
芥蓝耐热性鉴定及耐热转录因子MBF1c表达分析
2017-02-27 03:06王浩伟陈国菊陈长明曹必好邹丽芳雷建军
中国蔬菜 2017年2期
王浩伟 陈国菊 陈长明 曹必好 邹丽芳 雷建军
(华南农业大学园艺学院,广东广州 510642)
芥蓝耐热性鉴定及耐热转录因子MBF1c表达分析
王浩伟 陈国菊 陈长明 曹必好 邹丽芳 雷建军*
(华南农业大学园艺学院,广东广州 510642)
芥蓝喜凉性气候,耐高温能力较弱,夏秋季栽培高温逆境往往使其生长发育不良,影响其产量、品质。本试验采用人工气候箱高温处理的方法,测定10份芥蓝材料在不同高温胁迫下的生长指标和生理生化指标,分析10份芥蓝材料在不同高温胁迫下各项指标的相对变化率。结果表明:高温胁迫下芥蓝的热害指数与生长指标(地上部鲜质量、地下部鲜质量、节间长)可以简单、较好地反映出芥蓝材料间耐热性差异;37 ℃/25 ℃、40 ℃/32 ℃处理的生理生化指标相对变化率能较好地反映出芥蓝材料间的耐热性差异;高温能诱导MBF1c转录因子的表达,使其表达量升高,且耐热性强的材料相对表达量较高,该转录因子的表达可能对芥蓝耐热性起到一个正调控作用。综合评价结果表明,矮脚香菇在这10份芥蓝材料中耐热性最强。
芥蓝;耐热性;MBF1c
芥蓝(Brassica alboglabra L. H. Bailey)是十字花科芸薹属一、二年生草本植物,在我国广东、福建等地广为种植;以肥嫩的花薹、嫩叶或嫩株作为主要食用器官,营养价值丰富、质地脆嫩,可炒食或凉拌;已传入日本等东南亚各国和美洲、大洋洲、欧洲等地区(秦耀国 等,2009)。芥蓝是喜凉性蔬菜作物,在高温胁迫下生长和发育都会受到不同程度的影响,一般在秋冬季温度较低的时候栽培,早春也可以栽培,夏季栽培由于温度较高,虽然也能生长,但是产量比较低、品质会变差(杨暹和杨运英,2002)。如果能通过品种选育方法选育出耐高温的芥蓝品种,在高温条件下种植其产量、商品性不会受到太大的影响,则可显著提高芥蓝的经济效益。
在植物中MBF1与逆境胁迫有密切的关系,多种逆境都会诱导MBF1表达增强。在烟草和拟南芥中发现,高温和干旱同时处理可以使MBF1基因诱导表达(Ludmila et al.,2002)。在小麦研究中,以TAM107为材料克隆获得了在高温胁迫下上调表达剧烈的乙烯应答转录共激活因子TaMBF1c,通过转化酵母发现在48 ℃高温下重组酵母的耐热性明显强于转空载体(秦丹丹 等,2013)。转录因子MBF1c在拟南芥耐热过程中是十分重要的,它可以在热激反应中控制36种不同转录因子的表达来介导植物对高温的耐性(Suzuki et al.,2008)。
