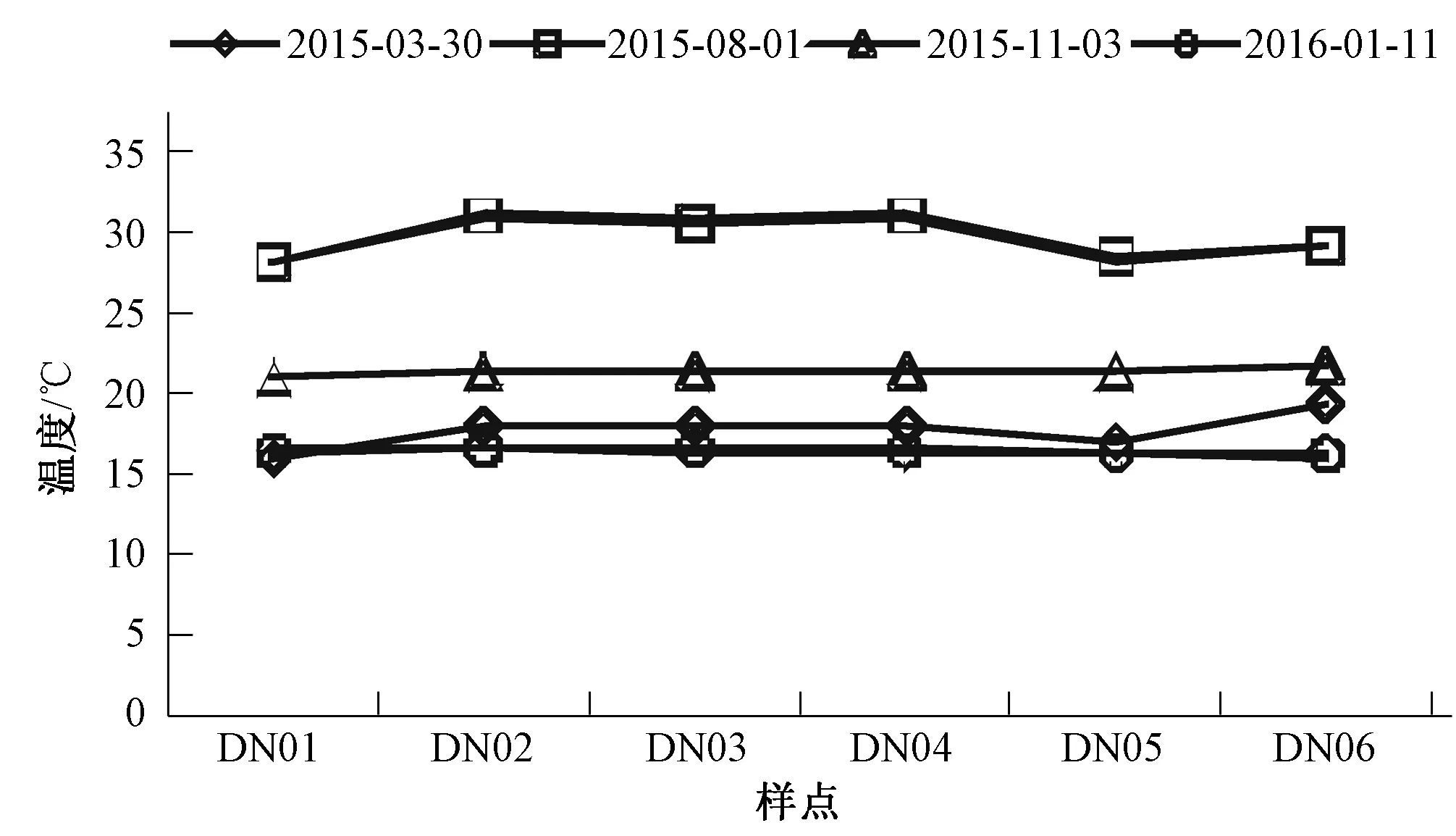
图3 大宁河各样点表层水温变化趋势Fig.3 The variation of the surface water temperature in different sampling stations in Daning River
3.1.2 真光层和混合层深度的变化
真光层深度Zeu按照Beer-Lambert原理,取表层PAR的1%的PAR对应水深以上区域[10,11],计算公式即为:
Zeu=1/Kdln (1/100)
(1)
式中:Kd为水下光衰减系数,m-1,根据Beer-Lambert原理计算得到。
混合层Zmix根据Montegut[12]提出的计算方法得到,按与表层水温首次相差0.5 ℃水温对应的水深计算。大宁河各样点真光层与混合层深度比较如图4所示。
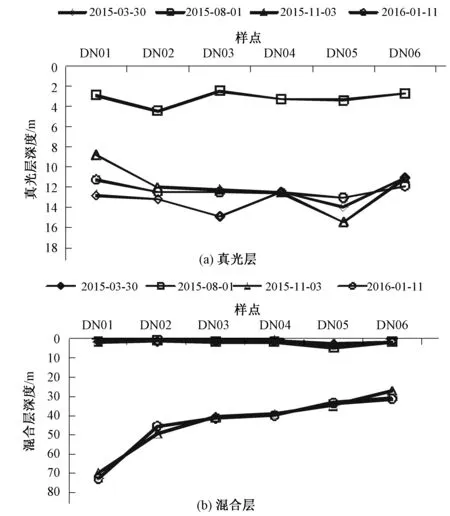
图4 大宁河真光层和混合层深度比较图Fig 4 The comparison plot of euphotic and mixing depth in Daning River
由图4(a)可见,在同一时期DN04真光层深度大于其他样点,导致这一变化可能与4号样点河道狭窄有关;河口DN01真光层深度小于其他样点。8月份真光层深度明显小于其他季节,真光层深度位于2.5~4.5 m之间,平均真光层深度为3.2 m;11月和次年1月真光层深度较深,平均真光层深度为12.2 m;3月平均真光层深度为13 m。导致真光层深度季节变化明显的原因是8月份属于汛期,水体中泥沙含量较高,水体透光性较弱,导致真光层深度较小。3月份、10月份和1月份为蓄水期,水体清澈,透光性较好,真光层深度较大。
比较各季度大宁河混合层深度图4(b)可以看出,3月份和8月份混合层深度沿程逐渐加深,在DN04达到最深,最深为4.8 m,DN05混合层深度又变浅;11月和1月混合层深度沿程减小,且在DN01和DN02样点间变幅最大,两者相差20 m,可能与1号样点位于河口,与长江干流水体掺混剧烈,故混合层深度较深。3月和8月混合层深度均较小,平均混合层深度分别为1.7和2.0 m;11月份和1月份混合层深度较深,最大可达73 m。
