
辐照育种杂交水稻Ⅱ优838的耐热性鉴定
2017-03-17 15:04王艳高鹏伏毅何江孙群黄敏
湖北农业科学 2016年23期
王艳+高鹏+伏毅+何江+孙群+黄敏
摘要:采用田间和实验室耐热鉴定相结合的方法对水稻Ⅱ优838的耐热性进行了系统鉴定。结果表明,在自然高温胁迫下,Ⅱ优838的剑叶叶绿素含量、结实率和产量比其他9个参试水稻品种都高;在实验室人工气候箱高温胁迫下,Ⅱ优838的剑叶叶绿素含量与超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)的活性比父本辐恢838受影响小。由此可知,Ⅱ优838对高温具有较好的耐受性,且在耐热性上父本辐恢838的贡献大于母本。
关键词:杂交水稻;Ⅱ优838;耐热性;结实率;叶绿素含量;抗氧化酶
中图分类号:S511;Q945.78 文献标识码:A 文章编号:0439-8114(2016)23-6061-06
DOI:10.14088/j.cnki.issn0439-8114.2016.23.012
Abstract: A set of field-based identification and laboratory analysis were performed to identify heat resistance of Ⅱ YOU 838. The results showed that chlorophyll content in flag leaf, seed setting rate and yield were the highest in Ⅱ YOU 838 compared with all other nice rice varieties tested under natural high temperature stress. The laboratory analysis showed that the chlorophyll content, SOD, POD, and CAT antioxidant enzyme activities were less affected by heat stress in Ⅱ YOU 838 compared with its paternal under high temperature stress in artificial climate chamber. Accordingly, Ⅱ YOU 838 had superior heat resistance, and its male parent contributed to this characteristic at a higher degree than its female parent.