筛选耐热芥蓝原始材料是选育强耐热性芥蓝品种最重要的环节。以往在大白菜、萝卜、辣椒、黄瓜等蔬菜作物上的耐热性研究较多,并取得了很大的进展,但关于芥蓝耐热性材料筛选的研究鲜见报道。本试验在前人研究基础上,对10份芥蓝材料的与耐热性有关的生长指标和生理生化指标进行了测定,筛选出耐热芥蓝材料,以期为选育耐热性强的芥蓝品种提供参考。
1 材料与方法
1.1试验材料
试验于2015年4月至2016年2月在华南农业大学园艺学院进行。供试材料为芥蓝品种中花芥蓝(1号)、小香菇(22号)、矮脚香菇(25号)、长叶早种(32号)、快大黄花芥蓝(36号)、粗条芥蓝(37号)、雏叶芥蓝(44号)、新会芥蓝(48号)、早格蓝(49号)及通过游离小孢子培养获得的59号,均为多代自交获得的稳定自交系。
1.2试验方法
1.2.1 高温对芥蓝幼苗生长量的影响 2015年11月,将10份芥蓝材料分别播种于72孔育苗盘中,在适宜温度(24 ℃)条件下生长至5~7片真叶;移入光照培养箱中,进行37 ℃/25 ℃(昼/夜,下同)高温处理;高温处理7 d后,每处理随机选取3株幼苗,测定地上部、地下部鲜质量及节间长。以在适温(24 ℃/12 ℃)条件下生长的各相应芥蓝材料为对照,每处理3次重复。
1.2.2 热害指数鉴定 2015年11月,将10份芥蓝材料分别播种于72孔育苗盘中,在适宜温度(24℃)条件下生长至4~5片真叶;每份材料取30株幼苗,分成3次重复,采用完全随机排列的方法置于光照培养箱中,进行40 ℃、12 h高温处理;处理4 d后,调查全部幼苗的热害情况,计算热害指数。热害指数越高,表明该材料耐高温胁迫的能力越差。
参考尹贤贵等(2001)的方法,制定热害分级标准:0级,无黄叶出现,没有热害症状;1级,有1片黄叶出现;3级,有2片黄叶出现;5级,有3片黄叶出现;7级,有4~5片黄叶出现;9级,整株死亡。

1.2.3 高温对芥蓝幼苗生理生化指标的影响 2015年4月,将10份芥蓝材料分别播种于72孔育苗盘中,在适宜温度(24 ℃)条件下生长至5~6片真叶;然后放入人工气候培养箱中,设置3个高温处理水平,分别为34 ℃/20 ℃、37 ℃/25 ℃、40℃/32 ℃,以24 ℃/12 ℃为对照,每处理3次重复;高温处理3 d后,每处理随机选取6株幼苗,测定生理生化指标。丙二醛含量测定采用硫代巴比妥酸法;脯氨酸含量测定采用酸性茚三酮法;可溶性蛋白含量测定采用考马斯亮蓝法;电导率测定采用电导仪;超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)法;过氧化物酶(POD)活性测定采用愈创木酚法;过氧化氢酶(CAT)活性测定采用紫外吸收法(李合生,2000)。
