
超表达牛奶子EutPDS提高番茄果实番茄红素含量*
2017-03-08 07:12程珍霞胡海涛王长春郭卫东
林业科学 2017年1期
程珍霞 胡海涛 杨 莉 王长春 郭卫东 杨 玲
(浙江师范大学化学与生命科学学院 金华 321004)
超表达牛奶子EutPDS提高番茄果实番茄红素含量*
程珍霞 胡海涛 杨 莉 王长春 郭卫东 杨 玲
(浙江师范大学化学与生命科学学院 金华 321004)
【目的】 牛奶子果实中富含番茄红素,八氢番茄红素脱氢酶(PDS)是番茄红素生物合成上游的重要酶。特异性超表达牛奶子EutPDS基因(GenBank登录号:GQ254067),以期提高番茄果实中的番茄红素含量。【方法】 通过RT-PCR方法从牛奶子果实中分离EutPDS基因cDNA及基因组全长,Southern杂交分析基因的拷贝数,专用软件分析此基因及蛋白结构。采用番茄2A11启动子构建果实特异性超表达载体,转化根瘤农杆菌GV3101后,借助农杆菌介导的叶盘法转化至‘中蔬四号’番茄。半定量RT-PCR检测转基因植株果实EutPDS基因的表达,高效液相色谱仪、实时定量PCR分别用于分析转基因番茄果实主要类胡萝卜素组分含量和内源类胡萝卜素代谢相关基因表达的变化。【结果】 从牛奶子中克隆的EutPDS基因长度为1 920 bp,含有1 749 bp ORF。其基因组序列无内含子,单拷贝。EutPDS编码1个582个氨基酸的蛋白,该蛋白与其他植物PDS有很高的同源性,具有典型的二核苷酸结合域、类胡萝卜素结合域。采用农杆菌介导方法转化番茄叶盘,PCR检测,潮霉素筛选,成功获得了2个EutPDS转基因番茄株系,其果实颜色较对照更红。半定量分析显示外源EutPDS基因在转基因番茄果实中超表达,实时定量PCR分析表明番茄红素生物合成上游2个内源基因SlPSY和SlZDS受影响显著上调,同时1个下游基因SlLCY-e的表达则显著下调,使得转基因番茄果实中番茄红素含量为野生型的2倍,但β-胡萝卜素含量无显著变化。【结论】 在番茄果实中特异性超表达牛奶子EutPDS基因可有效促进番茄果实中番茄红素的合成和积累。
牛奶子; 番茄; 番茄红素; 八氢番茄红素脱氢酶基因(PDS); 载体构建; 遗传转化
番茄红素是一种具有11个共轭双键的类胡萝卜素化合物,其抗氧化能力极强,通过清除游离的氧自由基延缓细胞的衰老,对人体健康有着至关重要的作用(Dimascioetal., 1989; Mülleretal., 2016)。但人体自身无法合成番茄红素,必须通过饮食从食物中摄取,其85%来源于番茄果实。随着人们对番茄红素的需求日益上升,有必要通过生物技术手段增加番茄中番茄红素含量,提高其营养品质。
植物类胡萝卜素生物合成途径已基本阐明,始于八氢番茄红素合成酶(PSY)催化2分子牻牛儿牻牛儿基焦磷酸向八氢番茄红素的转化,八氢番茄红素经过4步脱氢形成番茄红素,中间物番茄红素分2条支路在环化酶、羟化酶和加氧酶的作用下生成种类多样的类胡萝卜素(Direttoetal., 2007; Guoetal., 2009)。其中八氢番茄红素脱氢酶(phytoene desaturase, PDS)催化八氢番茄红素经过连续的2步脱氢反应转变为六氢番茄红素(邹礼平等, 2012; Liuetal., 2010)。许多植物的PDS基因已被克隆,但研究多限于序列比对及组织表达分析。近年来,应用类胡萝卜素合成途径中相关基因改良作物品质性状的研究取得了较大进展,积累β-胡萝卜素的“黄金稻米”(Beyeretal., 2002; Paineetal., 2005)和“黄金马铃薯”(Direttoetal., 2007)就是最著名的例子。理论上,超表达PDS基因促进八氢番茄红素向下游代谢反应可以提高番茄红素的含量,但由于对番茄红素生物合成的表达调控机制尚不完全清楚,在应用遗传工程技术提高植物靶器官番茄红素含量时,存在较大的盲目性(Dattaetal., 2014)。在小麦(Triticumaestivum)胚乳中特异性表达玉米(Zeamays)基因y1(编码PSY)和细菌crtI(编码PDS),转基因小麦的总类胡萝卜素含量上升了10.8倍(Congetal., 2009); 在小麦中同时表达细菌CrtB(编码PSY)和CrtI基因,其种子干质量的总类胡萝卜素含量提高了近8倍,β-胡萝卜素含量提高了65倍,维生素A前体含量提高了76倍(Wangetal., 2014)。同样,在水稻(Oryzasativa)种子中超表达PSY和crtI基因,转基因稻米中β-胡萝卜素而非番茄红素大幅增加(Dattaetal., 2014)。超表达PSY的转基因黄金稻米之所以富含β-胡萝卜素而非番茄红素,是因为胚乳中PDS基因的表达水平太低(Schaubetal., 2005)。在番茄(Solanumlycopersicum)中超表达SlPDS基因,获得的转基因株系果实中的番茄红素含量提高了1.4倍(邹礼平等, 2012)。这些研究结果说明,PDS基因可能参与调控番茄红素生物合成与积累。