Key words: hybrid rice; Ⅱ YOU 838; heat resistance; seed setting rate; chlorophyll content; antioxidant enzymes
水稻(Oryza sativa L.)是中國主要的粮食作物,其产量的高低直接影响到中国的社会稳定和粮食安全。近年来,极端高温气候常出现在水稻对高温敏感的抽穗开花期或灌浆期,使水稻遭受高温热害,导致大面积减产[1]。开展水稻耐热性研究,用以指导分子育种,加速选育耐热性强的优良品种是战胜高温自然灾害的根本措施。中国水稻耐热性育种起步较晚,虽已陆续开展水稻的耐热性研究,但因遗传分析群体一般不是育种群体,而育种群体常常又未能进行常规的遗传分析,使得耐热性遗传分析结果对水稻耐热性育种的指导有限[2]。因此,通过育种方法筛选出的耐热品种大多农艺性状不佳,难于直接利用。若采用已被广泛种植的高产稳产且农艺性状佳的种质资源来进行遗传分析,挖掘耐热基因用于遗传改良,培育出有育种价值的耐热水稻新品种,是能克服水稻耐热的遗传分析与育种工作相脱离这一问题的有效策略。
籼稻的耐热性优于粳稻,在籼稻中进行耐热性筛选将有更大的机会获得耐热性强的稻种资源[3]。籼稻Ⅱ-32A与辐恢838配组育成的杂交中稻组合Ⅱ优838产量稳定、耐受性好、农艺性状佳,其制种方法已获国家专利[4,5]。Ⅱ优838是第一个在四川省育种攻关项目中连续两年区域试验结果都比汕优63增产5%以上的组合,先后被列入四川省增粮工程项目和国家科技成果重点推广计划项目[6,7]。Ⅱ优838也是中国出口东南亚地区数量最多的水稻品种。随着在高温区域的大面积推广,Ⅱ优838耐高温热害的优点更加突显,成为水稻耐热研究的理想材料。目前,对Ⅱ优838的耐热性研究都停留于田间试验对其产量的研究,未见有对其耐热性进行系统鉴定和遗传分析的报道[8,9]。本研究对Ⅱ优838分别进行了田间和实验室耐热性鉴定,并分析了高温胁迫对Ⅱ优838及其亲本的相关性状影响,总结了Ⅱ优838耐热的遗传规律,旨在为水稻耐热性育种提供基因资源。
1 材料与方法
1.1 供试材料
田间耐热性鉴定试验共选用10个水稻品种:Ⅱ优838、辐恢838、Ⅱ-32B(恢复育性的Ⅱ-32A)由四川省原子能研究院供种;绵恢725、冈优725由绵阳市农业科学院供种;珍汕97B、明恢63、冈46B、Ⅱ优63由四川益邦种业有限责任公司供种;汕优63由新津县种子公司供种。这10个品种中包含了4组亲子代组合,其中Ⅱ优838是杂交子代,辐恢838是其父本,Ⅱ-32B是其母本;汕优63是子代,明恢63是其父本,珍汕97B是其母本;冈优725是子代,绵恢725是其父本,冈46B是其母本;Ⅱ优63是杂交子代,明恢63是其父本,Ⅱ-32B是其母本。
实验室耐热性鉴定试验采用Ⅱ优838及其父本辐恢838为材料。
1.2 试验方法与处理