将2015年大宁河各样点优势藻种硅藻和浮游植物总细胞密度与环境因子温度、真光层深度、混合层深度做相关性分析,结果如表4所示,硅藻细胞密度与混合层深度呈极显著正相关(R=0.567,P=0.004);蓝藻细胞密度与温度呈极显著正相关(R=0.682,P<0.01),与真光层深度呈极显著负相关(R=-0.605,P=0.02);绿藻细胞密度与温度呈极显著正相关(R=0.627,P=0.001),与真光层深度(R=-0.549,P=0.05)和混合层深度(R=-0.565,P=0.004)呈极显著负相关;总细胞密度与混合层深度呈极显著负相关(R=-0.678,P=0)。
浮游植物总细胞密度与温度、真光层深度均无显著相关性,与混合层深度呈极显著负相关。陈洋等研究发现藻类净初级生产力与混合层Zmix存在负相关,即混合层深度越小藻类净初级生产力越大[13]。在大宁河浮游植物分析中,春夏季混合层深度较小,且此时藻类大量繁殖,与上述研究混合层深度越小藻类生产力越大相符。蓝藻的生长受温度的影响很大,在适宜的温度范围内,温度每升高10 ℃,蓝藻类酶促反应速率提高1~2倍,其代谢速率和生长速率可相应提高[14],蓝藻大量繁殖通常发生在夏季气温和水温都较高的时段,本研究中夏季蓝藻大量繁殖,且与温度呈极显著正相关。绿藻的生长也与温度呈显著正相关。
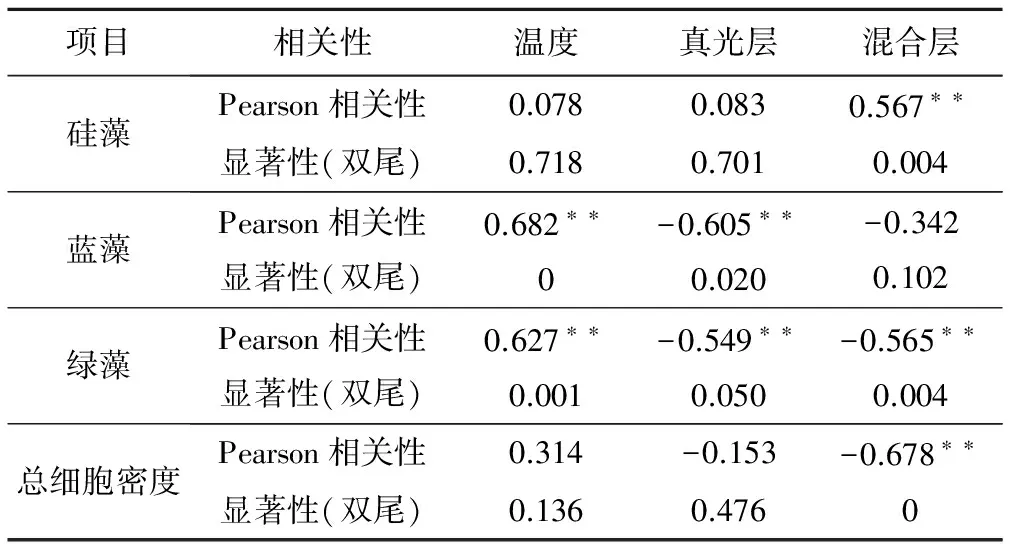
表4 大宁河硅藻细胞密度和总细胞密度与各环境因子的关系Tab.4 Correlation coefficients between phytoplankton abundance and environmental factors in Daning River
注:*表示差异性显著(P<0.05);**表示差异性极显著(P<0.01);N=24。
3.2 大宁河浮游植物与营养盐条件的关系
3.2.1 营养盐(氮、磷、硅)变化