牛奶子(Elaeagnusumbellata)为胡颓子科(Elaeagnaceae)胡颓子属植物,起源于我国,其成熟红色果实番茄红素含量高达0.54 mg·g-1,为番茄的5~18倍(Guoetal., 2009),很可能成为工厂化生产番茄红素的原料。前期的研究结果显示,牛奶子EutPDS基因表达在果实成熟早期的黄果期就大幅上调,可能与其番茄红素含量高有关(Guoetal., 2009)。本文在分析EutPDS基因结构基础上,构建果实特异性超表达载体,利用番茄遗传转化体系使其在番茄果实中稳定表达,显著提高了番茄红素含量,证实了EutPDS基因在番茄红素生物合成及积累中的重要功能,为今后的利用提供了理论依据。
1 材料与方法
1.1 试验材料
牛奶子定植于浙江师范大学生物园内,以成熟红色果实为试验材料(Guoetal., 2009)。番茄‘中蔬四号’种子由浙江省农业科学院提供,种植在盆中,正常管理。采摘破色期后10天的番茄果实作为成熟果,去除种子后将果肉切成小块,液氮速冻后-80 ℃保存,种子晒干后保存。TRIzol试剂、限制性内切酶、KOD高保真酶、Mix酶、T4 DNA连接酶、潮霉素等购自北京科百奥生物技术有限公司; 引物由Invitrogen上海生物技术公司合成; M-MLV逆转录酶、载体PMD 18-T购于大连宝生物生物技术有限公司。色谱标准品β-胡萝卜素购自美国Sigma公司,番茄红素购自北京绿百草科技发展有限公司。
1.2 目的基因EutPDS的克隆与分析
采用热硼酸盐方法提取牛奶子成熟果实中的总RNA,以oligo(dT)18为反转录引物,M-MLV逆转录酶合成一链cDNA(Guoetal., 2009),采用CTAB法提取牛奶子叶片DNA。跨EutPDS基因(GenBank登录号:GQ254067)ORF区设计特异引物(上游5′-ATGTCCCAGTGGGGGTGTACTAC-3′和下游5′-TGAACCTGTCCCTCTTTTGCT-3′),分别以果实一链cDNA和叶片DNA扩增出其cDNA及基因组全长序列。25 μL反应体系: 上下游引物(10 μmol·L-1)各1 μL,模板1 μL,10 × LaPCR缓冲液2.5 μL,dNTPs(2.5 mmol·L-1)1.6 μL,LaTaq DNA聚合酶(Takara)0.2 μL。PCR反应条件: 94 ℃ 3 min; 94 ℃ 15 s,60 ℃ 30 s,72 ℃ 6 min,35个循环; 72 ℃ 10 min。
分别用BamHⅠ,SacⅠ,EcoRⅠ和HindⅢ 4种内切酶消化叶片DNA。100 μL酶切体系包括10 × buffer 10 μL、内切酶4 μL、DNA 10 μL(15~20 μg)。混匀后37 ℃保温15 h。探针制备用上游引物5′-ATGTCCCAGTGGGGGTGTACTAC-3′和下游引物5′-CGGTTGAACTGTCCTGGTTTGT-3′。使用罗氏(Roche)公司Southern杂交试剂盒,按说明书操作。1.3 果实特异性EutPDS超表达载体的构建
根据EutPDS序列设计上游引物F(KpnⅠ)(5′-GGGTACCATGTCCCAGTGGGGGTGTACT-3′)和下游引物R(BamHⅠ)(5′-CGGATCCTCAAAAAGA GCCTGTCTCTG-3′),以牛奶子红果一链cDNA为模板,用高保真酶进行RT-PCR扩增。PCR反应条件: 94 ℃ 3 min; 94 ℃ 30 s,58.8 ℃ 30 s,68 ℃ 2 min,35个循环; 68 ℃ 10 min。产物经1%琼脂糖凝胶电泳检测,回收目的片段后连接至PMD 18-T载体上,并转化至大肠杆菌(Escherichiacoli)感受态DH5α,挑选阳性克隆送Invitrogen公司测序验证。在本实验室已经构建EutPSY基因的番茄果实特异性启动子2A11植物表达载体pCAMBIA 2A11-EutPSY-1301基础上,提取pCAMBIA 2A11-EutPSY-1301和EutPDS-PMD 18-T质粒,使用相同的内切酶KpnⅠ、BamHⅠ分别进行双酶切,回收得到EutPDS基因和2A11-1301载体片段,用T4 DNA连接酶连接,完成EutPDS基因替换EutPSY基因。转化大肠杆菌,进行双酶切检测,正确的重组质粒命名为pCAMBIA 2A11-EutPDS-1301。