在重庆市丰都县高家镇后池坝村的自然高温区域选取实验田进行田间耐热性鉴定,采用分期播种方法,从2011年3月10日至5月20日,每月10、20、30日播种,共8个播期。稻株采用单本栽插,每批每个材料移栽5行,每行12株,共60株为一个小区,株行距25.0 cm×13.3 cm。试验采用随机区组设计,共设3次重复。
实验室耐热性鉴定采用的Ⅱ优838和辐恢838都用常规方法播种并育秧于四川省原子能研究院水稻试验基地,待生長至孕穗期,选取长势一致的Ⅱ优838和辐恢838各12株,带土移栽入盆钵中,每盆3株,随机分为热处理组和对照组,每组2盆。在田间继续培养数日,开花当日将热处理组和对照组分别移入日平均气温37 ℃(8:00~19:00为42 ℃;19:00至第2天8:00为32 ℃)和日平均气温26 ℃(8:00~19:00为32 ℃;19:00至第2天8:00为22 ℃)的光照培养箱[型号PGX-300B,容积量320 L,参数设置为光照强度600 μmol/(m2·s),光暗循环11 h/13 h,湿度80%]中生长。两组的水肥等其他条件相同。在处理前1 d及热处理的第1、3、5、7天的19:00分别采集Ⅱ优838和辐恢838的热处理组和对照组剑叶,液氮冷冻保存用于叶绿素含量及抗氧化保护酶活性测定。采自不同稻株的3个剑叶作生物学重复。
1.3 农艺性状考察与气象数据记录
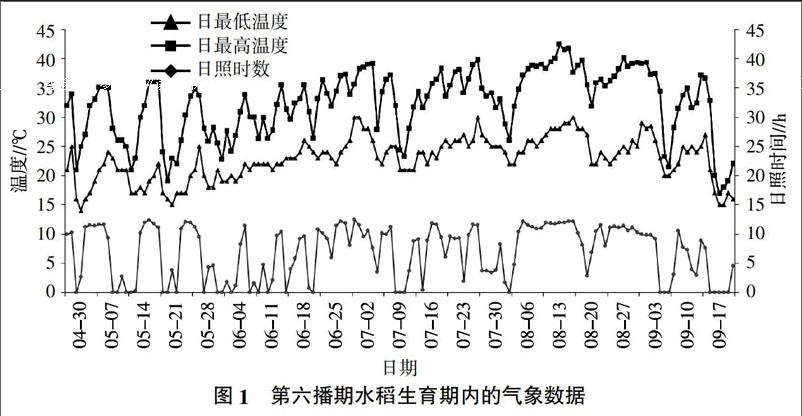
通过记录的气象数据发现第六播期(即4月30日播种)的各水稻品种在热敏感的抽穗开花和灌浆期正好遭遇持续高温天气,因此取第六播期内的各水稻品种进行农艺性状考察。从第六播期每小区中部选取10株单株,在水稻成熟后记录其株高、剑叶长、剑叶宽,再在收割晒干后,室内统计结实率和小区产量(平均单株产量×60株)。
2011年8月l7日(当日最高温度41.5 ℃)采集正处于抽穗开花期的各水稻品种剑叶,用于测定叶绿素含量。每个品种都选取不同的3株水稻植株作为生物学重复。
1.4 生理生化指标测定
叶绿素含量测定参照杨敏文[10]的方法。剑叶经液氮碾磨后,按质量体积比10%加入PBS,涡旋1 min后,在4 ℃下3 000g离心10 min获得酶粗提液,分别用购自南京建成生物工程研究所的超氧化物歧化酶(SOD)测试盒(目录号:A001-1)、过氧化氢酶(CAT)测试盒(目录号:A007)和过氧化物酶(POD)测试盒(目录号:A084-3)完成SOD、CAT和POD酶活性测定。各指标测定值均为新鲜叶片含量。
1.5 数据分析方法
数据用SPSS Statistics 17.0和Excel 2007软件进行分析和绘图。试验数据采用单因素方差分析的最小显著差法(LSD法)进行差异显著性分析。
2 结果与分析
2.1 Ⅱ优838的田间耐热性鉴定结果