1.2.4 实时荧光定量PCR检测 将10份芥蓝材料幼苗(5~6片真叶)放入人工气候培养箱中,每份材料10株,37 ℃高温处理1 h;每份材料随机选取3株,剪取相同部位叶片,提取RNA并反转成cDNA。以24 ℃/12 ℃条件下生长的各相应芥蓝材料为对照。
PCR扩 增 体 系(10 μL):SYBR Premix Ex TagⅡ(2 x)5 μL,Actin为内参基因,特异引物(上游引物:5′-AAAAGCGGTAAACCAAGC-3′,下游引物:5′-GTCTCACATCCACAGGCA-3′)各0.5 μL,稀释后的cDNA(50 ng)1 μL,ddH2O 3 μL。
PCR扩增程序:采用两步扩增法,94 ℃预变性30 s;94 ℃变性5 s,60 ℃退火30 s,40个循环,在每个循环结束后进行荧光信号采集;所有循环结束后进行溶解曲线分析(65~95 ℃)。以高温处理和对照样品的cDNA(稀释后)为模板进行实时定量PCR检测,每个样品进行3次技术重复,同时设置无模板的阴性对照。
1.2.5 综合评价 依照盛业龙等(2014)采用的模糊数学中的隶属函数数值法,计算10份芥蓝材料各指标的隶属函数值,并进行累加,综合比较各芥蓝材料的耐高温胁迫能力。

式中:X为某芥蓝材料某一指标的测定值;Xmax为所有参试材料某一指标测定值的最大值;Xmin为所有参试材料某一指标测定值的最小值。
本试验中,不同处理各项指标相对值均用该指标的变化率表示。
相对变化率=(处理-对照)/对照×100%
1.3数据处理
采用WPS软件和SPASS软件对试验数据进行整理及显著性测验、多重比较等。
2 结果与分析
2.1高温对芥蓝幼苗生长量的影响
从表1可以看出,供试各芥蓝材料经37 ℃高温处理后生长量相对变化率存在显著差异。59号地上部鲜质量、地下部鲜质量、节间长的相对变化率分别为25.16%、28.57%、27.42%,均高于其他材料;25号地上部鲜质量、地下部鲜质量、节间长的相对变化率分别为2.27%、4.35%、4.71%,是10份芥蓝材料中最低的;22号和1号地上部鲜质量、地下部鲜质量、节间长的相对变化率也较低。害指数分别为63%和58%;1号、22号、25号受热害影响较轻,热害指数分别为19%、13%、5%;除32号和44号材料间热害指数无显著差异外,其他各材料间差异均达显著水平。
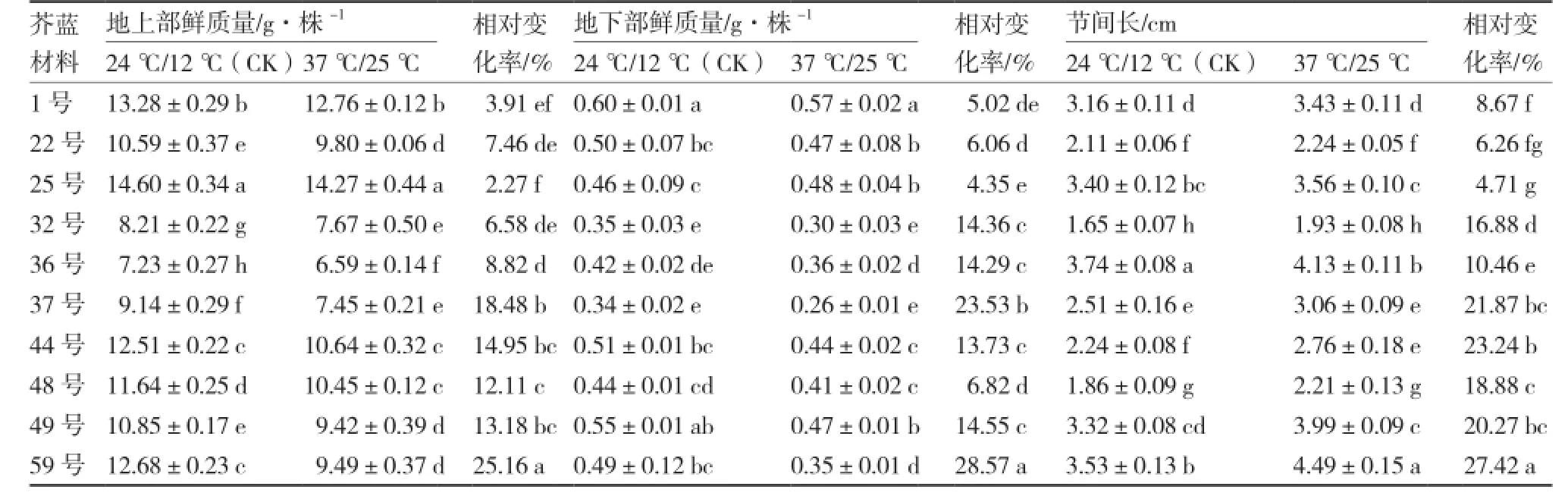
表1 高温对芥蓝幼苗生长量的影响
2.2热害指数鉴定结果
