
养分化学计量比对稻田土壤葡萄糖矿化及其激发效应的影响*
2017-02-23 05:50:56汤珍珠祝贞科沈冰洁胡亚军王娟庞静童成立葛体达吴金水
土壤学报 2017年1期
汤珍珠祝贞科沈冰洁胡亚军王 娟庞 静童成立葛体达吴金水
(1 湖北大学资源环境学院,武汉 430062)
(2 中国科学院亚热带农业生态研究所农业生态系统过程重点实验室,长沙 410125)
(3 中南林业科技大学环境科学与工程学院,长沙 410004)
养分化学计量比对稻田土壤葡萄糖矿化及其激发效应的影响*
汤珍珠1,2祝贞科2†沈冰洁2,3胡亚军2王 娟2庞 静1†童成立2葛体达2吴金水2
(1 湖北大学资源环境学院,武汉 430062)
(2 中国科学院亚热带农业生态研究所农业生态系统过程重点实验室,长沙 410125)
(3 中南林业科技大学环境科学与工程学院,长沙 410004)
选择13C-葡萄糖作为稻田土壤典型易利用态外源有机碳,通过室内培养试验,研究不同C/N/P/S计量比条件下,葡萄糖分解矿化的动态规律及其激发效应。结果表明,稻田土壤中葡萄糖-碳(C)快速矿化,60 d培养实验后,有65.5%~74.6%的葡萄糖-C矿化。养分元素的添加使土壤中葡萄糖-C快速转化碳库的比例逐渐由58%增加至65%,从而使葡萄糖-C矿化率提高了3.9%~12.5%,养分元素的添加量与葡萄糖-C快速转化碳库的比例和矿化率均表现出显著的正相关关系(R2= 0.63,p< 0.05;R2= 0.83,p<0.05)。葡萄糖-C矿化过程中,导致稻田土壤碳的累积负激发效应为-370~-570 mg kg-1,养分元素添加比例越大,其负激发效应越强,二者呈显著的负相关性(R2= 0.66,p<0.05)。研究表明,稻田土壤中易利用态碳的矿化受C/N/P/S元素计量比的影响,高比例养分元素的添加,促进土壤中易利用态碳的矿化,抑制土壤原有有机质的分解,增强负激发效应。本研究可为深入了解稻田生态系统碳循环、实现农田土壤肥力提升和温室气体减排提供理论依据。
稻田土壤;有机碳矿化;激发效应;养分元素计量学
稻田土壤有机质矿化过程是稻田土壤碳(C)循环最基本的过程之一,土壤中易利用态有机碳库通过调节土壤能源物质供应和微生物活性影响土壤碳库的有效性,与土壤生态系统中有机碳的迁移、固持和温室气体的释放具有密切的联系,成为土壤有机碳周转的动力和土壤质量演变的关键指标[1-3]。微生物分解有机质导致了有机碳的转化与呼吸利用,这同样发生于稻田土壤有机质的矿化等养分循环过程中[4-5]。生态系统的养分状态强烈影响微生物营养物质的浓度和计量比。生态系统碳循环和碳固定效率取决于关键养分元素输入量和需求之间的平衡,以及养分的有效性[6-8]。稻田生态系统碳循环的稳定性不仅会受到相关生物体对元素需求的强烈影响,也会受到周围环境化学元素平衡状况的影响[9],在相对稳定的条件下,稻田生态系统碳循环的微生物过程是由质量守恒原理和其他关键养分元素(如氮(N)、磷(P)、硫(S)等)的计量比控制的[7,10-11]。在外源输入大量有机碳条件下,稻田土壤微生物通过增加养分元素水解酶的生产力,获取满足它们生长需要的C、N和P等元素,达到适宜的生态化学元素计量比,从而调控土壤有机质的分解与转化[12]。所以,生物体对关键养分元素(如N、P)需求的改变将明显影响生态系统碳循环的过程[7,13]。
人类活动对C、N、P生物地球化学循环的影响逐渐加剧,而碳素的生物地球化学循环也取决于其和N、P等养分元素间的平衡关系,基于生态化学计量学原理,土壤碳固定取决于N和P对微生物的影响[14-16]。由于土壤 C/N/P比是反映土壤内部C、N、P循环的主要指标,综合了生态系统功能的变异性,容易测量,而且有助于确定生态过程对全球变化的响应,因而成为确定土壤C、N、P平衡特征的一个重要参数,而且凋落物和土壤C/N/ P比值也可成为C、N、P矿化作用和固持作用的指标[17-18]。因此,根据生态化学计量学原理,调节稻田土壤中养分元素与碳素的计量比,对于调控稻田土壤碳固持、减少温室气体排放具有重要的理论意义和应用价值。