2.1.1 遭遇自然高温胁迫情况 根据重庆市丰都县气象局提供的2011年气象记录资料,第六播期的各水稻品种生育期内(4月30日至9月17日)达到水稻花期高温不育危害指标35 ℃及以上温度的时间有59 d,平均最高温度达到了32.4 ℃,且8月16日出现42.4 ℃的最高日最高温度。2011年8月 8日至22日和8月24日至9月6日分别是两个持续高温时段,每天的最高气温都达到了35 ℃以上,且除3 d外每天的日照时间都超过了9 h。第六播期的各水稻品种在这两个高温时段内正好处于热敏感的抽穗开花期和灌浆期,完整的遭遇了自然高温胁迫,因此选第六播期内的各水稻品种进行田间耐热性鉴定。图1记录了第六播期水稻生育期内的温度和日照时间。
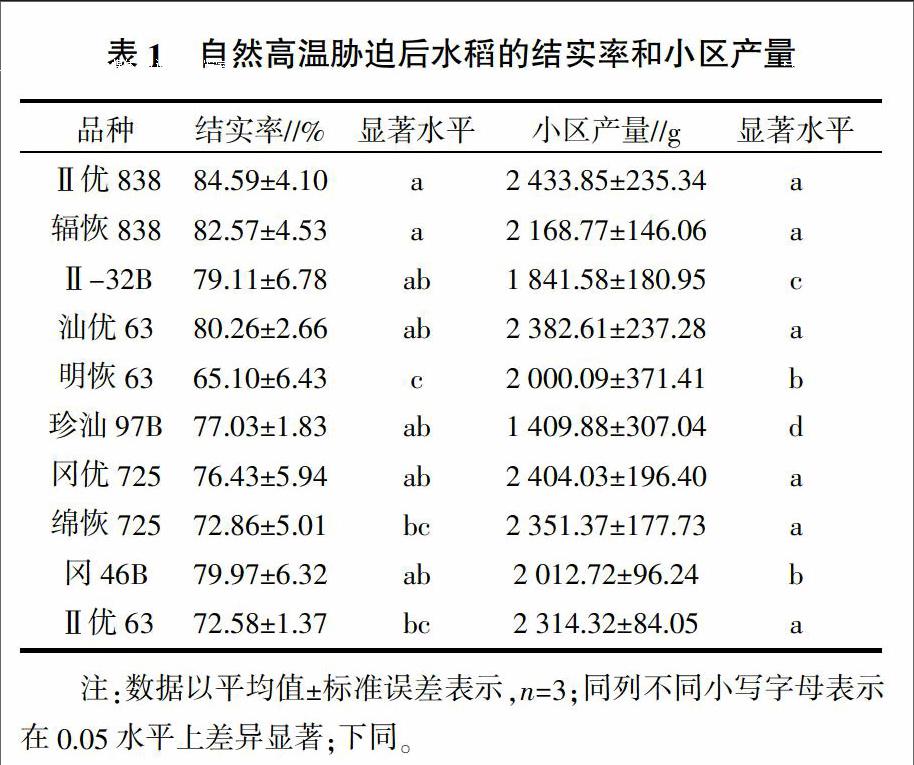
2.1.2 高温胁迫下水稻结实率和小区产量 由表1可以看出,第六播期的10个参试水稻品种在遭遇自然高温胁迫后,Ⅱ优838的结实率是最高的,其余水稻品种的结实率由高到低依次为辐恢838、汕优63、冈46B、Ⅱ-32B、珍汕97B、冈优725、绵恢725、Ⅱ优63、明恢63。遭遇自然高温胁迫后,Ⅱ优838的小区产量是10个水稻品种中受影响最小的,且Ⅱ优838和父本辐恢838的小区产量都显著高于母本Ⅱ-32B(P<0.05)。其他组合中母本水稻的小区产量也都显著低于其子代和父本恢复系的小区产量(P<0.05)。
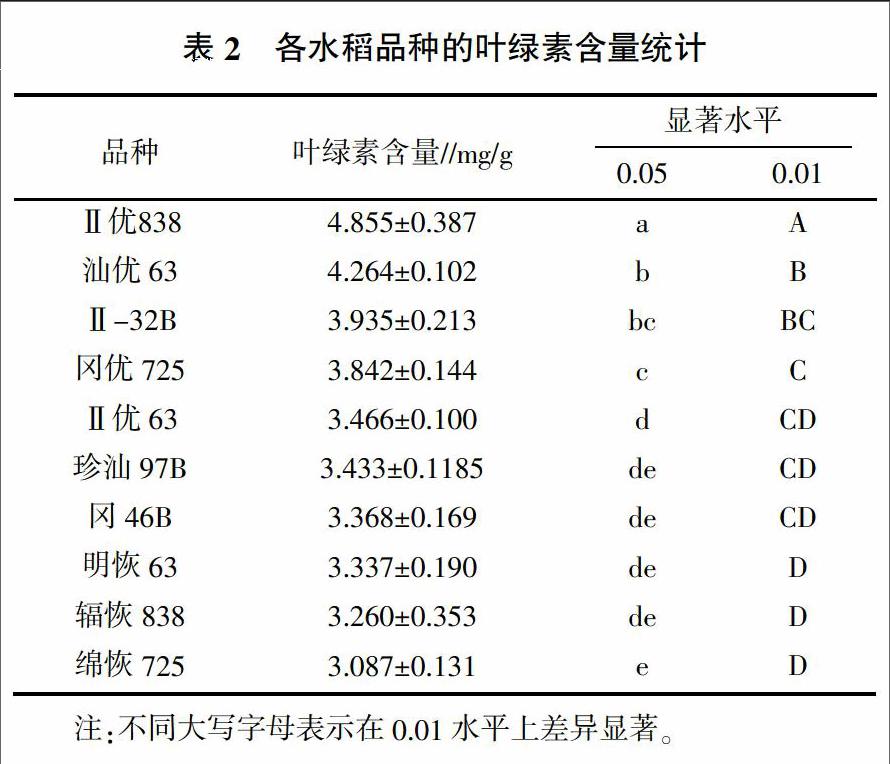
2.1.3 高温胁迫下开花期水稻剑叶叶绿素含量 8月17日,除冈46B和珍汕97B的第七播期和第八播期正好处于抽穗开花期外,其余水稻品种的第六播期正好处于抽穗开花期。各水稻品种抽穗开花期剑叶的叶绿素含量和差异显著性分析结果见表2,在高于40 ℃的自然高温下,Ⅱ优838的剑叶叶绿素含量最高,与其他品种间存在极显著差异(P<0.01)。
2.1.4 高温对结实率的影响 根据水稻幼穗分化和开花灌浆的生育时期特点,将相应阶段的抽穗前15 d的活动积温(12.0 ℃作为起点温度)定义为自变量a,抽穗后15 d的活动积温定义为自变量b,抽穗后16~30 d的活动积温定义为自变量c,抽穗后15 d高于27.5 ℃的天数定义为自变量d,抽穗后15 d高于27.5 ℃的积温定义为自变量e;将Ⅱ优838及其亲本各播期结实率的平均值作为因变量(y)进行回归分析(结实率参见已发表数据[11]),结果见表3。从表3可以看出,Ⅱ优838与父本辐恢838的结实率都与各自抽穗后15 d的活动积温存在显著的线性回归关系(P<0.05),母本Ⅱ-32B的结实率与其抽穗后15 d的活动积温存在极显著的线性回归关系(P<0.01);而Ⅱ优838与两个亲本的结实率都与各自在抽穗前15 d和抽穗后16~30 d的活动积温没有显著的线性相关,表明抽穗后15 d是Ⅱ优838与两个亲本对温度最敏感的阶段,如果在抽穗后15 d遭遇高温会对Ⅱ优838及其亲本的结实率产生很大的影响。