
重金属Cd胁迫对元宝枫光合特性的影响
2017-03-15 17:45:53韦忠刚
天津农业科学 2017年2期
韦忠刚
摘 要:为了研究重金属Cd对元宝枫光合作用的影响,通过对元宝枫水培苗进行不同浓度的Cd胁迫处理,分析元宝枫叶片中叶绿素含量及其净光合速率的变化。结果表明,随着Cd胁迫浓度的增加,元宝枫叶片中叶绿素a、叶绿素b、叶绿素总量显著降低,净光合速率也明显下降;Cd低浓度胁迫下,元宝枫叶片光饱和点变化不大,Cd高浓度胁迫下,元宝枫叶片光饱和点显著降低。重金属Cd胁迫严重阻碍了元宝枫叶片的光合作用。
关键词:元宝枫;叶绿素;光合作用
中图分类号:S792.35 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2017.02.025
Abstract: To study the effect of heavy metal Cd on photosynthesis of Acer truncatum, the chlorophyll content and its net photosynthetic rate (P (subscript n)) in leaves of A. truncatum were analyzed by Cd stress treatment. The results showed that the total chlorophyll a, chlorophyll b and the total chlorophyll contents decreased significantly and leaf net photosynthetic rate (Pn) of leaves decreased with the increase of the stress concentration. Under low concentration stress, the light saturation point of A. truncatum leaves changed little, and the light saturation point of A. truncatum leaves decreased significantly under high concentration stress. The photosynthesis of A. truncatum leaves was seriously hindered by heavy metal stress.
Key words: Acer truncatum;chlorophyll;photosynthesis
元寶枫(Acer truncatum)属槭树科槭树属,是我国特有的树种,分布范围广,变异丰富。元宝枫以其独特的叶形、叶色等特点受到广泛关注,是我国重要的园林绿化树种和彩色叶树种,应用前途广泛[1]。随着社会和经济的发展,重金属污染带来的问题日趋增多,已对人类的健康和生存构成较大的威胁。土壤重金属是指密度在5.0 g·cm-3以上的金属元素,对于植物来说,一些重金属(如Cu,Zn等)是植物生长发育进程中的必需微量元素[2],其缺乏会导致植物产生相应的缺素症,如苹果小叶病、小麦“穗而不孕”、黄瓜“花而不实”等;但当土壤中重金属含量超过植物所能接受的临界值时,就会对植物产生胁迫作用,轻则导致植物体代谢过程发生紊乱,生长发育受到抑制;重则导致植物死亡,改变植物群落结构[3]。光合作用是生物界最基本的物质代谢和能量代谢,重金属胁迫通过抑制植物的光合作用从而改变植物的生长代谢,且抑制程度与胁迫程度呈正相关。徐向华[4]研究发现,商陆成熟叶片叶绿素含量随Mn2+处理浓度的提高而降低;郭智等[5]通过对龙葵幼苗进行Cd胁迫发现,其幼苗叶片净光合速率随Cd2+浓度的增加比对照下降6.01%~61.71%。众多研究表明,重金属胁迫能够影响叶绿素的合成,且随着胁迫浓度的增加,膜系统崩溃,叶绿体球形皱缩,会出现大量的脂类小球,导致叶绿体功能遭到破坏,严重影响植物的生长发育[6]。
本试验通过对元宝枫扦插苗进行不同的Cd胁迫处理,测定其光合生理指标的变化,旨在了解Cd胁迫对元宝枫光合作用的影响,为元宝枫的栽培与引种利用提供参考。
1 材料和方法
1.1 试验材料
供试材料为元宝枫种质资源汇集与优良品种选育课题组选育的元宝枫当年生扦插苗。
1.2 试验方法