土壤碳尤其是易利用态碳在土壤碳库的周转过程中起着关键的作用,葡萄糖作为土壤中植物根际沉积碳以及外源输入的低分子量有机碳的典型代表,研究其在不同养分元素计量比条件下的周转,对探究外源碳在土壤碳库中的转化动态及与N、P、S养分元素的耦合关系,以及揭示稻田外源碳在不同C/N/P计量比条件下矿化的元素计量学调控机制,均具有重要的科学意义。本研究选择13C-葡萄糖作为稻田土壤典型易利用态外源有机碳,研究在不同C/N/P/S计量比营养元素添加条件下,葡萄糖分解矿化的动态规律,通过定量13C-CO2的生成动态,以及对土壤原有有机质的激发效应,探究土壤碳转化的元素计量学调控机制。
1 材料与方法
1.1 供试土壤
为了考察不同氮磷养分元素添加水平下,土壤碳周转的计量学特征,本研究选择位于中国科学院桃源农业生态试验站(111°26′26.8″~26′28.7″E,28°55′47.8″~55′48.3″N)长期种植水稻的低磷土壤,定位试验始于2000年。所采土壤属于母质为第四纪红黏土的水稻土(简育水耕人为土)、熟化程度基本相近的典型水稻土耕作层(0~20 cm)土壤,土壤13C自然丰度为-29.11‰。该地区属典型亚热带湿润气候,海拔92.2~125.3 m,年均气温16. 5℃,年均降水量1 400 mm。土壤均用直径为5 cm 的不锈钢土钻采集。运回实验室后的土壤样品取 1 kg 室内风干,分别过0.25 mm 和0.149 mm 筛,用于测定土壤基本理化性质。剩余土样风干后过2 mm筛,备用。供试土壤基本理化性质见表 1。
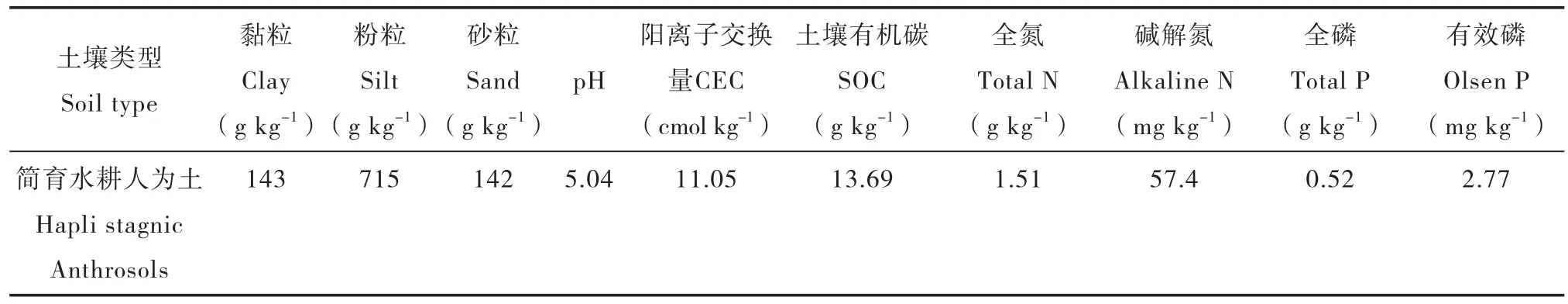
表1 供试土壤基本理化性质Table 1 Properties of the soil used in the experiment
1.2 土壤培养实验
实验开始前,将过2 mm筛的土壤置于50 L 塑料桶中,淹水2~3 cm,25℃恒温预培养14 d。称取预培养后混匀的土壤10 g(干土计)于500 ml厌氧瓶中,加入一定量的去离子水,使水面保持2~3 cm,每瓶添加葡萄糖为17.4 mg,葡萄糖-C的添加量相当于5% 土壤有机碳(SOC),13C-葡萄糖的丰度(atom%13C)为5%(Cambridge isotope laboratories,Tewksbury,MA,美国)。养分元素的添加按照表2中所列的添加量,依次在处理2~6中分别以NH4NO3、KH2PO4和Ca2SO4的形式加入N、P、S养分元素,其中葡萄糖、NH4NO3、KH2PO4和Ca2SO4均以水溶液的形式加入。最后,用玻璃棒将土水混合均匀,并用去离子水冲洗玻璃棒和瓶壁,加盖培养。
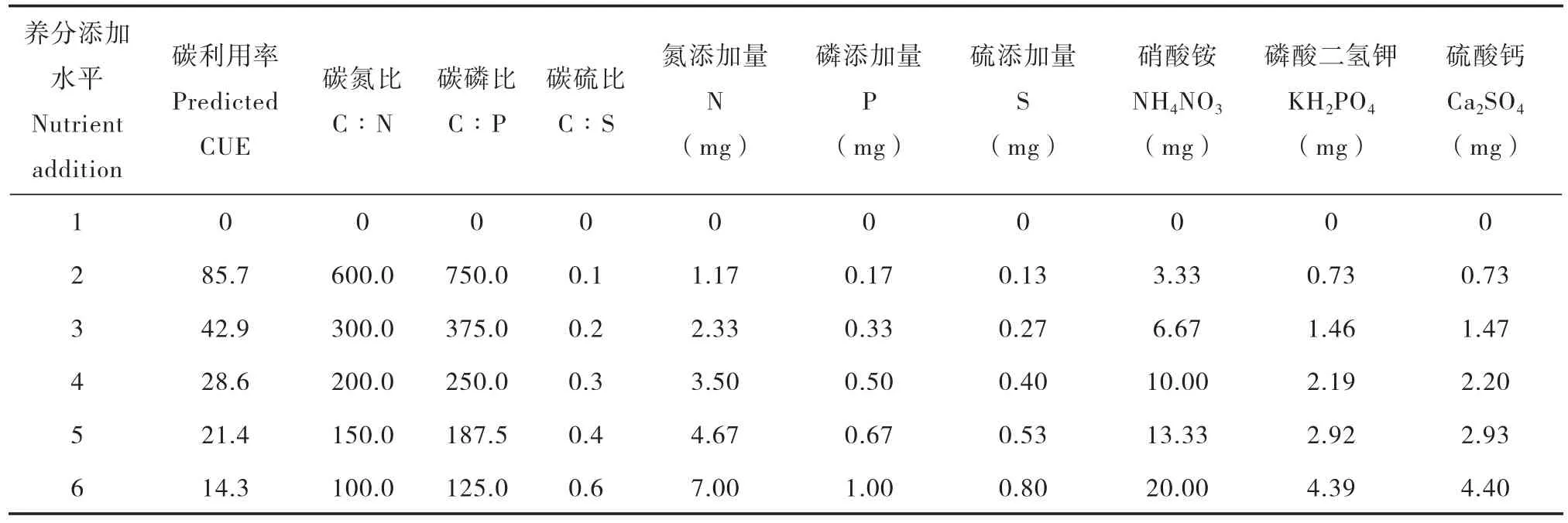
表2 不同养分元素添加水平下,氮磷硫元素添加量与碳/养分元素的计量比Table 2 Relationship of N,P,and S addition rate with and stoichiometric ratio of C/nutrients
以土壤中不加葡萄糖和养分元素作为对照(CK),并设置在土壤中加入12C-葡萄糖处理,作为自然丰度对照,用以计算生成的气体中13C含量。每处理设置4个重复。培养实验在25℃恒温培养室中进行。
1.3 测定方法
土壤pH 测定以水为浸提剂,水土比为2.5∶1;土壤黏粒含量采用比重计法测定;阳离子交换量采用乙酸铵交换法测定;土壤有机碳和全氮采用碳氮元素分析仪(VARIO MAX C /N,德国)测定(干烧法);土壤全磷用微波消煮法测定;碱解氮采用碱解扩散法测定;有效磷采用碳酸氢钠提取―钼锑钪比色法测定;速效钾采用乙酸铵提取―火焰分光光度计测定。