1.4 番茄的遗传转化
将重组质粒pCAMBIA 2A11-EutPDS-1301转入根瘤农杆菌(Agrobacteriumtumefaciens)GV3101,菌液PCR筛选及双酶切验证。利用叶盘法转化番茄(邹礼平等, 2012),使用20 mg·L-1潮霉素筛选。以番茄再生植株叶片基因组DNA为模板,根据载体上的GUS标记基因序列合成引物(上游5′-GTGAATCCGCACCTCT-3′和下游5′-ATCGCCGCT TTGGACATA-3′),进行PCR检测。
1.5 半定量RT-PCR分析EutPDS基因的表达
取3个成熟番茄果实,用TRIzol法提取总RNA,逆转录合成一链cDNA为模板,根据EutPDS的5′非保守域设计特异引物F(5′-GCTACGATGA AAGAACTGGCTAA-3′)和R(5′-AGGGACGGCAAG GTTCACAA-3′)(Guoetal., 2009),进行PCR扩增。采用番茄泛素基因(SlUBI)为内参基因(F: 5′-AAGCAATGGATGCTGAGGCT-3′; R: 5′-GAAGGTG CCGTTGAATGACA-3′)(Tikunovetal., 2013)。PCR反应程序: 95 ℃ 3 min; 95 ℃ 30 s,60 ℃ 30 s,72 ℃ 15 s,30个循环; 72 ℃ 10 min。
1.6 番茄果实主要类胡萝卜素组分测定
用3个成熟果实进行类胡萝卜素的提取、浓缩、样品的制备及高效液相色谱分析,均参照胡海涛等(2014)的方法,在450 nm下检测,重复3次。运用Excel 2007软件对数据进行统计分析,P<0.05为差异显著。
1.7 实时定量PCR分析番茄内源类胡萝卜素代谢相关基因表达
番茄SlPSY、SlPDS、ζ-胡萝卜素脱氢酶(SlZDS)、番茄红素β-环化酶(SlLCY-b)、番茄红素ε-环化酶(SlLCY-e)等基因的引物序列参考文献(Tikunovetal., 2013),用SYBR® Premix Ex TaqTMⅡ(TaKaRa公司)在StepOnePlusTM Real-Time PCR System(ABI公司)分析成熟番茄果实中类胡萝卜素合成基因的表达,SlUBI为内参基因,计算基因的相对表达量(Guoetal., 2009)。
2 结果与分析
2.1 牛奶子EutPDS基因的结构
分别以牛奶子叶片DNA、果实cDNA为模板,扩增出大小一致的基因,全长约1 920 bp(图1); 测序结果也表明EutPDS基因的DNA序列与其cDNA序列一致,说明其没有内含子。EutPDS基因的ORF为1 749 bp,编码582个氨基酸,计算出的分子质量为65.2 kDa,等电点为8.12。Southern杂交结果表明,EutPDS基因在牛奶子基因组中只有1个拷贝,番茄等大多数植物中的PDS为单拷贝(Mannetal., 1994)。序列比对分析发现,EutPDS氨基酸序列与花椰菜(Brassicaoleraceavar.botrytis)(ABV46593)、杏树(Prunusarmeniaca)(AAX33347)和拟南芥(Arabidopsisthaliana)(AAK64084)的同源性最高,与番茄(ABR57230)同源性也达80%。EutPDS蛋白含有典型的与FAD或者NAD(P)结合的βαβ吡啶二核苷酸结合域、类胡萝卜素结合域(图2)。
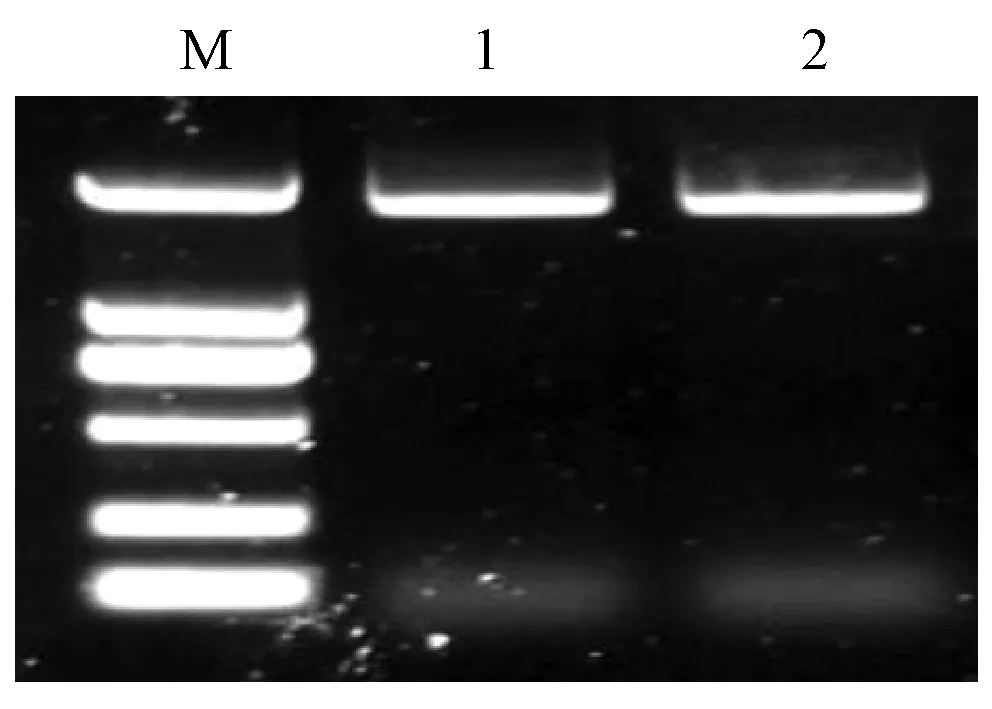
图1 牛奶子EutPDS基因全长扩增产物Fig.1 Agarose gel electrophoresis of PCR products of full-length EutPDS from Elaeagnus umbellataM. DL2000 marker. 1. DNA全长; 2. cDNA全长。1.Full-length DNA; 2. Full-length cDNA.