Ⅱ-32B对抽穗后15 d高于27.5 ℃的时间和高于27.5 ℃的积温最敏感,与结实率间都存在极显著的负相关,且存在0.01水平上的线性回归关系。Ⅱ优838和辐恢838的结实率受高温的影响不如Ⅱ-32B明显,表现出一定的耐受性。
2.1.5 Ⅱ优838与其亲本性状的相关性分析 对第六播期的Ⅱ优838及其亲本的株高、剑叶长、剑叶宽和结实率进行相关性分析。Pearson相关系数和显著性差异结果表明,Ⅱ优838的株高与父本辐恢838的株高在0.05水平上呈显著正相关,相关系数r=0.38。Ⅱ优838的剑叶宽与父本辐恢838的剑叶宽呈极显著正相关(r=0.78,P<0.01),与母本Ⅱ-32B的剑叶宽呈显著正相关(r=0.40,P<0.05)。Ⅱ优838的结实率与其剑叶宽在0.01水平上呈极显著负相关,r=0.72。Ⅱ优838的结实率与其父本和母本的结实率呈极显著正相关(r分别为0.55和0.53,P<0.01)。相同热胁迫条件下,Ⅱ优838的性状受影响程度跟父本辐恢838更相似些。
2.2 Ⅱ优838的人工气候箱耐热性鉴定结果
2.2.1 高温胁迫对剑叶叶绿素含量的影响 光合作用是水稻遭受高温热害最敏感的过程之一,叶片叶绿素含量常作为反映光合效率的重要指标[12,13]。由图2可知,在热胁迫第1天时,Ⅱ优838与辐恢838的剑叶叶绿素含量都增大,Ⅱ优838的高温处理组与对照组间具有显著差异(P<0.05)。随着高温处理时间的增加,叶绿素含量开始持续下降,在高温处理第7天时降幅最大,Ⅱ优838下降了15.4%,辐恢838下降了22.6%。相同条件下,Ⅱ优838的叶绿素含量下降幅度比辐恢838小。辐恢838从高温处理第3天开始高温处理组的叶绿素含量就已显著低于对照组(P<0.05),而Ⅱ优838从第5天开始高温处理组的叶绿素含量才显著低于对照组(P<0.05)。
2.2.2 高温胁迫对剑叶抗氧化酶活性的影响 SOD是植物体内清除活性氧自由基的关键酶,其活性的强弱与植物抗氧化能力密切相关[14]。由表4可以看出,高温处理后Ⅱ优838和辐恢838剑叶中SOD活性都增高,而对照组在7 d中SOD活性没有明显差异。高温处理第7天时,Ⅱ优838的高溫处理组SOD活性增幅最大,且与对照组具有极显著差异(P<0.01)。
POD是植物体内负责清除H2O2的重要酶类,可催化H2O2转化为H2O,使细胞受到抗氧化保护[15]。高温胁迫后,Ⅱ优838和辐恢838的剑叶POD活性都增大。热胁迫第3天时,Ⅱ优838的POD活性增幅最大,与对照组差异显著(P<0.05),且比辐恢838高温处理组高51%。高温处理7 d中,Ⅱ优838和辐恢838对照组的剑叶POD活性都很稳定,样品间差异不明显。