根据预实验结果,实验前20 d气体产生速率较快,随后逐渐减慢,因此设置采气时间为,实验前10 d为每天采气一次,10~20 d为每2天采气一次,最后分别在第25天、30天和60天用注射器采集30 ml厌氧瓶的顶空中的气体,分别注入2个10 ml真空瓶中,分别测定CO2气体浓度以及CO2中13C丰度。每次采气结束后,向厌氧瓶中鼓空气10 min,然后加盖继续培养。CO2气体浓度用气相色谱(Shimadzu,Kyoto,日本)测定。土壤、葡萄糖和CO2的13C丰度用MAT253同位素质谱仪(Thermo Fisher Scientific,Waltham,美国)测定。
1.4 数据处理
CO2气体的13C含量计算方法如下[5,6]:

式中,(Atom13C%)L和(Atom13C%)UL分别指土壤中加入标记和未标记葡萄糖产生CO2气体的丰度;CCO2为CO2气体的浓度,mg kg-1。
葡萄糖-C的累积矿化量是各取样点CO2生成量的总和,葡萄糖-C的矿化率用13C-CO2的量与加入土壤中葡萄糖-13C总量的比值表示。
激发效应(PE)计算公式为[6,19]:
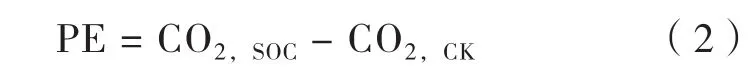
式中,CO2,SOC表示添加葡萄糖处理中来源于土壤有机质(SOC)的CO2的量,mg kg-1;CO2,CK表示对照处理中(CK)CO2的量,mg kg-1。
所有测定结果均为4次重复的平均值,所得数据采用 Origin 8.5和SPSS 16. 0软件进行处理与统计分析。不同处理差异显著性用One-way ANOVA(单因素方差分析)检验,多重比较采用 Duncan 法。
2 结 果
2.1 不同元素计量比条件下土壤外源碳的矿化
土壤碳在模拟田间环境的培养过程中,分解矿化生成一定量的CO2,而养分元素的添加促进了CO2生成。相比不添加葡萄糖土壤(CK)矿化生成的CO2量(3 387 mg kg-1),添加葡萄糖和养分元素后CO2生成量提高1.5倍~1.6倍,而且N、P等养分元素添加比例越高,生成的CO2量越多(图1A)。同时,土壤中添加的葡萄糖快速矿化生成13CO2,60 d培养实验结束时,有138.6 mg kg-1的13CO2生成,而在处理2~6中,随着养分元素添加比例的依次增加,葡萄糖矿化生成13CO2的量提高了1.7%~8.3%(图1B)。
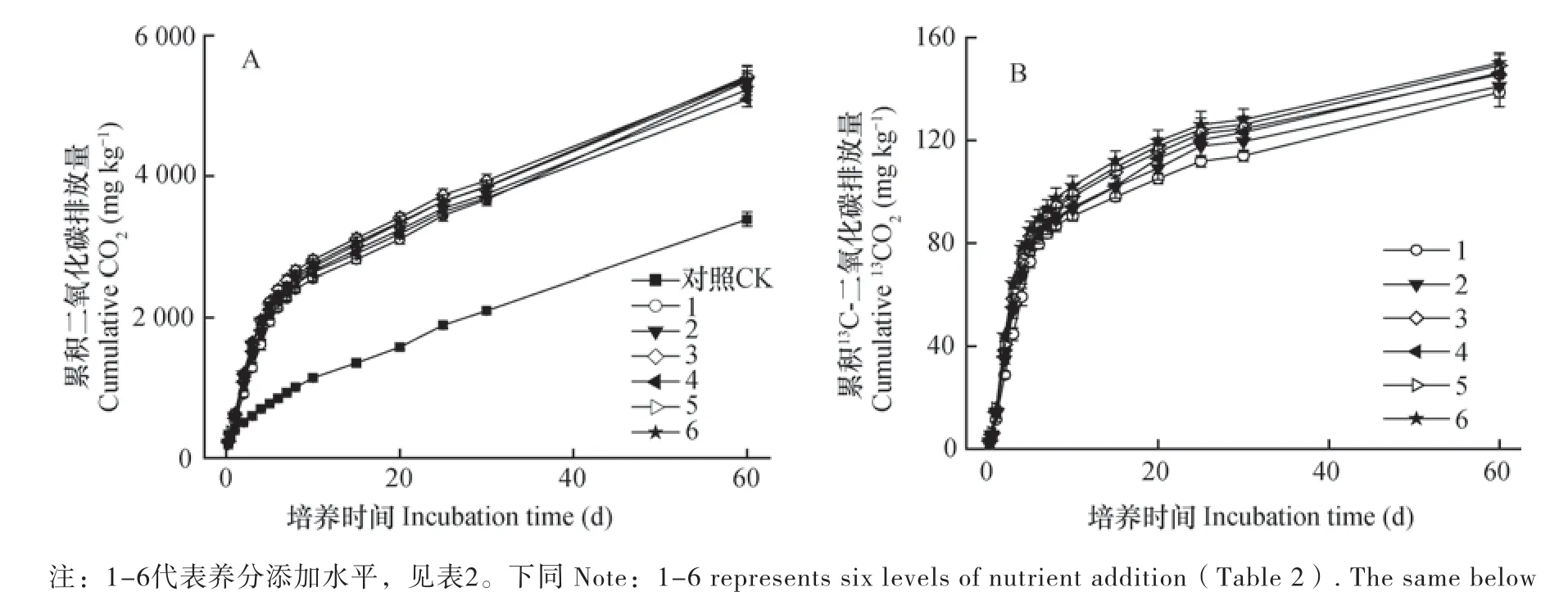
图1 不同元素计量比条件下葡萄糖矿化产生CO2(A)和13CO2(B)规律Fig. 1 Cumulative CO2(A)and13CO2(B)production from mineralization of glucose relative to stoichiometric ratio of nutrient elements
土壤中添加的葡萄糖快速分解,60 d培养实验结束时,有65.5%~74.6%的葡萄糖-C矿化,养分元素的加入提高了葡萄糖的矿化速率。相比处理1(不加NPS等养分元素),养分元素的加入使葡萄糖-C的矿化率提高了3.9%~12.5%(图2A)。同时,不同C/N/P/S计量比条件下,即随着C与养分元素的比值降低,葡萄糖-C矿化率逐渐增加,60 d培养实验后,葡萄糖-C的累积矿化率与养分元素添加量之间表现出显著的正相关关系(R2=0.83,p<0.05)(图2B)。
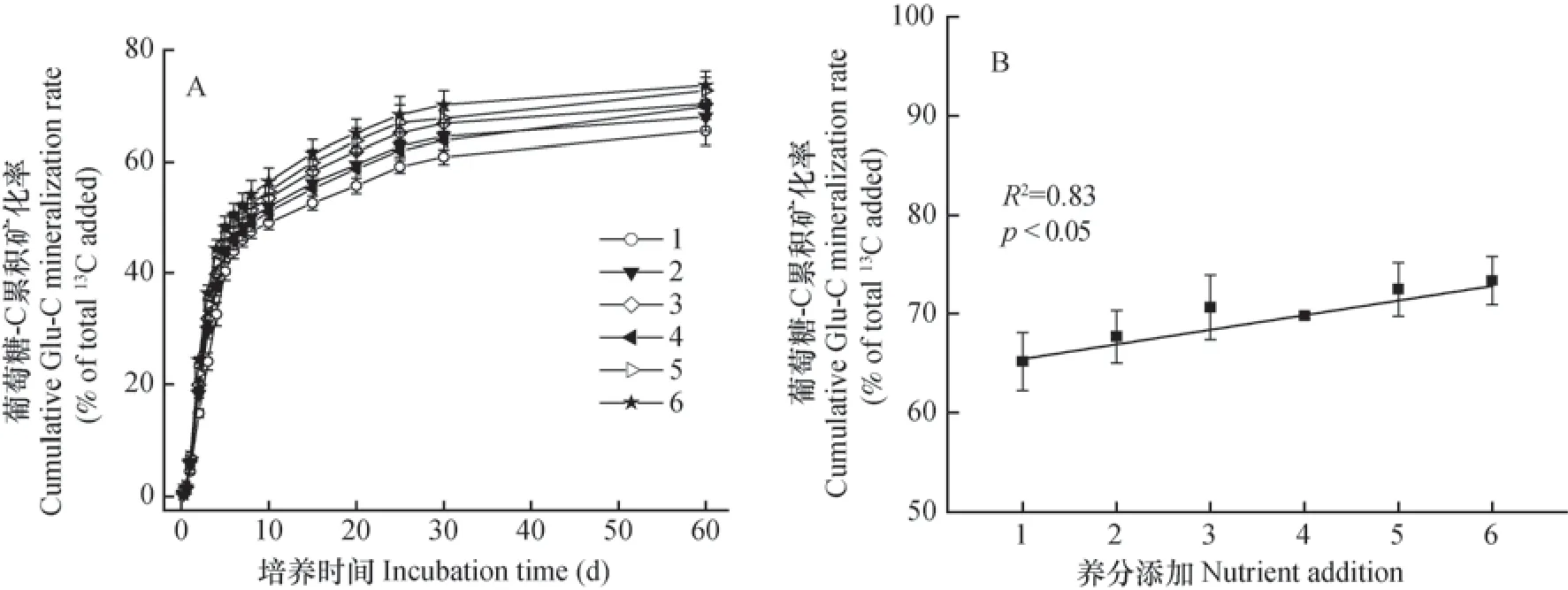
图2 不同元素计量比条件下的葡萄糖-C矿化率(A)和葡萄糖60天累积矿化率与元素计量比之间的关系(B)Fig. 2 Cumulative glucose mineralization rate at the end of 60 days incubation(A)and relationships of stoichiometric ratio of nutrient elements with cumulative glucose-C mineralization rate(B)