2.2 果实特异性超表达载体的构建
以牛奶子成熟果实一链cDNA为模板,合成带有酶切位点的引物,用高保真酶进行PCR扩增,得到1条约2 kb的清晰条带,回收测序。连接转化后,获得EutPDS-PMD 18-T质粒。将扩增产物定向连接至pCAMBIA301-2A11载体(图3)。经酶切验证的重组质粒pCAMBIA 2A11-EutPDS-1301转化至根瘤农杆菌GV3101中,菌液PCR及双酶切结果显示重组载体已成功转化到农杆菌中。2.3 番茄的遗传转化及PCR检测
将果实特异性超表达载体PCAMBIA 2A11-EutPDS-1301通过农杆菌介导法转化至番茄‘中蔬四号’,获得转基因再生植株(图4)。特异扩增转基因再生苗的GUS能够鉴定表达载体的T-DNA区是否插入到番茄基因组中,检测结果见图5。对番茄转化株系T1代种子进行潮霉素筛选,得到2个结实的转基因阳性株系。
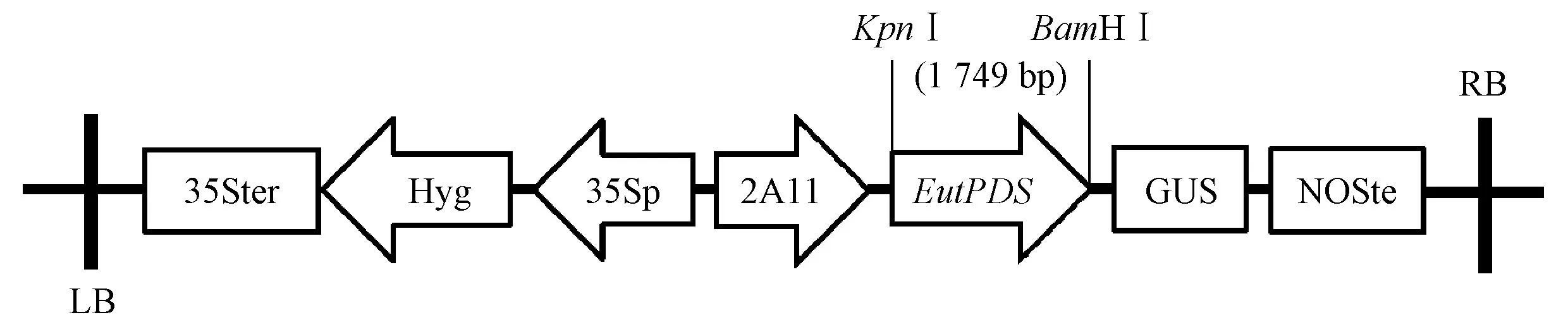
图3 牛奶子EutPDS基因超表达载体的构建Fig.3 Construction of EutPDS-overexpression vector

图4 转基因番茄植株的再生及结实Fig.4 Regeneration and fruit set of transgenic tomato plantsA. 无菌苗; B. 不定芽的分化; C. 不定芽的生长; D. 生根; E. 再生植株的移植; F. 转基因番茄果实。 A. Aseptic plantlet; B. Differentiation of adventitious buds; C. Growth of adventitious buds; D. Rooting; E. Transplantation of regenerated plants; F. Transgenic tomato fruit.
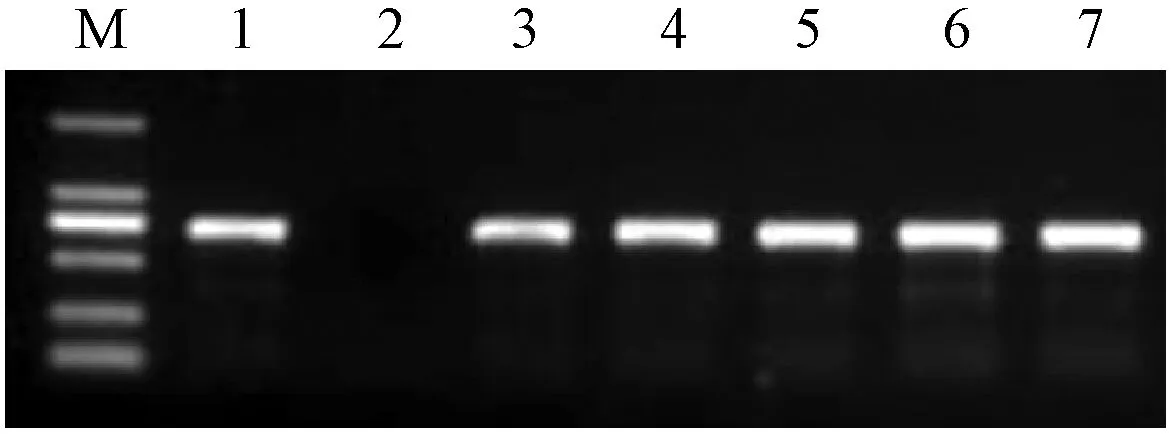
图5 转基因番茄再生植株的PCR检测Fig.5 PCR identification of transgenic tomatoesM. DL2000 marker. 1. 阳性对照; 2. 野生型; 3-7. 转基因番茄。1. Positive control; 2. Wild type; 3-7. Transgenic tomato.
2.4 转基因番茄成熟果实中EutPDS的表达
利用半定量RT-PCR分析外源基因EutPDS在番茄中的表达,发现2个转基因株系中出现了预期的EutPDS基因条带,而野生型中没有检测到(图6),说明转基因番茄中外源EutPDS基因已在番茄果实中组成型表达。
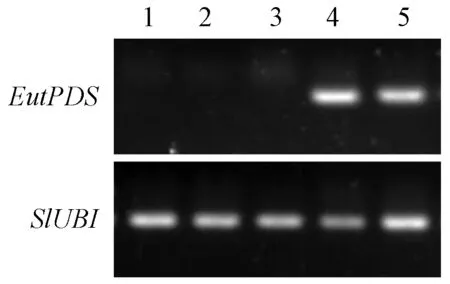
图6 EutPDS基因在转基因及野生型番茄中的半定量分析Fig.6 Semi-quantitative PCR analysis of EutPDS expression in the tomato fruit of transgenic line and wild type1-3. 野生型; 4,5. 转基因株系OE-1,OE-2。 1-3. Wild type; 4,5. Transgenic line OE-1 and OE-2, respectively.