从图5中可以看出,大宁河各样点营养盐浓度在空间上无明显变化规律,但季节变化明显。总氮浓度在1月份高于其他月份,平均浓度达到2.4 mg/L,而其他3个月份总氮浓度较低,平均浓度为1.3 mg/L。总磷浓度除个别样点外(DN03),全年浓度较低,平均浓度仅有0.074 mg/L。氮磷比冬季1月份大于其他季节,TN/TP达35∶1,秋季氮磷比为26∶1,夏季平均氮磷比为14∶1,春季氮磷比为20∶1。水体中硅酸盐浓度在冬季1月份达到最大,其平均浓度为9 mg/L,其他月份硅浓度相差不大,平均浓度为7 mg/L。
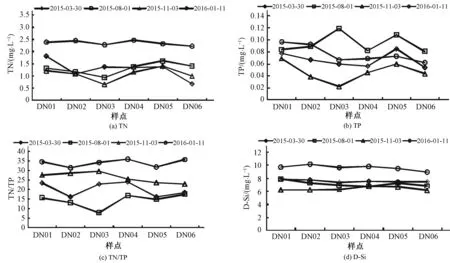
图5 大宁河各季度营养盐(氮、磷、硅)变化图Fig 5 Seasonal variation of nutrients(N,P and Si) in Daning River
3.2.2 浮游藻类与营养盐条件的关系
营养盐是浮游藻类生长不可或缺的环境条件,其关系十分密切,营养盐含量的变化可以影响浮游藻类的生长,而藻类的生长繁殖又可能会导致营养盐含量的变化[15]。氮磷浓度比值与藻类增殖有密切关系。据有关文献:日本学者认为当氮磷比在10∶1~25∶1范围时,藻类生长与氮、磷浓度存在直线相关关系;氮磷比在12∶1~13∶1时最适宜藻类生长;中国环境科学院刘玉生等人提出:氮磷比为10∶1时,微囊藻比增殖速率接近最大值,比值为10∶1~15∶1时生长率最大。氮磷比也影响藻类种群组成,较高氮磷比容易形成蓝藻水华[16]。比较图5(c)可以看出春季大宁河的氮磷比在10∶1~25∶1范围内,藻类生长与氮磷浓度呈正相关,春季大宁河藻类大量繁殖。夏季平均氮磷比为14∶1,此时藻类生长率最大,藻细胞密度较其他季节高;秋冬季各样点氮磷比浓度远高于25∶1,氮磷比较高反而抑制了藻类的生长繁殖,此时表层水体藻细胞浓度较春夏两季少。许多研究表明,硅酸盐浓度的增加是导致硅藻生长的重要因素[17,18]。由图5(d)可以看出,冬季1月份硅酸盐浓度最高,而此时硅藻为优势藻种,说明硅酸盐浓度的升高对于硅藻的生长具有促进作用。
3.3 大宁河浮游植物与水动力条件的关系