2.2 不同元素计量比条件下土壤中葡萄糖碳库的分配
养分元素的添加不仅改变葡萄糖的矿化率,而且还通过增加葡萄糖-C向微生物快速转化碳库的分配比例,加快葡萄糖的转化,从而改变葡萄糖的分解动态。利用一级动力学双指数模型拟合葡萄糖-C矿化率的动态变化[7],由图3可知,随着养分元素添加量的增加,C与养分元素的比例降低,土壤中葡萄糖-C能够被微生物快速代谢的部分(称为葡萄糖-C快库,a1)分配的比例逐渐由58%增加至65%;与之相应的,葡萄糖-C被暂时固定于微生物体内以及被土壤矿物或团聚体稳定的部分,该部分葡萄糖-C的周转速率较慢,称为葡萄糖-C慢库(a2)[7],这部分的比例则由42%逐渐降低至35%,土壤中葡萄糖-C快库、慢库的比例与养分元素添加梯度之间均表现出显著的线性关系(R2= 0.63,p<0.05和R2= 0.61,p<0.05)。

图3 不同元素计量比条件下土壤中葡萄糖-C快库(a1)与慢库(a2)的比例及其与元素计量比之间的关系Fig. 3 Relationships of stoichiometric ratio of nutrient elements with the ratio of readily available C(a1)and slowly available C (a2)in13C glucose
2.3 不同元素计量比条件下的激发效应
土壤中葡萄糖分解过程中,其累积激发效应总体上表现为负激发效应,且随时间负激发效应越来越强,累积负激发效应为-370~-570 mg kg-1(图4A)。在不加养分元素条件下(处理1),葡萄糖矿化的初始阶段(前10天)表现出正激发效应,此后表现为负激发效应,相对养分元素添加条件下,其正激发的时间持续更久;添加养分元素后(处理2~6),葡萄糖-C矿化过程中均表现出负激发效应,而且养分元素的添加比例越大,其负激发效应越强。在60 d的葡萄糖培养实验结束时,累积激发效应与养分元素计量比之间呈现显著的负相关关系(R2= 0.66,p<0.05)(图4B)。
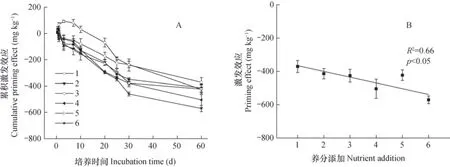
图4 不同元素计量比条件下的累积激发效应(A)及60天时累积激发效应与元素计量比的关系(B)Fig. 4 Cumulative priming effect of initial 60 days(A)and relationships of stoichiometric ratio of nutrient elements with cumulative priming effect(B)
3 讨 论
3.1 元素计量比对土壤外源碳矿化的影响
C、N、P等元素对生物个体、生态系统结构与功能均具有重要作用,而且C、N、P等元素的循环过程相互耦合[18,21],土壤C的生物地球化学循环取决于其和N、P等养分元素间的平衡关系。基于生态化学计量学原理,土壤C周转取决于N和P对微生物的影响[17,21]。土壤中的N、P是生态系统中最常见的限制性元素,是土壤有机碳矿化的主要限制因素[22-23]。美国明尼苏达州沼泽湿地的研究结果显示,施用N、P 肥料增加了土壤碳矿化[24]。Fisk等[25]通过培养实验表明,随着外源 P 输入量的增加,土壤有机碳的矿化速率和累积矿化量均增大。本研究也发现,在不同C/N/P/S计量比条件下,随着N、P、S添加量的增加,稻田土壤碳的矿化量逐渐增加,葡萄糖-C矿化率逐渐增加,且葡萄糖-C的累积矿化率与养分元素添加量之间表现出显著的相关性。表明在土壤碳含量一定的条件下,随着养分元素供应量的增加,微生物为了满足自身生长对碳源和能源的需求而不断提高对碳的利用转化[17,21];同时葡萄糖-C的累积矿化率与土壤中添加的元素计量比呈现出显著的相关性(R2= 0.83,p<0.05),表明养分元素的添加促进了微生物对碳源的利用,元素计量比调控葡萄糖的矿化。