试验于2016年8月在山东省泰安市邱家店镇西郭庄基地进行。以元宝枫无性系为材料,取15 cm长的插穗在塑料盒中进行水培,不间断通气,每盒30株,待生根后转至1/2改良Hoagland营养液中,pH值=6.0,每14 d换一次营养液。培养 7 d后,保留2个叶芽,其余抹去,继续培养至28 d。28 d后,选取长势一致的植株用于Cd2+胁迫处理。称取定量的CdCl2·2.5 H2O(分析纯),以1/2改良Hoagland 营养液配制成50,100 mg·L-1的Cd2+培养液,pH值=6.0;对照组为等量的1/2改良Hoagland 营养液。每处理无性系1株,3次重复,胁迫处理7 d后测定相应指标。
1.3 测定项目及方法
1.3.1 叶绿素含量测定 叶绿素含量采用丙酮法进行测定。
1.3.2 光合速率日变化测定 采用英国PP-Systems公司产CIRAS-1进行元宝枫净光合速率日变化测定, 7:00—17:00每隔2 h测定一次。
1.3.3 光合响应曲线测定 于晴天的9:00—11:00,用CIRAS-1型便携式光合测定仪进行光合响应曲线测定。采用LED红/蓝光源,设定光合有效辐射梯度为0(CK处理为等量的1/2改良Hoagland 营养液),50,100,150,200,250,300, 400,500, 600,800,1 000,1 200,1 300,1 400,1 600,1 800 μmol·(m-2·s-1),CO2浓度采用大气CO2浓度。
1.4 数据分析
数据采用Excel,DPS 7.05,SPSS 13.0进行分析处理。
2 结果与分析
2.1 Cd胁迫处理下元宝枫叶片叶绿素含量变化
从表1可以看出,不同质量浓度胁迫下,元宝枫叶绿素含量差异显著;随着胁迫浓度的增加,元宝枫叶绿素a、叶绿素b、叶绿素总量显著降低;与对照相比,50 mg·L-1 CdCl2·2.5 H2O处理下叶绿素a、叶绿素b、叶绿素总量分别降低了41%,30.4%,38.6%;100 mg·L-1 CdCl2·2.5 H2O处理下叶绿素a、叶绿素b、叶绿素总量分别降低了96%,38.8%,82.2%。可见,重金属Cd胁迫严重降低了叶绿素的合成,且胁迫浓度越高叶绿素含量降低越多,从而影响植物光合作用,阻碍植物的正常生长。
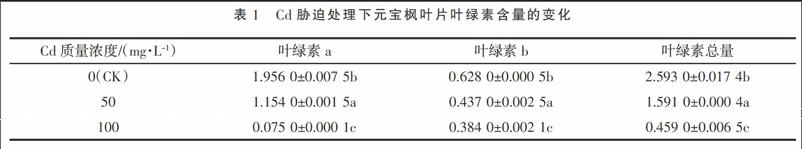
2.2 Cd胁迫处理下元宝枫叶片光合速率日变化
从图1可以看出,元宝枫叶片的Pn日变化呈单峰型,随着Cd胁迫浓度的增加,元宝枫叶片的净光合速率逐渐降低。正常情况下,元宝枫叶片净光合速率随光强的增加而升高。在Cd胁迫处理条件下,随着光强的增加,元宝枫的净光合速率下降较快,可见,高光强加快了植物的蒸腾作用,使得本身受重金属毒害的元宝枫失水加重,加上叶绿素合成降低,使得元宝枫叶片净光合速率下降更明显。而由于胁迫时间的加重,使得元宝枫叶片伤害程度加深,以至于净光合速率接近于0,可见,重金属Cd胁迫严重影响了元宝枫的正常生长。
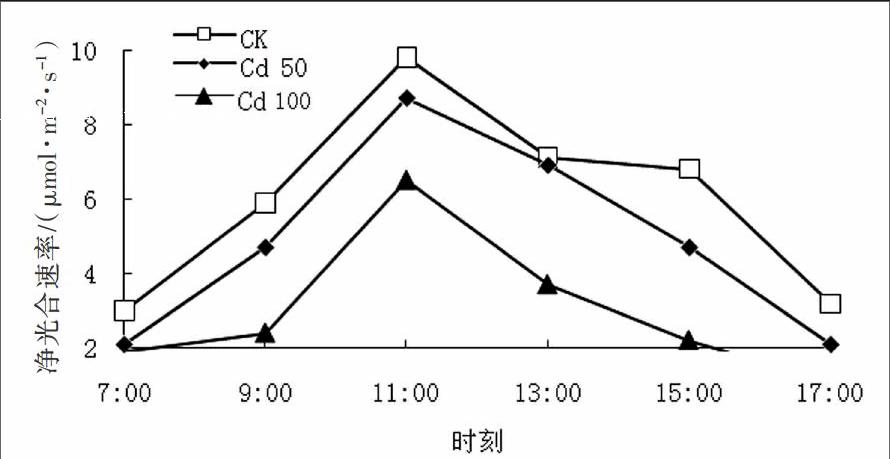
2.3 Cd胁迫处理下元宝枫叶片Pn—PAR响应曲线
由图2可知,元宝枫叶片的Pn—PAR响应曲线趋势相同,净光合速率均随光强的增加而升高;与对照相比,Cd胁迫处理下元宝枫净光合速率较低且下降更为明显;在轻度Cd胁迫下,元宝枫的Pn—PAR响应曲线与对照相比差别不大,通过拟合曲线方程可得,对照元宝枫叶片的LSP为1 321.77 μmol·(m-2·s-1),50 mg·L-1 Cd处理下元宝枫叶片的LSP为1 287.45 μmol·(m-2·s-1),可见,在元宝枫叶片接受相同光照条件下,植物自身的防御系统可以抵御一定逆境胁迫。而在重度胁迫,100 mg·L-1 Cd处理下元宝枫叶片的净光合速率显著降低,其叶片的LSP为1 021.64 μmol·(m-2·s-1),光饱和点降低,植物不能利用更多的光,容易产生光抑制,严重影响了植物的光合作用,阻碍了元宝枫的正常生长。
3 结论与讨论
重金属胁迫下植物体内产生大量的活性氧自由基,同时,重金属离子可以替代酶蛋白反应活性中心的金属离子,使酶蛋白变性失活,导致植物死亡[7]。研究发现,Cd2+可诱导产生活性氧,抑制一些抗氧化酶活性。在烟草中,Cd2+能促进膜脂过氧化,降低SOD,CAT,APX,GR等酶的活性[8]。孙涛等[9]研究认为,Cd2+进入细胞溶质后首先引起细胞抗氧化能力瞬时缺失,如引起AsA-GSH循环2个关键酶GR和APX的瞬时失活,启动H2O2的积累,进而诱导抗氧化系统的二次防御作用。本试验研究发现,重金属Cd胁迫降低了元寶枫叶片叶绿素含量,进而降低元宝枫叶片的净光合速率,严重阻碍了元宝枫的光合作用。