水动力条件变化引起的藻类群落结构变化,主要变现为水体混合加剧导致优势种群的转换[19]。Huisman[20]通过人工混合搅动Nieuwe Meer湖,研究混合搅动对上浮型和下沉型浮游植物种类光照竞争的影响,结果表明:混合扰动可能导致浮游植物不同种类之间产生对光照的竞争,并导致湖中浮游植物各类组成的变化,在低扰动扩散条件下,上浮型的有毒藻----微囊藻成为优势种,而硅藻在高扰动扩散时成为优势种。大宁河由于旅游业的发展,其中快艇往来频繁,故表层水体掺混剧烈,由Huisman的研究可以看出,硅藻在高扰动水体中成为优势种,说明行船扰动对于春冬季硅藻的生长繁殖起到了一定的促进作用。
浮游植物通常只有在表层水体中才能获得光照进行繁殖,虽然一些浮游植物可以通过浮力或者游泳保持在表层水体,但大部分浮游植物由于重力作用发生一定沉降。因此可通过水体垂向掺混,将位于下层水体中的浮游植物带入表层,接受光照从而进行繁殖[21]。在河水相对静止的河流中,扰动使得水体温跃层被打破是硅藻水华暴发的一个重要成因。另一方面,由于硅藻自身密度一般大于水体密度,因此在水体中具有下沉趋势,而水体的扰动可以减缓这一趋势[22,23]。大宁河由于强烈的行船扰动,使得密度较大的硅藻细胞减缓了下沉的趋势,更有利于硅藻在上层水体进行光合作用。同时扰动可以使水体掺混剧烈,混合层深度加深(见图4),与硅藻细胞密度和混合层深度呈显著正相关这一结论也是相对应的。说明大宁河中强烈的行船扰动不仅使得表层水体的水动力条件发生改变,而且也使混合层深度加深,而这两方面的因素均对硅藻的生长繁殖具有促进作用。
4 结 语
(1) 大宁河的水华暴发情势呈现显著的季节性演替,水华暴发呈多种复合藻种同时大量增殖态势而少有单一藻种长时间占优现象,大宁河浮游植物优势藻种呈硅藻(小环藻、直链藻)→蓝藻(鱼腥藻、卵囊藻)、绿藻(小球藻)→硅藻(小环藻)的演替规律。
(2) 大宁河独特的水动力条件对硅藻的生长有一定的促进作用,同时行船扰动使得混合层深度加深,混合层深度与硅藻的生长呈正相关,在一定混合层深度以内,混合层深度越大,越有利于硅藻的生长。
(3)水温是影响大宁河蓝藻季节演替最显著的因子,蓝藻的生长与温度呈显著正相关,在一定温度范围内,温度越高,越有利于蓝藻的生长。绿藻的生长是多种环境条件共同作用的结果。
□
[1] 刘德富,黄钰铃,纪道斌,等.三峡水库支流水华与生态调度[M].北京:中国水利水电出版社,2013.
[2] 邓春光.三峡库区富营养化研究[M].北京:中国环境科学出版社,2007.