土壤活性有机碳库通过调节土壤能源物质和微生物活性影响着土壤碳库的有效性,与土壤生态系统中有机碳的迁移、固持以及CO2、CH4的释放有密切联系,成为土壤有机碳分解矿化的动力和土壤质量演变的关键指标[26]。基于葡萄糖-C矿化率的一级双指数模型,得到土壤中葡萄糖-C主要是以微生物易利用态的快速转化碳库(a1)形式存在,其比例为60%左右;养分元素的添加改变了葡萄糖-C碳库的分配比例,随着N、P等养分元素的增加,葡萄糖-C向快速转化碳库分配的比例逐渐增加,慢速转化碳库的比例逐渐降低,且两种碳库的分配比例均与养分元素添加量表现出显著的线性关系(R2= 0.63,p<0.05和R2= 0.61,p<0.05),这表明,葡萄糖-C作为易利用态碳源在土壤碳周转过程中主要以快库形式存在。此外,在不同养分条件下土壤微生物群落结构可能不同[6,8],受元素计量比调控,在低养分元素添加条件下,微生物利用C的能力较弱,使得外源的葡萄糖-C转化速率降低,慢速转化的比例增加,反之,高养分元素添加条件下,微生物对碳的需求增加,会优先利用葡萄糖,满足微生物生长需要的元素计量比,使得葡萄糖-C快速转化部分的比例增加。
3.2 葡萄糖激发效应的计量学调控机制
土壤中新输入的有机碳能促进或阻碍土壤有机碳的矿化,引起正的或负的激发效应[20]。有机质的矿化分解主要在土壤微生物的作用下进行,当新鲜有机质添加到土壤中,土壤微生物种群结构立即发生变化,根据土壤微生物对不同有机物质的分解能力可将它们分为受C源限制和受N源限制的两大类微生物[6-7]。由于本研究中用的外源C是葡萄糖,所以在培养实验的初始阶段N、P等养分元素是微生物的主要限制元素。当只加入葡萄糖而不添加养分元素时(处理1),在大量葡萄糖-C的刺激下,微生物需要有足够的N、P、S等养分元素满足其生长对养分元素的计量比需求;而且,本实验用的土壤是采自长期定位试验田的低磷水稻土(有效磷含量为2.7 mg kg-1(表1)),处于养分元素受限条件,因此,在培养实验初期(0~10 d),微生物需要通过分解土壤有机质获取N、P等养分元素,所以产生正激发效应;而在后期(10~60 d),随着外源碳的消耗、土壤中溶出的N、P等养分元素可以满足微生物需求,微生物不再处于养分受限环境,减弱对土壤原有有机质的分解,因而产生负激发效应。
在土壤中添加葡萄糖,并同时按照元素计量比添加养分元素时(处理2~6),在培养实验最初的1~2 d内,表现为正激发效应,这可能是因为在培养试验的初始阶段,外源养分元素的输入,刺激了处于养分元素受限条件下的土壤微生物,使其在利用外源葡萄糖-C的同时也加快了对土壤原有有机质的分解[6];在短暂的正激发后,各养分元素添加水平下,均表现出了负激发效应,这可能是因为土壤中原有的或是外源添加的N、P、S养分元素能够满足微生物对养分元素的需求,而且葡萄糖-C也满足微生物需要的C量,使得微生物减弱对土壤原有有机质的分解,产生了负激发效应,保护了土壤原有有机碳[27]。
不同C/N/P/S计量比条件下,60 d时的累积激发效应与元素计量比呈现显著的线性关系(R2= 0.66,p<0.05),表明一定量的N、P、S养分元素供应使微生物对养分元素的需求减弱,降低土壤原有有机质的分解。葡萄糖-C的添加促使土壤微生物优先利用该易利用态碳源以维持生长,养分元素越多负激发越强,可能因为较多的养分元素更易于满足微生物生长的需求,从而减弱对土壤中原有养分元素的依赖,降低土壤原有有机质的分解,进而增加土壤有机质的积累[13]。
4 结 论