由图1可见,参试各芥蓝材料间热害指数差异显著,热害表现最明显的是59号,热害指数高达66%;36号、37号受热害胁迫也比较严重,热
相对变化率如图3所示。34 ℃/20 ℃处理下,相对变化率最大的是59号,为223%;相对变化率较小的是25号和22号,分别为22%和26%。37℃/25 ℃处理下,相对变化率最大的还是59号,
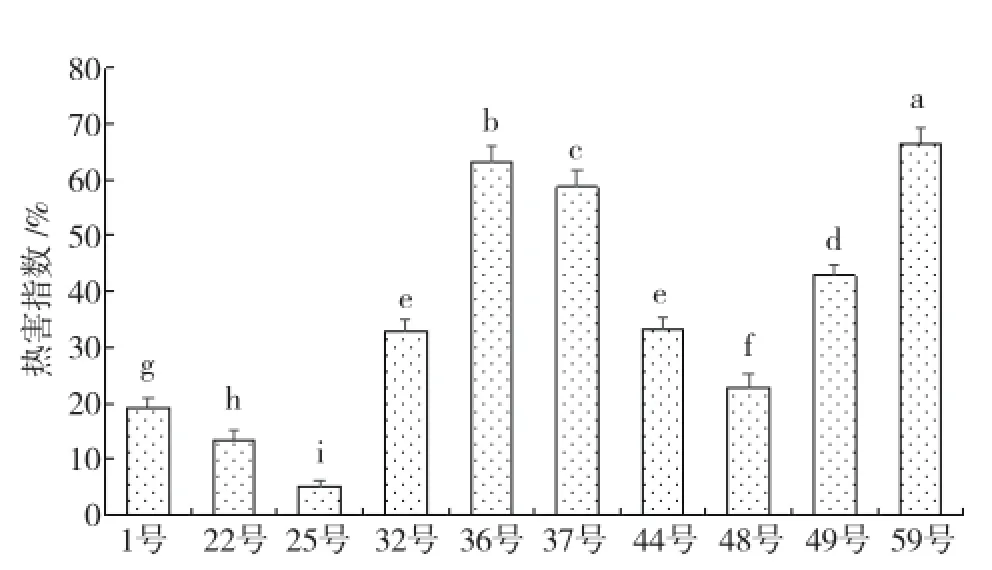
图1 不同芥蓝材料的热害指数
2.3高温对芥蓝幼苗生理生化指标的影响
由图2可见,随着温度的升高,各芥蓝材料的相对电解质渗透率均逐渐升高;在24 ℃/12 ℃条件下各材料间相对电解质渗透率相差不大,但随着温度的升高各材料间相对电解质渗透率的差异越来越大。从增长趋势来看,24 ℃/12 ℃到34 ℃/20 ℃、37 ℃/25 ℃到40 ℃/32 ℃这两个温度段间相对电解质渗透率的增加幅度较大,34 ℃/20 ℃到37 ℃/25℃温度段间增加幅度较为平缓。

图2 不同芥蓝材料高温胁迫下的相对电解质渗透率
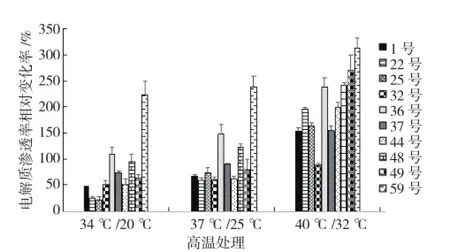
图3 不同芥蓝材料高温胁迫下相对电解质渗透率的相对变化率
各芥蓝材料高温胁迫下相对电解质渗透率的为238%;相对变化率较小的是22号、32号和44号,分别为59%、60%和62%。40 ℃/32 ℃处理下,相对变化率最大的亦是59号,为324%;相对变化率最小的是32号,为88%。
从表2可以看出,无论是在适温下还是在高温下,各芥蓝材料间脯氨酸含量都有显著差异;随着温度的升高,各材料脯氨酸含量都呈增加的趋势。3个高温处理下25号的脯氨酸含量相对变化率均为最高,分别为163.40%、625.06%、731.12%。34℃/20 ℃处理下,22号、25号、37号、44号的脯氨酸含量相对变化率显著高于其他材料;37 ℃/25℃处理下,脯氨酸含量相对变化率最高的是25号,其次是59号、32号、44号、1号,最小的是36号;40 ℃/32 ℃处理下,脯氨酸含量相对变化率由高到低依次为:25号、44号、1号、22号、32号、59号、49号、36号、48号、37号;25号、44号和1号在37 ℃/25 ℃和40 ℃/32 ℃处理下脯氨酸含量相对变化率都较高。
从表3可以看出,随着温度的升高,各芥蓝材料的丙二醛含量都有不同程度的升高,且各材料间丙二醛含量差异显著;59号在3个高温处理下丙二醛含量都较高,而25号的丙二醛含量都较低。37 ℃/25 ℃处理下,各材料的丙二醛含量相对变化率由高到低依次为:59号、36号、37号、44号、32号、49号、1号、48号、22号、25号;其中,59号、36号、37号的丙二醛含量相对变化率较高,25号和22号较低。