[3] 钟成华,幸治国,赵文谦,等. 三峡水库蓄水后大宁河水体富营养化调查及评价[J]. 灌溉排水学报,2004,(3):20-23.
[4] Zeng H, Song L R, Yu Z G, et al. Distribution of phytoplankton in the Three-Gorge Reservior during rainy and dry seasons [J].Science of the Total Environment, 2006,367:999-1 009.
[5] 郑丙辉,曹承进,秦延文,等.三峡水库主要入库河流氮营养盐特征及其来源分析[J].环境科学,2008,29(2):310-315.
[6] 曹承进,秦延文,郑丙辉,等.三峡水库主要入库河流磷营养盐特征及其来源分析[J].环境科学,2008,(2):2 310-2 315.
[7] Zhang JL, Zheng BH, Liu LS, et al. Seasonal variation of phytoplankton in the Daning River and its relationships with environmental factors after impounding of the Three Gorges Reservoir: a four-year study[J]. Procedia Environ Sci, 2010,(2):1 479-1 490.
[8] 张佳磊,郑丙辉,刘录三,等.三峡水库试验性蓄水前后大宁河富营养化状态比较[J].环境科学,2012,(10):3 382-3 389.
[9] 张佳磊,郑丙辉,熊超军,等.三峡大宁河水体光学特征及其对藻类生物量的影响[J].环境科学研究,2014,(5):492-497.
[10] Khanna D R, Bhutiani R, Chandra K S. Effect of the euphotic depth and mixing depth on phytoplanktonic growth mechanism[J]. International Journal of Environmental Research, 2009,3(2):223-228.
[11] Sakshaug E, Bricaud A, Dandonneau Y, et al. Parameters of photosynthesis: definitions, theory and interpretation of result [J]. Journal of Plankton Research,1997,19(11):1 637-1 670.
[12] Clement D M, Gurvan M, Aibert S F, et al. Mixed layer depth over the global ocean: an examination of profile data and a profile-based climatology [J].Journal of Geophysical Research: Ocean,2004,109(C12):1-20.
[13] 陈 洋,杨正健,黄钰铃,等. 混合层深度对藻类生长的影响研究[J]. 环境科学,2013,(8):3 049-3 056.
[14] 陈 玲,赵建夫.环境监测[M].北京:化学工业出版社,2004:373-375.
[15] 周小玉. 千岛湖浮游藻类演替格局与环境因子的关系及其机理的初探[D]. 上海:上海海洋大学,2012.
[16] 孔繁翔,高 光.大型浅水富营养化湖泊中蓝藻水华形成机理的思考[J].生态学报,2005,25(3):589-595.
[17] Habib OA, Tippett R, Murphy KJ. Seasonal changes in phytoplankton community structure in relation to physico-chemical factors in Loch Lomond, Scotland[J]. Hydrobiologia, 1997,350:63-79.
[18] Hutchins DA, Bruland KW. Iron-limited diatom growth and Si:N uptake ratios in a coastal upwelling regime[J]. Nature, 1998,393:561-564.
[19] 吴晓辉,李其军. 水动力条件对藻类影响的研究进展[J]. 生态环境学报,2010,(7):1 732-1 738.
[20] Huisman J, Sharples H, Stroom J M, et al.Changes in turbulent mixing shift competition for light between phytoplankton species[J].Ecology,2004,85(11):2 960-2 970.
[21] Reynolds C S. Ecology of phytoplankton[M].Cambridge:Cambridge University Press,2006:38-49.
[22] Huisman J, Jonker R R, Zonneveld C, et al. Competition for light between phytoplankton species: experimental tests of mechanistic theory [J].Ecology,1990,80(1):211-222.
[23] Chisholm S W.WhatLimits Phytoplankton Growth?[J].OCEANUS,1992,35:36-46.
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
小天使·三年级语数英综合(2022年3期)2022-04-17
当代水产(2021年8期)2021-11-04
潍坊学院学报(2020年2期)2021-01-18
环境科学导刊(2021年2期)2021-01-09
军事文摘(2020年20期)2020-11-16
黑龙江水利科技(2020年8期)2020-01-12
当代水产(2019年8期)2019-10-12
当代水产(2019年9期)2019-10-08