同时,研究发现,在重金属胁迫下,植物呼吸作用紊乱,不仅减少了供给植物正常生命活动的能量,而且还会使一部分能量转移到适应逆境变化中,从而抑制植物的生长发育[10-11]。刘登义等[12]研究发现,黄豆和小麦种子在Cu和As处理下呼吸强度降低,萌发需要的能量和产物不足,从而对种子萌发产生了明显的抑制作用。0.1 mmol·L-1 Cd处理小麦根部时,对照组和试验组的呼吸强度基本没有差异;当胁迫浓度增大到1 mmol·L-1时,小麦根部的呼吸作用明显被抑制。一般认为,低浓度的Cd刺激植物呼吸酶和三羧酸循环式呼吸增强,而高浓度的Cd使酶活性受到抑制,呼吸作用下降[13]。重金属污染是目前我国亟待解决的问题之一,而土壤植物修复技术是一种新兴的治理、缓解土壤重金属污染并能够回收土壤中重金属的技术手段。由于植物本身的差异及离子毒性效应的不同,使得对植物重金属污染的适应机制的研究也变得极为复杂[14-16]。本试验通过不同浓度的重金属处理,研究了其对元宝枫光合作用的影响,但其降低元宝枫叶片叶绿素含量和净光合速率的机制、通过何种途径置换了光合电子传递链中的哪些金属离子、抑制了哪些光合基因的表达以及如何提高元宝枫抵抗重金属的能力,还有待于进一步研究。
参考文献:
[1]臧德奎.园林树木学[M].北京:中国建筑工业出版社,2007:340.
[2]郭亚平,胡曰利.土壤-植物系统中重金属污染及植物修复技术[J].中南林学院学报,2005,25(2):59-62.
[3]王忠.植物生理学[M].北京:中国农业出版社,2000:121.
[4]徐向华.超积累植物商陆吸收累积锰机理研究[D].杭州:浙江大学,2006.
[5]郭智,原海燕,奥岩松.镉胁迫对龙葵幼苗光合特性和营养元素吸收的影响[J].生态环境学报,2009,18(3):824-829.
[6]李永杰.6种城市绿化树种苗木对土壤Cu、Pb污染的生理响应及耐性评价[D].北京:北京林业大学,2010.
[7]史贵涛,陈振楼,李海雯,等.城市土壤重金属污染研究现状与趋势[J].环境监测管理与技术,2006,18(6):9-12, 24.
[8]田生科.超积累东南景天(Sedum alfredii Hance)对重金属(Zn/Cd/Pb)的解毒机制[D].杭州:浙江大学,2010.
[9]孙涛, 张玉秀,柴团耀. 印度芥菜重金属耐性机理研究进展[J]. 中国生态农业学报, 2011(1):222-234.
[10]孔祥生,张妙霞,郭秀璞.Cd2+毒害对玉米幼苗细胞膜透性及保护酶活性的影响[J].农业环境科学学报,1999,18(3):133-134.
[11]张磊,于燕玲,张磊.外源镉胁迫对玉米幼苗光合特性的影响[J].华北农学报,2008,23(1):101-104.
[12]刘登义,王友保.Cu、As对作物种子萌发和幼苗生长影响的研究[J].应用生态学报,2002,13(2):179-182.
[13]郎明林, 张玉秀, 柴团耀. 植物耐重金属超富集机理研究进展[J]. 西北植物学报, 2003(11) : 2020-2029.
[14]李子芳,刘惠芬,熊肖霞,等.镉胁迫对小麦种子萌发幼苗生长及生理生化特性的影响[J].农业环境科学学报,2005,24(S1):17-20.
[15]何俊瑜,任艳芳.镉胁迫对水稻种子萌发、幼苗生长和淀粉酶活性的影响[J].华北农学报,2008,23(S1):131-134.
[16]白瑞琴,晁公平,孙华,等.重金属镉胁迫对蜀葵、二月蓝种子萌发和幼苗生长的毒害效应研究[J].华北农学报,2009,24(2):134-138.
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
中国生物防治学报(2021年1期)2021-04-18 03:26:07
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
山东林业科技(2019年2期)2019-06-03 10:10:56
吉林农业(2019年9期)2019-01-06 06:06:24
中国果业信息(2019年1期)2019-01-05 17:41:42
生物学教学(2017年9期)2017-08-20 13:22:32
Coco薇(2016年5期)2016-06-03 09:17:41
农村.农业.农民(2016年24期)2016-04-02 05:00:25
考试周刊(2016年6期)2016-03-11 08:14:32