从表4可以看出,各芥蓝材料的可溶性蛋白含量均随温度的升高而逐渐递增,且不同材料间可溶性蛋白含量差异显著,但是变化幅度不一;各温度处理下59号的可溶性蛋白含量都较低,而37号的可溶性蛋白含量都较高。34 ℃/20 ℃处理下,可溶性蛋白含量相对变化率较高的是25号和44号,分别为95.41%和94.17%,而32号和49号的相对变化率较低;37 ℃/25 ℃处理下,可溶性蛋白含量相对变化率最高的是32号,为128.82%,相对变化率最低的是49号;40 ℃/32 ℃处理下,各材料的可溶性蛋白含量相对变化率呈增加的趋势,25号和32号的相对变化率较高,分别为130.73%和131.42%,而49号的相对变化率最低。
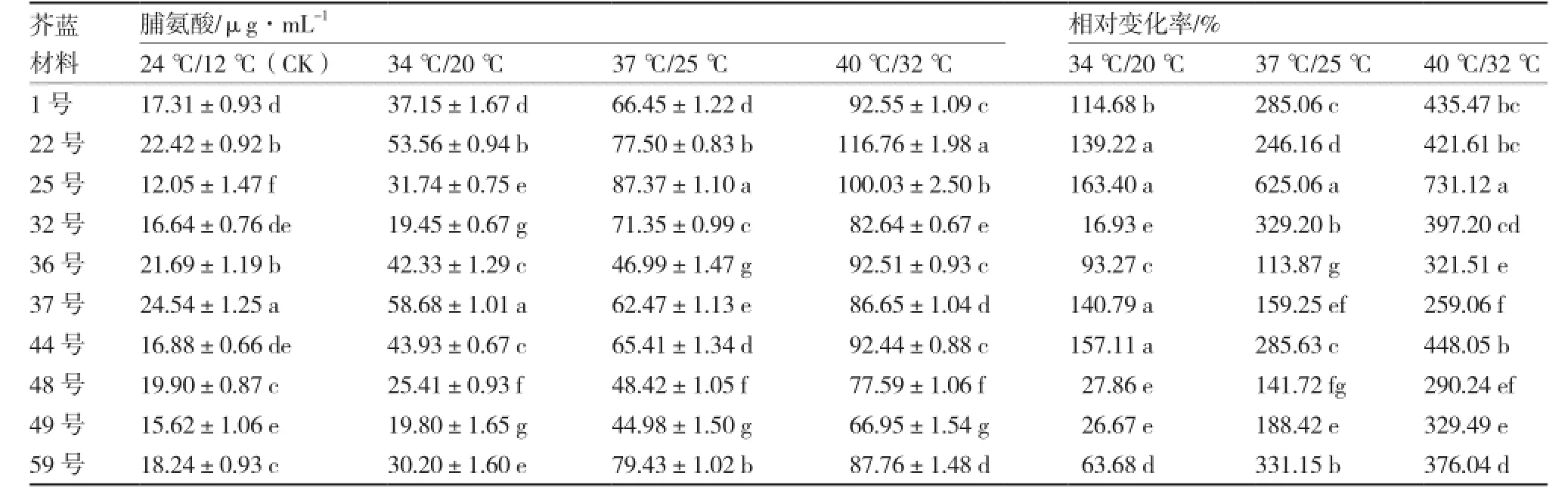
表2 不同芥蓝材料高温胁迫下脯氨酸含量及相对变化率
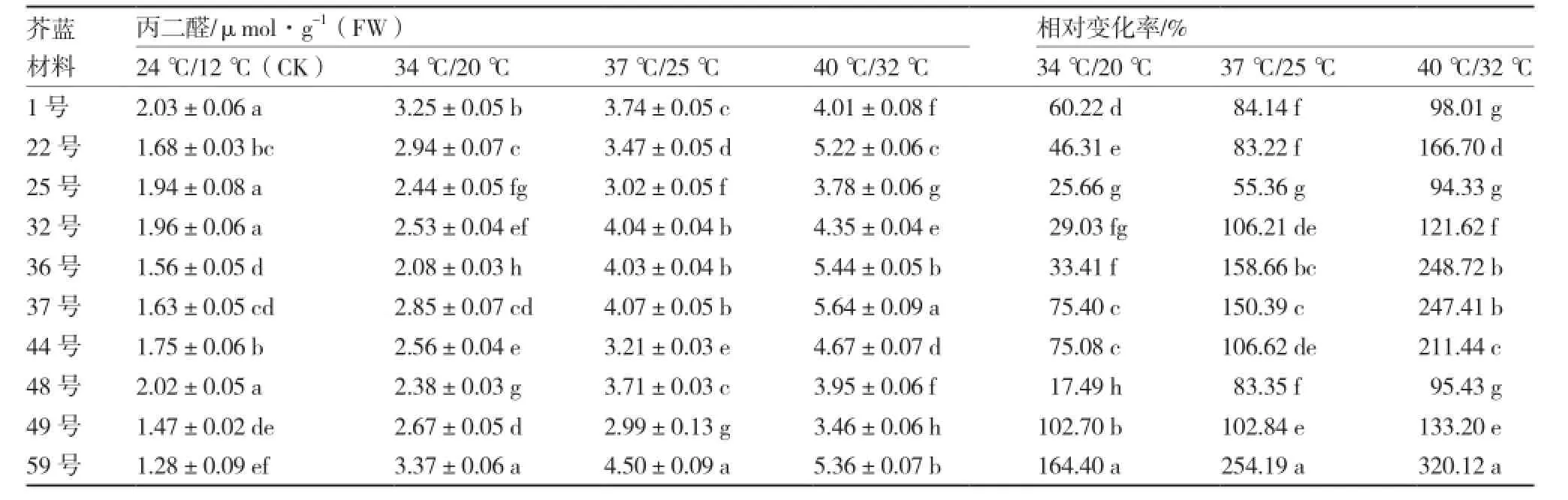
表3 不同芥蓝材料高温胁迫下丙二醛含量及相对变化率
从表5可以看出,各个温度下不同芥蓝材料间SOD活性都有显著差异,且随着温度的升高各材料的SOD活性均呈逐渐上升的趋势。25号和1号在3个高温处理下的SOD活性相对变化率都较高,分别为197.08%、232.61%、338.36%和102.67%、429.68%、481.54%;而59号的SOD活性相对变化率最低;40 ℃/32 ℃处理下,各芥蓝材料SOD活性相对变化率由高到低依次为:1号、25号、48号、32号、22号、37号、44号、36号、49号、59号。
不同温度条件下各芥蓝材料的POD活性变化如表6所示。各材料的POD活性均随温度的升高呈上升的趋势,且各材料间POD活性差异显著。25号在24 ℃/12 ℃处理下的POD活性不高,只有1.33 μg·g-1,但在高温胁迫下酶活性急剧上升,34 ℃/20 ℃、37 ℃/25 ℃、40 ℃/32 ℃处理下的POD活性分别为3.97、6.21、8.71 μg·g-1,相对变化率分别达198.92%、366.76%、557.47%;其次是22号,37 ℃/25 ℃、40 ℃/32 ℃处理下的POD活性相对变化率分别为198.25%和222.65%;59号在3个高温处理下的POD活性相对变化率都较低。
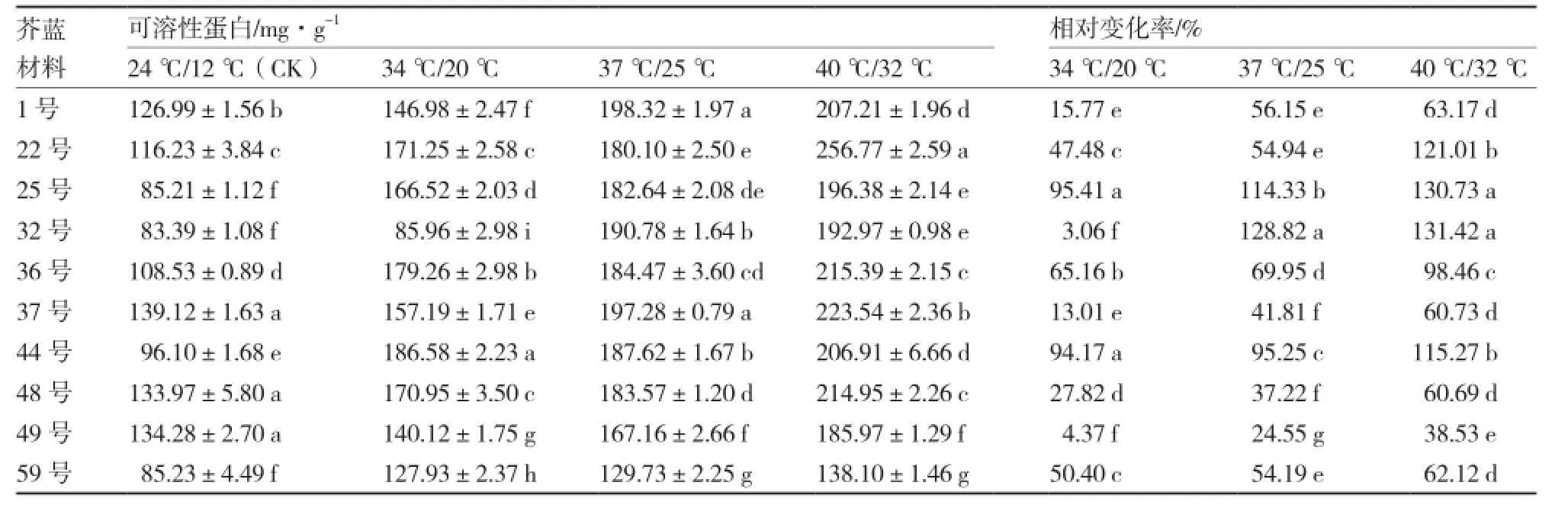
表4 不同芥蓝材料高温胁迫下可溶性蛋白含量及相对变化率
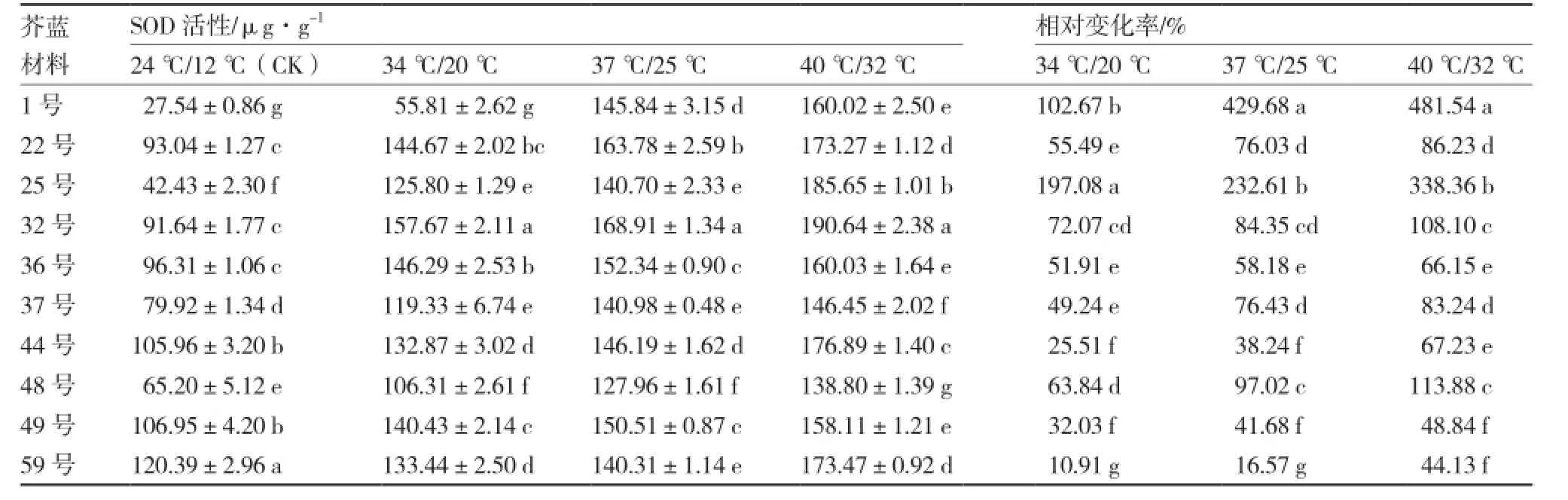