稻田土壤中外源碳的矿化受养分元素的影响,高比例的N、P、S养分元素的添加,使土壤有机碳向易利用态碳库分配,进而促进土壤外源有机碳的矿化,增加土壤碳的负激发效应。在6个C/N/P/S元素计量比条件下,土壤碳的转化与养分元素添加水平表现出显著的线性关系,说明稻田土壤有机碳的转化受元素计量比调控。然而,本研究只选择了葡萄糖作为典型碳源,探讨了在6个养分元素梯度下矿化的计量学调控特征,而对于外源碳在土壤中的转化及其生态酶学与分子生物学机制方面的研究还需加强,从而能够在广度和深度上更好地阐释生态计量学的调控机制。
参 考 文 献
[1]Lal R. Soil carbon sequestration impacts on global climate change and food security. Science,2004,304 (5677):1623—1627
[2]Pan G,Li L,Wu L,et al. Storage and sequestration potential of topsoil organic carbon in China’s paddy soils. Global Change Biology,2004,10(1):79—92
[3]张艳鸿,窦森,董珊珊,等. 秸秆深还及配施化肥对土壤腐殖质组成和胡敏酸结构的影响. 土壤学报,2016,53(3):694—702
Zhang Y H,Dou S,Dong S S,et al. Effect of deep incorporation of corn stover combined chemical fertilizer on composition of soil humus and structure of humic acid in soil(In Chinese). Acta Pedologica Sinica,2016,53(3):694—702
[4]Pump J,Conrad R. Rice biomass production and carbon cycling in13CO2pulse-labeled microcosms with different soils under submerged conditions. Plant and Soil,2014,384(1):213—229
[5]Tian J,Dippold M,Pausch J,et al. Microbial response to rhizodeposition depending on water regimes in paddy soils. Soil Biology & Biochemistry,2013,65:195—203
[6]Chen R,Senbayram M,Blagodatsky S,et al. Soil C and N availability determine the priming effect:Microbial N mining and stoichiometric decomposition theories. Global Change Biology,2014,20(7):2356—2367
[7]Creamer C A,Jones D L,Baldock J A,et al. Stoichiometric controls upon low molecular weight carbon decomposition. Soil Biology & Biochemistry,2014,79:50—56
[8]Sinsabaugh R L,Manzoni S,Moorhead D L,et al. Carbon use efficiency of microbial communities:Stoichiometry,methodology and modeling. Ecology Letters,2013,16(7):930—939
[9]吴金水,葛体达,祝贞科. 稻田土壤碳循环关键微生物过程的计量学调控机制探讨. 地球科学进展,2015,30(9):1006—1017