CAT是植物体内的活性氧清除酶,CAT和H2O2具有较高的亲和力,主要清除线粒体电子传递和脂肪酸氧化中产生的H2O2。高温处理后,Ⅱ优838和辐恢838的CAT活性都降低,但Ⅱ优838的CAT活性仍都高于辐恢838。高温处理第3天时,CAT活性的下降幅度最大,Ⅱ优838与对照相比下降了30%,辐恢838与对照相比下降了41%。高温处理7 d中,Ⅱ优838和辐恢838对照组剑叶的CAT活性变化不大。
3 小结与讨论
重庆市丰都县的自然高温条件达到了对水稻田间耐热性检验的要求。田间耐热性鉴定试验中,第六播期的各水稻品种在热敏感的抽穗开花期和灌浆期正好每天都遭遇了日最高气温35 ℃以上的持续高温胁迫,Ⅱ优838与其他9个参试水稻品种相比,胁迫后结实率和产量都最高,表明在田间遭遇自然高温后Ⅱ优838能表现出优于其他水稻品种的耐受性,产量受高温影响最小。相同自然高温胁迫下,Ⅱ优838的结实率和小区产量都高于父本辐恢838,且父本的结实率和小区产量又都高于母本Ⅱ-32B。有研究也发现Ⅱ优838具有很好的耐热性,其耐热性优于包括汕优63在内的许多参试水稻品种,且各杂交水稻组合的耐高温能力与其父本的耐高温能力呈显著的线性正相关[16-18]。目前,水稻耐热性鉴定并无统一标准,又因水稻材料、鉴定时期、热处理方式和评价标准等的不同,研究结果很难统一[19]。胡声博等[20]研究认为在38 ℃的人工气候箱内高温处理的3 d中,Ⅱ优838开花期处于高温时颖花的受精率受热胁迫影响很大。本研究发现,Ⅱ优838无论在田间自然高温胁迫后还是在42 ℃的人工气候箱高温胁迫后,都能有好的耐受性表现,推测在受高温胁迫过程中Ⅱ优838自身可能存在一种有效的避害自卫方式,虽然高温会影响Ⅱ优838的受精率,但其自身的有效避热防御措施能使它最终保持较高的结实率。植物的耐热性表现分为耐受和避害两种方式。遭遇高温胁迫后,Ⅱ优838的成功避害也是其耐热的一种表现。
光合效率降低通常被认为是光合作用对高温胁迫响应的主要表现,而叶绿素含量是反映植物光合效率高低的重要生理指标[21]。在自然高温胁迫下,Ⅱ优838剑叶叶绿素含量极显著高于其他水稻品种(P<0.01),能保持较高的光合效率。剑叶是重要的功能叶,在水稻生育期内的大部分时间都具有光合同化作用,其性状能在一定程度上影响结实率[22]。Ⅱ优838的结实率与亲本的结实率呈极显著正相关,且与父本辐恢838的相关系数更高。Ⅱ优838的剑叶宽与亲本的剑叶宽呈显著正相关,且与父本辐恢838的相关性高于母本Ⅱ-32B。水稻生育期内的活动积温与8个播期的结实率的回归分析结果表明,抽穗后15 d是Ⅱ优838与两个亲本对温度最敏感的阶段,而Ⅱ优838和父本辐恢838的结实率受高温胁迫影响程度比母本Ⅱ-32B更小,表现出一定的耐热性。综上所述,推测父本辐恢838对杂交子代Ⅱ优838耐热性的贡献更大。