表5 不同芥蓝材料高温胁迫下SOD活性及相对变化率
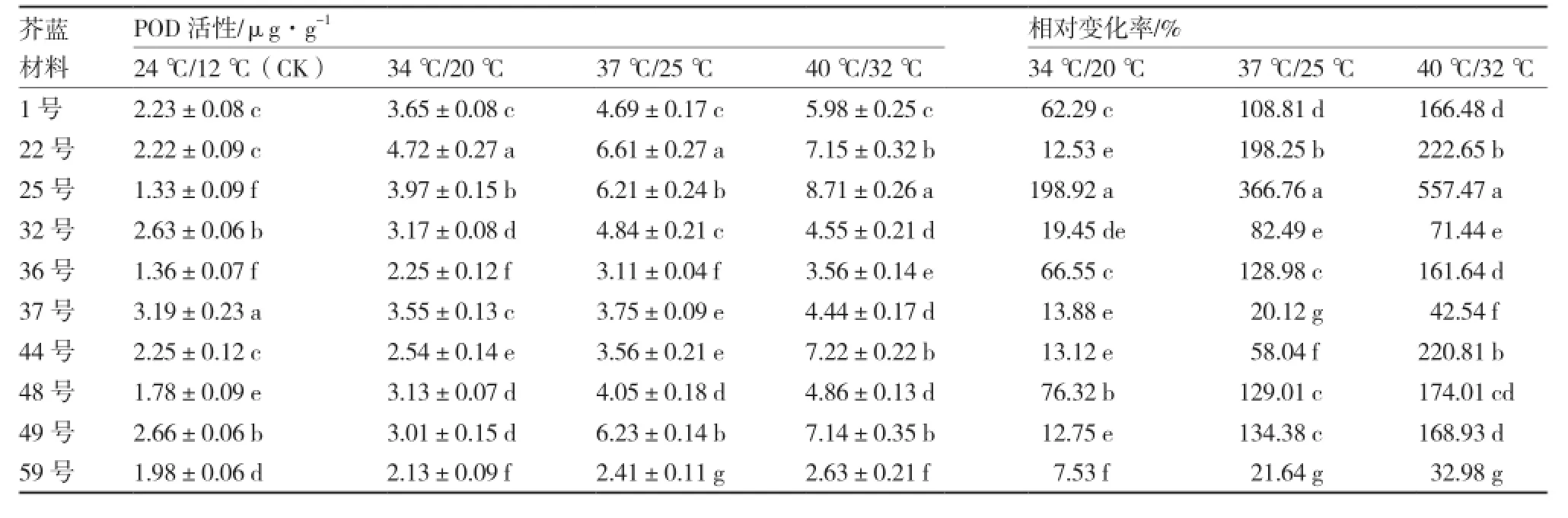
表6 不同芥蓝材料高温胁迫下POD活性及相对变化率
从表7可以看出,参试10份芥蓝材料的CAT活性均随着温度的升高呈上升趋势,且各材料间CAT活性差异显著;24 ℃/12 ℃处理下,22号的CAT活性最高,其次是25号、36号和1号;3个高温处理下,22号和25号的CAT活性较高,而59号的CAT活性较低。48号在3个高温处理下的CAT活性相对变化率均为最高,分别达到了885.39%、1 051.40%、1 399.73%,而59号的CAT活性相对变化率均较低;40 ℃/32 ℃处理下,各芥蓝材料CAT活性相对变化从大到小依次为:48号、22号、25号、44号、1号、36号、49号、32号、37号、59号。
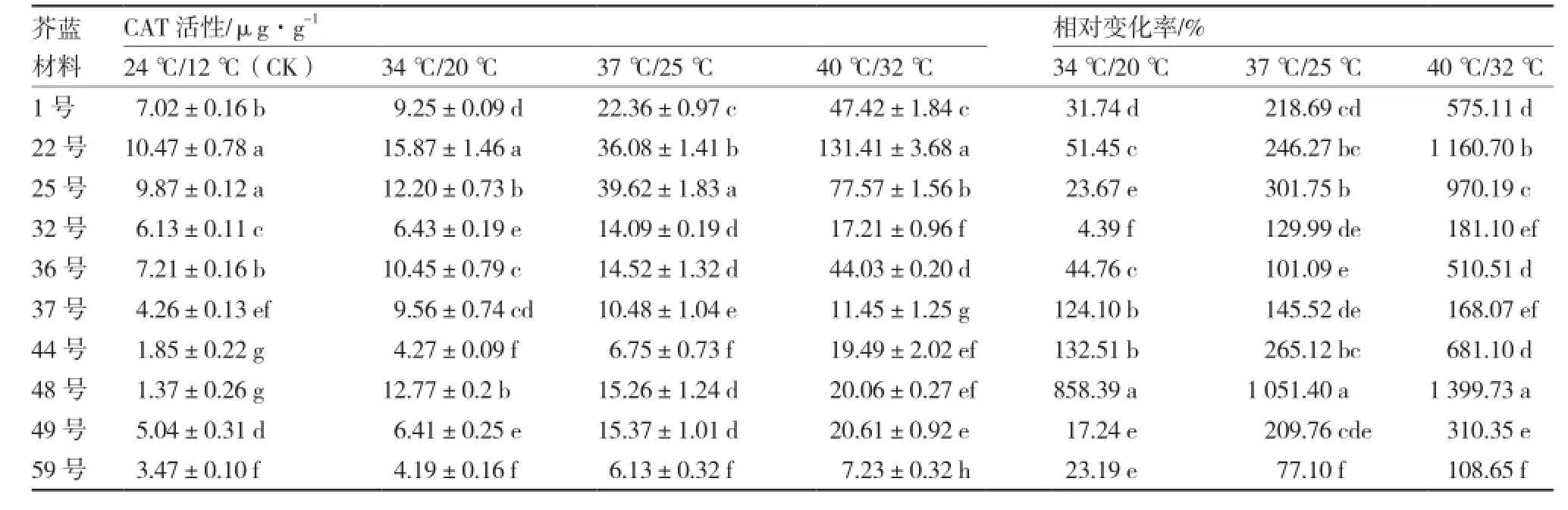
表7 不同芥蓝材料高温胁迫下CAT活性及相对变化率
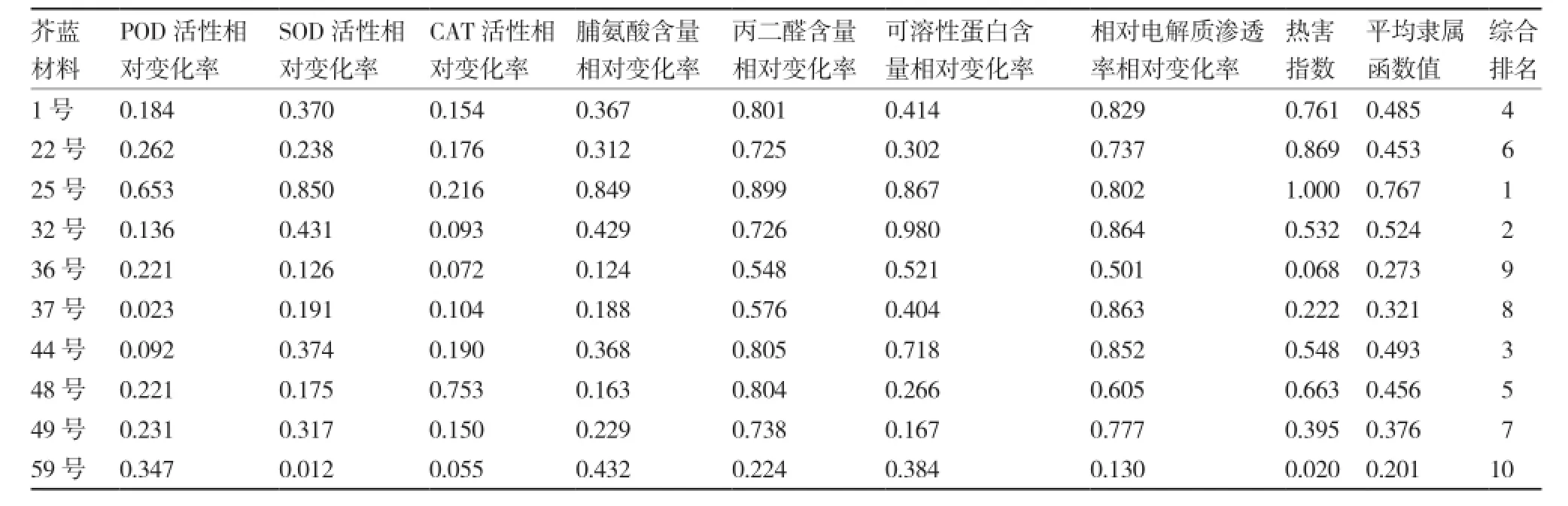
表8 不同芥蓝材料高温胁迫下生理生化指标相对变化率的隶属函数值
2.4隶属函数综合法评价结果
综合各测定指标的隶属函数值,25号综合排名第1,59号综合排名最后;各芥蓝材料的耐热性由高到低依次为:25号、32号、44号、1号、48号、22号、49号、37号、36号、59号(表8)。
2.5高温对芥蓝幼苗MBF1c表达量的影响
由图4可见,37 ℃高温处理1 h后,除37号外其他各芥蓝材料的MBF1c表达量都呈增加趋势;25号在受到高温胁迫后,MBF1c表达量急剧上升,上升幅度最大。
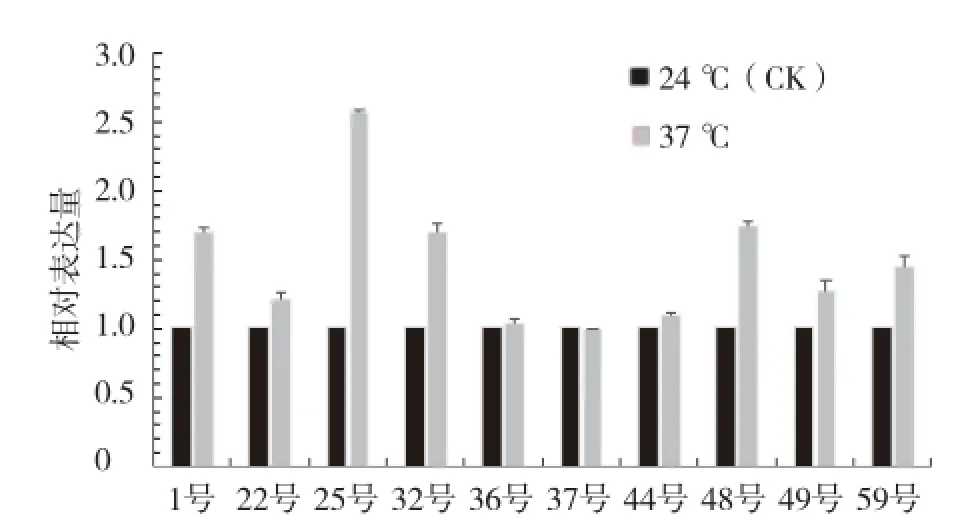
图4 高温胁迫对芥蓝幼苗MBF1c表达量的影响
3 结论与讨论
高温胁迫能使植物生长受阻,使植物的生长量减少,叶片变黄枯萎,幼苗徒长,根系发育受阻;此外,在热胁迫下通过测定植物生理生化指标的变化也可以鉴定植物的耐热性,且在大白菜(司家钢等,1995)、番茄(王冬梅,2003)、萝卜(梁超英,2006)、黄瓜(李建建,2006)上都有报道。
本试验结果表明,高温胁迫会使芥蓝幼苗徒长,且耐热材料的相对变化率要小于不耐热材料。这与刘燕燕等(2005)在不结球白菜、陈日远等(2006)在芥蓝上的研究结果一致。经过40 ℃高温处理后,10份芥蓝材料都出现了不同程度的热害症状。比较热害指数与生长量的相对变化率,发现高温对芥蓝的生长量有一定影响,热害指数低的芥蓝材料地上部、地下部鲜质量减少量较小,节间长增加量较大;热害指数高的芥蓝材料地上部、地下部鲜质量减少量较大,节间长增加量较小。