Wu J S,Ge T D,Zhu Z K. Discussion on the key microbial process of carbon cycle and stoichiometric regulation mechanisms in paddy soils(In Chinese). Advances in Earth Science,2015,30(9):1006—1017
[10]Haynes R J. Labile organic matter as an indicator of organic matter quality in arable and pastoral soils in New Zealand. Soil Biology & Biochemistry,2000,32 (2):211—219
[11]Sinsabaugh R L,Follstad Shah J J. Ecoenzymatic stoichiometry and ecological theory. Annual Review of Ecology,Evolution,and Systematics,2012,43 (1):313—343
[12]Spohn M,Chodak M. Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils. Soil Biology & Biochemistry,2015,81:128—133
[13]Kirkby C A,Richardson A E,Wade L J,et al. Carbon-nutrient stoichiometry to increase soil carbon sequestration. Soil Biology & Biochemistry,2013,60:77—86
[14]Ross D J,Speir T W,Kettles H A,et al. Soil microbial biomass,C and N mineralization and enzyme activities in a hill pasture:Influence of season and slow-release P and S fertilizer. Soil Biology & Biochemistry,1995,27(11):1431—1443
[15]夏昕,石坤,黄欠如,等. 长期不同施肥条件下红壤性水稻土微生物群落结构的变化. 土壤学报,2015,52 (3):697—705
Xia X,Shi K,Huang Q R,et al. The changes of microbial community structure in red paddy soil under long-term fertilization(In Chinese). Acta Pedologica Sinica,2015,52(3):697—705
[16]王绍强,于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报,2008,28(8):3937—3947
Wang S Q,Yu G R. Ecological stoichicmetry characteristics of ecosystem carbon nitrogen and phosphorus elements(In Chinese). Acta Ecologica Sinica,2008,28(8):3937—3947