实验室人工气候箱高温胁迫后,Ⅱ优838和辐恢838的剑叶叶绿素含量都降低,但Ⅱ优838的叶绿素含量下降得较少,其光合系统受到高温的影响更小,表现出一定的耐热性。值得关注的一点是,在高温胁迫第1天时,Ⅱ优838与辐恢838的剑叶叶绿素含量都增大,Ⅱ优838的高温处理组与对照组间存在显著差异(P<0.05)。在高温胁迫初期,叶绿素含量的增大有助于维持较高的光合效率,推测这是Ⅱ优838表现出一定耐热性的原因。通过分析不同高温胁迫持续时间下Ⅱ优838抗氧化保护酶活性的变化,进一步了解Ⅱ优838对热胁迫的响应情况,有助于总结Ⅱ优838的耐热机制[23]。SOD、CAT和POD都是植物抗氧化的主要酶系,其活性水平决定了细胞膜脂过氧化反应的程度,一定程度上能反映植物在逆境条件下的耐受性[24]。不同高温胁迫时间下,SOD、CAT和POD的酶活性变化情况与前人研究[21,25]很相似,但也因水稻品种和热处理方式的不同,存在一些差异。在连续7 d的高温胁迫中,两个品种的抗氧化保护酶的活性都出现变化,在前期变化较大,到后期变化都减缓。高温胁迫后,Ⅱ优838的剑叶中SOD、CAT和POD的酶活性明显大于父本辐恢838,表明Ⅱ优838的抗氧化酶促保护系统比辐恢838更为活跃,可以更有效地清除热胁迫产生的活性氧,减轻热胁迫造成的膜脂过氧化伤害,延缓剑叶衰老速度[26]。水稻剑叶功能叶的衰老速度与产量呈正相关,这正好与田间自然高温胁迫后Ⅱ优838产量高于辐恢838的结果相符[11]。相同高温胁迫条件下,Ⅱ优838的抗氧化酶活性更高,可以更好地维持细胞膜的稳定性,而微观结构的稳定性同耐热性是密切相关的,表明Ⅱ优838比辐恢838更适应热胁迫压力,能表现出好的耐受性。这在生理水平上再次证明了Ⅱ优838的耐热性。
田间耐热性鉴定和实验室耐热性鉴定结果都表明Ⅱ优838具有优于包括其父本辐恢838在内的其他参试水稻品种的耐热性,在其耐热性上父本辐恢838的贡献更大。Ⅱ优838遭遇高温热害后仍能保持较高的产量,表现出好的耐热性,也是其一直广受推广的一个原因。
参考文献:
[1] 王志刚,王 磊,林 海,等.水稻高温热害及耐热性研究进展[J].中国稻米,2013,19(1):27-31.
[2] 章元明.植物数量遗传学的建立、发展与展望[J].南京农业大学学报,2012,35(5):19-24.
[3] 杨梯丰,张少红,王晓飞,等.水稻抽穗开花期耐热种质资源的筛选鉴定[J].华南农业大学学报,2012,33(4):585-588.
[4] 邓达胜,广华蓉.杂交稻恢复系辐恢838的选育和利用[J].杂交水稻,1996(4):10-12.
[5] 广华容,邓达胜,杨成明,等.杂交中籼新组合Ⅱ优838[J].杂交水稻,1996(5):31.