电解质渗透率在植物耐热性鉴定中应用较早,是能反映植物耐热性的一个重要指标。高温胁迫下植物细胞膜易受到伤害,导致细胞膜通透性增加,电解质外渗,电导率升高,从而使植物体受到损伤(潘光辉 等,2001)。本试验中,不同高温处理下,24 ℃/12 ℃到34 ℃/20 ℃、37 ℃/25 ℃到40 ℃/32℃这两个温度段间电导率增长幅度比34 ℃/20 ℃到37 ℃/25 ℃温度段间的大,前者可能是由于温度区间跨度大,细胞膜受损严重;后者则可能是因为处理温度较高,细胞膜受损更严重。研究各芥蓝材料的电解质渗透率在不同温度条件下的变化规律发现,在34 ℃/20 ℃处理下各芥蓝材料的电解质渗透率相对变化率排列顺序与热害指数大致相符,采用此温度下测定的电解质渗透率能简单地反映出芥蓝的耐热性。
在高温胁迫下,丙二醛含量的积累量变化反映了细胞膜脂过氧化程度,在一定程度上反映了植物的耐热性;耐热性强的芥蓝材料丙二醛含量的积累量变化小,耐热性较差的芥蓝材料丙二醛含量的积累量变化大。本试验中发现:在高温胁迫下,耐热性强的芥蓝材料脯氨酸含量相对变化率较大,耐热性差的芥蓝材料相对变化率较小,说明在高温胁迫下细胞中会大量积累脯氨酸,作为渗透调节物质使细胞结构稳定,对作物起到一定的保护作用。
本试验通过荧光定量法测定MBF1c的表达量,发现在短时间高温处理后,9份芥蓝材料的MBF1c表达量都有不同程度的升高,说明高温会诱导MBF1c表达;耐热性强的芥蓝材料表达量上升幅度更大,说明MBF1c可能对芥蓝的耐热性起正调控作用。如果能在沉默MBF1c后再对各芥蓝材料的耐热性指标进行一次综合鉴定,观察各芥蓝材料在MBF1c沉默后各项指标是否发生相应的变化,更能进一步说明MBF1c与芥蓝耐热性的关系。
植物的耐热性往往是由外界环境温度和植物体内许多复杂代谢因素共同作用的结果,不能单凭某一指标就判定植物的耐热性。高温胁迫对植物的影响最直观的是表现在外部形态和生长情况上,而在植物体内由于不同耐热机制发挥的效果不同而导致各项生理指标对鉴定耐热性的贡献有所不同。本试验全面地测定了与植物耐热性有关的生长指标和生理指标,设置不同胁迫温度用于探索能简单准确地反映出芥蓝耐热性的方法,发现室内鉴定中各指标的绝对含量不能较好的反映出芥蓝材料间的耐热性差异,而使用相对变化率较为准确。综合评价结果表明,矮脚香菇(25号)在这10份芥蓝材料中耐热性最强。
陈日远,刘厚诚,刘晴,黄丹凤.2006.高温对芥蓝苗期生长及菜薹产量和品质的影响.青岛:中国园艺学会第七届青年学术讨论会.