2.5 转基因番茄成熟果实的类胡萝卜素组分
野生型及转基因番茄果实的主要类胡萝卜素组分为β-胡萝卜素和番茄红素。与野生型比较,2个转基因株系番茄果实中的β-胡萝卜素含量没有显著变化,但番茄红素含量显著增高(P<0.05),从而使总胡萝卜素含量也显著上升; 且以EutPDS表达量相对高的株系OE-1中的番茄红素和总胡萝卜素含量增幅较大,分别为(133.81±14.49)、(161.58±18.24)μg·g-1(以鲜质量计),约为野生型的2倍(图7)。说明外源基因EutPDS的超表达提高了番茄红素的含量。
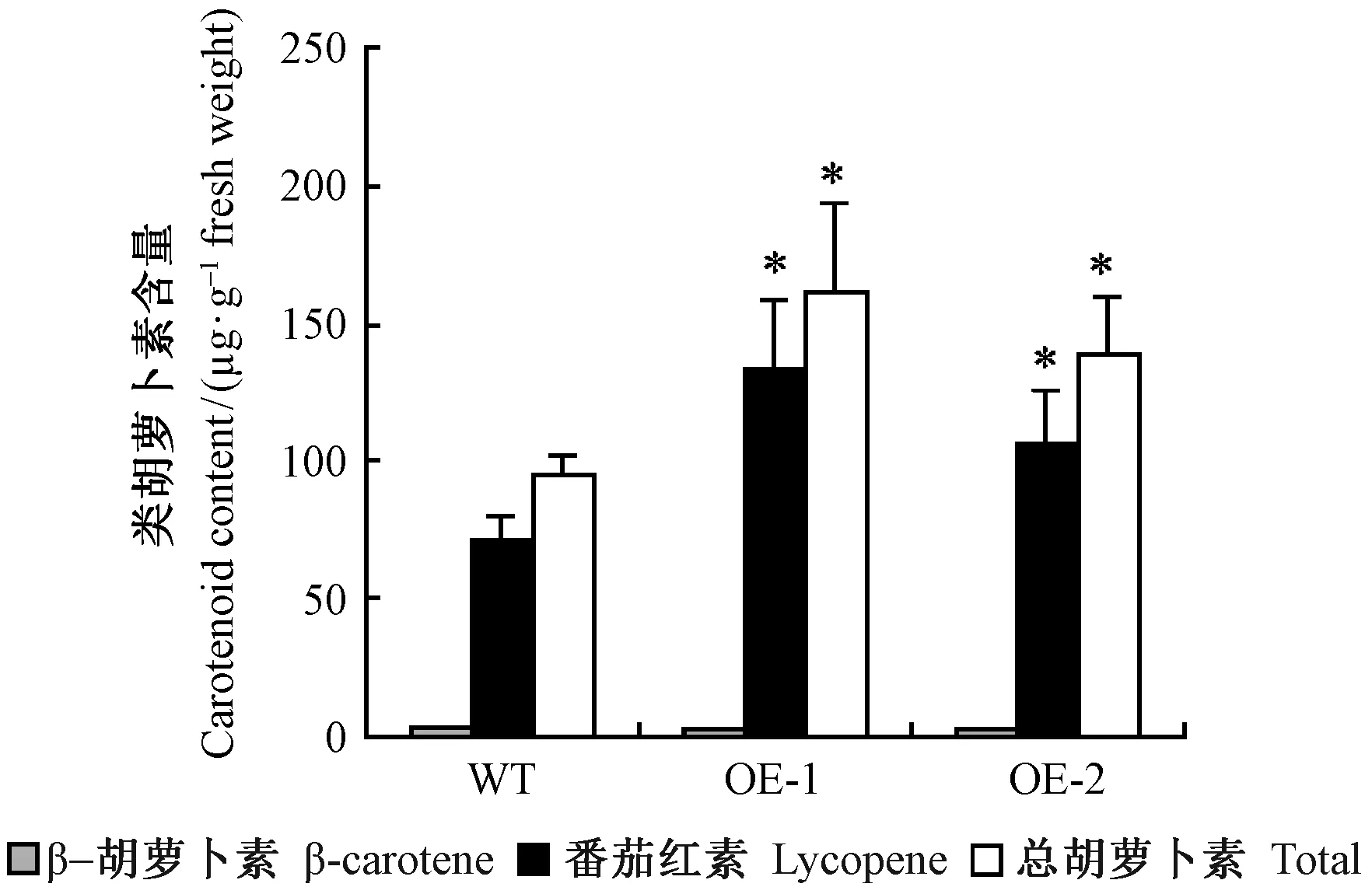
图7 转基因番茄成熟果实中类胡萝卜素的含量Fig.7 Carotenoids content of ripe transgenic tomato fruitsWT: 野生型; OE-1, OE-2: 转基因株系。*:表示P<0.05差异显著。WT: Wild type; OE-1, OE-2: Transgenic lines. *:Significance at 0.05 level.