[6] 邓文敏,邓达胜,吴万义,等.杂交水稻恢复系辐恢838的选育与应用[J].辐射研究与辐射工艺学报,2005,23(2):125.
[7] 邓达胜,陈 浩,邓文敏,等.水稻恢复系辐恢838及其衍生系的选育和应用[J].核农学报,2009,23(2):175-179.
[8] 黄智敏,成尚廉,李成尧.杂交中稻Ⅱ优838耐高温性分析[J].杂交水稻,1998,13(4):34-35.
[9] 郭 军,顾根宝,范同广,等.中国杂交水稻Ⅱ优838在越南母本直播制种高产技术研究[J].杂交水稻,2005,20(4):33-35.
[10] 杨敏文.快速测定植物叶片叶绿素含量方法的探讨[J].光谱实验室,2002,19(4):478-481.
[11] 苏 鹏,冯 慧,陈 浩,等.杂交水稻在自然高温胁迫下的耐热性研究[J].西南农业学报,2013,26(4):1387-1391.
[12] 张顺堂,张桂莲,陈立云,等.高温胁迫对水稻剑叶净光合速率和叶绿素荧光参数的影响[J].中国水稻科学,2011,25(3):335-338.
[13] 唐 婷,郑国伟,李唯奇.植物光合系统对高温胁迫的响应机制[J].中国生物化学与分子生物学报,2012,28(2):127-132.
[14] UEDA Y,UEHARA N,SASAKI H,et al. Impacts of acute ozone stress on superoxide dismutase(SOD) expression and reactive oxygen species (ROS) formation in rice leaves[J].Plant Physiology and Biochemistry,2013,70:396-402.
[15] SHAH K,NAHAKPAM S. Heat exposure alters the expression of SOD,POD,APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars[J].Plant Physiology and Biochemistry,2012,57:106-113.
[16] 黃智敏,成尚廉,李成尧,等.Ⅱ优838作中稻栽培的结实率与耐高温性状[J].湖北气象,1998(1):21-23.
[17] 曾汉来,卢开阳,贺道华,等.中籼杂交水稻新组合结实性的高温适应性鉴定[J].华中农业大学学报,2000,19(1):1-4.
[18] 雷东阳,陈立云,李稳香,等.杂交水稻抽穗扬花期高温对结实率及相关生理特性的影响[J].杂交水稻,2006,21(3):68-71.
[19] 杨永杰,符冠富,熊 杰,等.高温对水稻的影响及水稻耐热性测评方法研究[J].中国稻米,2012,18(1):39-40.
[20] 胡声博,张玉屏,朱德峰,等.杂交水稻耐热性评价[J].中国水稻科学,2012,26(6):751-756.
[21] 张桂莲,陈立云,张顺堂,等.抽穗开花期高温对水稻剑叶理化特性的影响[J].中国农业科学,2007,40(7):1345-1352.
[22] 王复标,黄福灯,程方民,等.水稻生育后期叶片早衰突变体的光合特性与叶绿体超微结构观察[J].作物学报,2012,38(5): 871-879.
[23] 冯汉青,李 芳,管冬冬,等.两种外源化学物影响PEG胁迫对水稻根系的伤害[J].广西植物,2013(4):439-442.
[24] SAHU S,DAS P,RAY M,et al.Osmolyte modulated enhanced rice leaf catalase activity under salt-stress[J].Advances in Bioscience and Biotechnology,2010,1(1):39-46.
[25] 杜尧东,李键陵,王 华,等.高温胁迫对水稻剑叶光合和叶绿素荧光特征的影响[J].生态学杂志,2012,31(10):2541-2548.
[26] 吴志华,李天会,周旭东.微量元素营养对桉树无性系苗木叶绿素及荧光参数影响[J].广西植物,2013(6):832-838.
猜你喜欢
现代园艺(2016年7期)2017-01-09
农家科技下旬刊(2016年9期)2016-12-15
河北渔业(2015年10期)2015-10-15