李合生.2000.植物生理生化实验原理和技术.北京:高等教育出版社.
李建建.2006.高温胁迫对黄瓜幼苗生理生化特性影响及其生理机制的研究〔硕士论文〕.兰州:甘肃农业大学.
梁超英.2006.萝卜耐热鉴定和叶片生化物指标含量研究〔硕士论文〕.杨凌:西北农林科技大学.
刘燕燕,沈火林,刘以前.2005.高温胁迫对不结球白菜幼苗生长及生理指标的影响.华北农学报,20(5):25-29.
潘光辉,王文强,尹贤贵,张赟,杨琦凤.2001.电导法鉴定番茄耐热性与田间结果的比较.西南园艺,29(2):26-27.
秦丹丹,谢颂朝,刘刚,姚颖根,孙其信,彭惠茹.2013.小麦中编码未知蛋白的热胁迫响应基因TaWTF1的克隆和功能分析.植物学报,48(1):34-41.
秦耀国,杨翠芹,曹必好,陈国菊,雷建军.2009.芥蓝遗传育种与生物技术研究进展.中国农学通报,(18):296-299.
盛亚龙,王莎莎,许美玲.2014.应用隶属函数法综合评价不同烤烟品种苗期抗旱性.南方农业学报,(10):1751-1758.
司家钢,孙日飞,吴飞燕,钮心恪.1995.高温胁迫对大白菜耐热性相关生理指标的影响.中国蔬菜,(4):4-6.
王冬梅.2003.番茄耐热性鉴定方法的研究及其种质资源的筛选鉴定〔硕士论文〕.哈尔滨:东北农业大学.
杨暹,杨运英.2002.温度对芥蓝生长发育与菜薹形成的影响.中国蔬菜,(4):33-34.
尹贤贵,罗庆熙,王文强,张赟,潘光辉,杨琦凤,尹诗麟.2001.番茄耐热性鉴定方法研究.西南农业学报,14(2):62-65.
Ludmila R,Hongjian L,Ron M.2002.The combined effect of drought stress and heat shock on gene expression in tobacco.Plant Physiology,130(3):1143-1151.
Suzuki N,Bajad S,Shuman J,Shulaev V,Mittler R.2008.The transcriptional co-activator MBF1c is a key regulator of thermotolerance in Arabidopsis thaliana.Journal of Biological Chemistry,283(14):9269-9275.
Identification of Heat Tolerance in Chinese Kale and the Expression Analysis of Heat Tolerance Transcription Factor MBF1c
WANG Hao-wei,CHEN Guo-ju,CHEN Chang-ming,CAO Bi-hao,ZOU Li-fang,LEI Jian-jun*
(College of Horticulture,South China Agricultural University,Guangzhou 510642,Guangdong,China)
Chinese kale(Brassica alboglabra L. H. Bailey)is a thermophilic vegetable,which can’t well grow at high temperature.High temperaturewould prevent Chinese kale from growing and result in poor quality of its seedlings,seriously influence yield and quality in later period in summer and autumn.Taking 10 Chisnses kale lines as experimental materials,the growth indexs and physiology indexs under different high temperature stress were measured.The relative change rate of 10 Chinese kale lines under different high temperature stress was analyzed.The results showed that heat damage index and relative growth could be used to determine the heattolerance ability of Chinese kale lines,which can be most quick and convenient.The difference in heat tolerance among the varieties could be indicated by physiological and chemical index under 37℃/25 ℃ or 40℃/32 ℃.After the treatment of 37 ℃for 1 hour the expression variation of MBF1c was increased.According to the comprehensive analysis,‘Aijiaoxianggu’was the most heat-tolerant lines.
Chinese kale;Heat tolerance;MBF1c
王浩伟,男,硕士研究生,专业方向:芥蓝育种及分子生物学,E-mail:503469851@qq.com
*通讯作者(Corresponding author):雷建军,男,教授,博士生导师,专业方向:芥蓝、辣椒育种及分子生物学,E-mail:jjlei@scau.edu.cn
2016-07-16;接受日期:2016-10-16
广东省科技厅项目(2013B051000069,2015B020202009),广州市科创委项目(201508030021)
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
湖南电力(2021年4期)2021-11-05
中国核电(2021年3期)2021-08-13
初中生世界·九年级(2020年9期)2020-09-21
火力与指挥控制(2016年8期)2016-09-21
安徽农学通报(2015年4期)2015-07-30
中学生理科应试(2014年12期)2015-01-15