2.6EutPDS超表达的番茄果实中类胡萝卜素合成基因的表达
番茄果实中类胡萝卜素的生物合成始于SlPSY催化2分子牻牛儿牻牛儿基焦磷酸向八氢番茄红素的转化,八氢番茄红素经过SlPDS和六氢番茄红素脱氢酶(SlZDS)的连续脱氢生成番茄红素,中间物番茄红素在2种番茄红素环化酶(SlLCY-b、SlLCY-e)的作用下继续代谢形成β-胡萝卜素、α-胡萝卜素等,这些胡萝卜素经过羟化酶和加氧酶的作用形成种类多样的类胡萝卜素及其他下游代谢产物(邹礼平等, 2012; Liuetal., 2010)。选取超表达EutPDS显著的转基因番茄株系OE-1,利用实时定量PCR分析表明,EutPDS基因超表达对转基因植株果实中内源类胡萝卜素合成相关基因表达的影响不同: 与野生型比较,除SlPDS基因表达没有显著变化外,番茄红素生物合成上游基因SlPSY和SlZDS均极显著上调(P<0.01); 下游基因SlLCY-b的表达没有显著变化,SlLCY-e的表达则极显著下降(图8)。
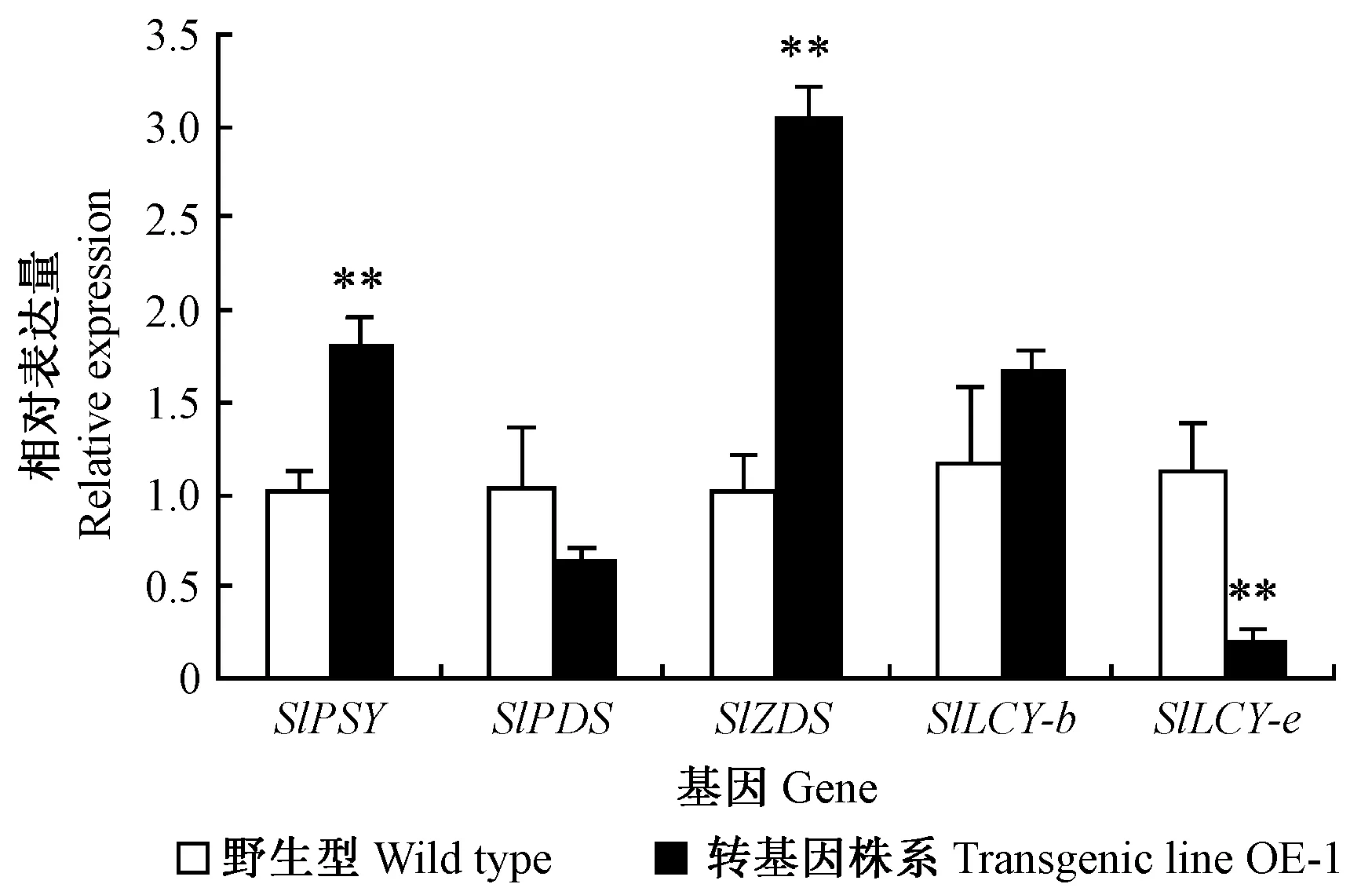
图8 实时定量PCR分析转基因番茄果实内源类胡萝卜素生物合成基因的表达Fig.8 Real-time quantitative PCR analysis of the expression of endogenous carotenoid biosynthetic genes in transgenic tomato fruit**表示P<0.01差异显著Significance at 0.01 level.
3 讨论
朱海生等(2011)克隆了草莓(Fragaria×ananassa)PDS基因,序列比对发现其编码的氨基酸序列与其他物种具有很高的相似性,该基因表达水平在红色果实中最高,在叶片中最低。陈段芬等(2008)克隆了中国水仙(Narcissustazettavar.chinensis)PDS基因,其在花中表达最高。EutPDS基因主要在叶、花和成熟果实中表达(Guoetal., 2009)。EutPDS基因没有内含子,与番茄(X71023)、拟南芥(ATCHRIV38)和水稻(AC079633)基因组中PDS基因的多个外显子/内含子结构(≥10)(Mannetal., 1994; Matthewsetal., 2003)不同。EutPDS在基因组中为单拷贝,番茄、玉米和水稻基因组中的PDS也只有1个拷贝(Aracrietal., 1994; Matthewsetal., 2003),温州蜜柑(Citrusunshiu)、大豆(Glycinemax)有2~3个拷贝(Bartleyetal., 1991; Kitaetal., 2001),PDS基因的低拷贝数特征可能是植物的共性(Kitaetal., 2001)。
番茄红素的抗氧化能力较β-胡萝卜素强(Dimascioetal., 1989; Mülleretal., 2016),食品营养价值更高,因此人们试图利用基因工程的方法提高可食用植物靶器官中番茄红素的含量。果实特异启动子驱动下,在‘Ailsa Craig’遗传背景下超表达噬夏孢欧文菌(Erwiniauredovora)PSY基因,转基因番茄果实中的番茄红素、β-胡萝卜素、总类胡萝卜素含量分别提高了1.8~2.1,1.6~2.7和2~4倍(Fraseretal., 2002)。使用胚乳特异性启动子在小麦中共表达来源于噬夏孢欧文菌的PSY和PDS基因,其种子(干质量)的总类胡萝卜素、β-胡萝卜素含量分别提高到8倍和65倍,但番茄红素的含量仅略增加(Wangetal., 2014)。Schaub等(2005)认为,超表达水仙花(Narcissuspseudonarcissus)PSY的转基因黄金稻米之所以富含β-胡萝卜素而非番茄红素,是因为胚乳中PDS基因的表达水平太低。在番茄‘中蔬5号’中超表达内源SlPDS基因,获得的转基因株系果实中的番茄红素含量平均提高了1.4倍(邹礼平等, 2012); 但将噬夏孢欧文菌的PDS组成型表达载体转入番茄‘Ailsa Craig’,果实中β-胡萝卜素提高了3倍,而番茄红素含量减半,类胡萝卜素的总量也只有野生型的50%(Römeretal., 2000)。由此可见,番茄红素生物合成的调控机制极其复杂。植物类胡萝卜素生物合成受一系列酶或基因调控,某一种类胡萝卜素合成酶催化活性的改变或代谢中间物含量的变化会调节其他类胡萝卜素合成内源基因的表达(Beyeretal., 2002; Paineetal., 2005; Direttoetal., 2007)。此外,使用的基因及其来源、启动子、受体遗传背景等的不同均会影响转基因植株靶器官中类胡萝卜素的组分及其含量(Paineetal., 2005; Direttoetal., 2007; Wangetal., 2014)。
4 结论
牛奶子果实富含番茄红素,EutPDS表达水平在果实成熟早期就大幅上调。本文在番茄果实中特异性超表达外源EutPDS基因,不但克服了SlPDS酶的限制,而且显著诱导了番茄红素生物合成上游内源基因SlPSY和SlZDS的表达、显著下调下游内源基因SlLCY-e表达,增加了番茄红素生物合成上游原料的供给,降低分支途径的代谢消耗,从而显著促进番茄红素的合成与积累,番茄红素含量为野生型对照组的2倍,β-胡萝卜素的含量则无显著变化。转基因番茄株系果实中的番茄红素积累显著提高,不但证实了EutPDS在番茄红素合成中重要的定向调节作用,而且为EutPDS作为番茄红素生物反应器的候选功能基因提供了理论依据。
陈段芬, 彭镇华, 高志民. 2008. 中国水仙八氢番茄红素脱氢酶基因(PDS)的克隆及表达分析. 分子植物育种, 6(3): 574-578.
(Chen D F, Peng Z H, Gao Z M. 2008. Cloning and expression analysis ofPDSgene inNarcissustazettavar.chinensis. Molecular Plant Breeding, 6(3): 574-578. [in Chinese])
胡海涛, 金晓琴, 成龙平, 等. 2014. 密花胡颓子果实主要类胡萝卜素组分及含量分析. 中国农业科学, 47(8): 1652-1656.
(Hu H T, Jin X Q, Cheng L P,etal. 2014. Determination of carotenoids in the fruit ofElaeagnusconfertaRoxb. by high performance liquid chromatography. Scientia Agricultura Sinica, 47(8): 1652-1656. [in Chinese])
朱海生, 李永平, 温庆放, 等. 2011. 草莓八氢番茄红素脱氢酶基因PDS的克隆及特征分析. 园艺学报, 38(1): 55-60.
(Zhu H S, Li Y P, Wen Q F,etal. 2011. Cloning and characterization ofPDSgene inFragaria×ananassa. Acta Horticulturae Sinica, 38(1): 55-60. [in Chinese])
邹礼平, 高和平, 钟亚琴. 2012. 八氢番茄红素脱氢酶基因超表达载体的构建及表达鉴定. 湖北农业科学, 51(2): 393-395.
(Zou L P, Gao H P, Zhong Y Q. 2012. Construction of overexpression vector for phytoene dehydrogenase gene and its expression identification in tomato. Hubei Agricultural Sciences, 51(2): 393-395. [in Chinese])
Aracri B, Bartley G E, Scolnik P A,etal. 1994. Sequence of the phytoene desaturase locus of tomato. Plant Physiology, 106(2): 789.
Bartley G E, Viitanen P V, Pecker I,etal. 1991. Molecular cloning and expression in photosynthetic bacteria of a soybean cDNA coding for phytoene desaturase, an enzyme of the carotenoid biosynthesis pathway. Proceedings of the National Academy of Sciences USA, 88: 6532-6536.
Beyer P, Al-Babili S, Ye X,etal. 2002. Golden rice: introducing the beta-carotene biosynthesis pathway into rice endosperm by genetic engineering to defeat vitamin A deficiency. The Journal of Nutrition, 132(3): 506S-510S.
Cong L, Wang C, Chen L,etal. 2009. Expression ofphytoenesynthase1 and carotene desaturasecrtIgenes result in an increase in the total carotenoids content in transgenic elite wheat (TriticumaestivumL.). Journal of Agricultural & Food Chemistry, 57(18): 8652-8660.
Datta K, Sahoo G, Krishnan S,etal. 2014. Genetic stability developed forβ-carotene synthesis in BR29 rice line using dihaploid homozygosity. PLoS One, 9: e100212.
Dimascio P, Kaiser S, Sies H. 1989. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Archives of Biochemistry and Biophysics, 274(2): 532-538.
Diretto G, Al-Babili S, Tavazza R,etal. 2007. Metabolic engineering of potato carotenoid content through tuber-specific overexpression of a bacterial mini-pathway. PLoS One, 2: e350.
Fraser P D, Romer S, Shipton C A,etal. 2002. Evaluation of transgenic tomato plants expressing an additional phytoene synthase in a fruit-specific manner. Proceedings of the National Academy of Sciences USA, 99(2): 1092-1097.
Guo X, Yang L, Hu H,etal. 2009. Cloning and expression analysis of carotenogenic genes during ripening of autumn olive fruit (Elaeagnusumbellata). Journal of Agricultural and Food Chemistry, 57(12): 5334-5339.
Kita M, Komatsu A, Omura M,etal. 2001. Cloning and expression ofCitPDS1, a gene encoding phytoene desaturase in citrus. Bioscience Biotechnology and Biochemistry, 65(6): 1424-1428.
Liu S, Sun L, Yang L,etal. 2010. Construction of phytoene desaturase (Pds) gene expression vector with tomato fruit specific E8 promoter. Food Science, 31(13): 196-199.
Mann V, Pecker I, Hirschberg J. 1994. Cloning and characterization of the gene for phytoene desaturase (Pds) from tomato (Lycopersiconesculentum). Plant Molecular Biology, 24(3): 429-434.
Matthews P D, Luo R, Wurtzel E T. 2003. Maize phytoene desaturase and zeta-carotene desaturase catalyse a poly-Z desaturation pathway: implications for genetic engineering of carotenoid content among cereal crops. Journal of Experimental Botany, 54(391): 2215-2230.
Müller L, Caris-Veyrat C, Lowe G,etal. 2016. Lycopene and its antioxidant role in the prevention of cardiovascular diseases—A critical review. Critical Reviews in Food Science and Nutrition, 56(11): 1868-1879.
Paine J A, Shipton C A, Chaggar S,etal. 2005. Improving the nutritional value of golden rice through increased pro-vitamin A content. Nature Biotechnology, 23(4): 482-487.
Römer S, Fraser P D, Kiano J W,etal. 2000. Elevation of the provitamin A content of transgenic tomato plants. Nature Biotechnology, 18(6): 666-669.
Schaub P, Al-Babili S, Drake R,etal. 2005. Why is golden rice golden (yellow) instead of red?. Plant Physiology, 138(1): 441-450.
Tikunov Y M, Molthoff J, de Vos R C H,etal. 2013. NON-SMOKY GLYCOSYLTRANSFERASE1 prevents the release of smoky aroma from tomato fruit. The Plant Cell, 25(8): 3067-3078.
Wang C, Zeng J, Li Y,etal. 2014. Enrichment of provitamin A content in wheat (TriticumaestivumL.) by introduction of the bacterial carotenoid biosynthetic genesCrtBandCrtI. Journal of Experimental Botany, 65(9): 2545-2556.
(责任编辑 徐 红)
Overexpression ofEutPDSGene fromElaeagnusumbellataIncreases Lycopene Content in Tomato Fruit
Cheng Zhenxia Hu Haitao Yang Li Wang Changchun Guo Weidong Yang Ling
(CollegeofChemistryandLifeSciences,ZhejiangNormalUniversityJinhua321004)
【Objective】 Lycopene has a strong anti-oxidative activity, which could remove free radicals of oxygen to delay the aging of cells of human body. Phytoene desaturase (PDS) is an important upstream enzyme of lycopene biosynthesis. The fruit ofElaeagnusumbellatawas found to accumulate high level of lycopene. This study was aimed to enhance lycopene component of tomato(Solanumlycopersicum)by overexpressingEutPDS(GenBank number: GQ254067) driven by fruit-specific promoter. 【Method】 The full-length cDNA and genomic DNA ofEutPDSgene were isolated fromE.umbellatafruit using RT-PCR. The copy number was determined by southern blot hybridization. The structural features ofEutPDSgene and protein were analyzed by using special software. A recombinant plasmid carryingEutPDSgene driven by fruit-specific promoter 2A11 was constructed. The expression vector was transferred into theAgrobacteriumtumefaciensstrain GV3101, and then introduced into a tomato variety ‘Zhongshu 4’ byAgrobacterium-mediated leaf disc method. Semi-quantitative RT-PCR was used to analyzeEutPDSexpression. High performance liquid chromatography and real-time RT-PCR were used to identity the main components of carotenoids and endogenous carotenoid gene expression in tomato fruits of both transgenic lines (OE) and wild-type, respectively. 【Result】 The clonedEutPDScDNA has 1 920 bp in length and contained a 1 749 bp ORF. TheEutPDSgene was not interrupted by any intron and exists as a single copy inE.umbellatagenome.EutPDSencodes a protein of 582 amino acids, which showed a high identity with other plant PDSs. Both a potential dinucleotide-binding motif and the carotenoid-binding domain were identified. The overexpressing vector ofEutPDSwas transformed into tomato leaves byAgrobacterium-mediated method. Two transgenic lines ofEutPDSwere obtained after PCR detection and hygromycin screen. The transgenic fruits were redder than wild type. Semi-quantitative PCR analysis showed thatEutPDSgene was overexpressed in transgenic tomato fruit. Real-time quantitative PCR analysis demonstrated that two endogenous upstream genes of lycopene biosynthesis,SlPSYandSlZDS, were significantly upregulated while a downstream geneSlLCY-ewas significantly downregulated in the fruit of OE-1 compared with the wild-type. Lycopene content in transgenic tomato fruits was nearly twice as much as that in the wild-type, butβ-carotene content was not significantly different.【Conclusion】 Fruit-specific overexpression ofEutPDSgene in tomato can significantly increase the biosynthesis and accumulation of lycopene.
Elaeagnusumbellata;Solanumlycopersicum; lycopene; phytoene desaturase gene (PDS); vector construction; genetic transformation
10.11707/j.1001-7488.20170108
2016-01-18;
2016-07-07。
国家自然科学基金面上项目(31071775); 浙江省科技计划项目(2008C24G2030004)。
S718.46
A
1001-7488(2017)01-0062-08
*杨玲为通讯作者。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
食品安全导刊(2021年20期)2021-08-30
四川农业科技(2019年5期)2019-07-01
新农村(浙江)(2019年5期)2019-05-29
新疆农垦科技(2016年10期)2016-06-15
浙江柑橘(2016年1期)2016-03-11
中国酿造(2016年12期)2016-03-01
中国果菜(2016年9期)2016-03-01
医学研究杂志(2015年8期)2015-06-22
郑州大学学报(理学版)(2014年2期)2014-03-01