[17]Alberti G,Vicca S,Inglima I,et al. Soil C:N stoichiometry controls carbon sink partitioning between above-ground tree biomass and soil organic matter in high fertility forests. iForest-Biogeosciences and Forestry,2015,8:195—206
[18]Li Y,Wu J,Liu S,et al. Is the C∶N∶P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles,2012,26 (4):GB4002
[19]Phillips D,Newsome S,Gregg J. Combining sources in stable isotope mixing models:Alternative methods. Oecologia,2005,144(4):520—527
[20]Kuzyakov Y. Priming effects:Interactions between living and dead organic matter. Soil Biology & Biochemistry,2010,42(9):1363—1371
[21]Heuck C,Weig A,Spohn M. Soil microbial biomass C∶N∶P stoichiometry and microbial use of organic phosphorus. Soil Biology & Biochemistry,2015,85:119—129
[22]Liu D Y,Song C C. Effects of phosphorus enrichment on mineralization of organic carbon and contents of dissolved carbon in a freshwater marsh soil. China Environmental Science,2008,28(9):769—774
[23]Spohn M,Kuzyakov Y. Phosphorus mineralization can be driven by microbial need for carbon. Soil Biology & Biochemistry,2013,61:69—75
[24]Keller J K,Bridgham S D,Chapin C T,et al. Limited effects of six years of fertilization on carbon mineralization dynamics in a Minnesota fen. Soil Biology & Biochemistry,2005,37(6):1197—1204
[25]Fisk M,Santangelo S,Minick K. Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests. Soil Biology & Biochemistry,2015,81:212—218
[26]Dharmakeerthi R S,Hanley K,Whitman T,et al. Organic carbon dynamics in soils with pyrogenic organic matter that received plant residue additions over seven years. Soil Biology & Biochemistry,2015,88:268—274
[27]Kirkby C A,Richardson A E,Wade L J,et al. Nutrient availability limits carbon sequestration in arable soils. Soil Biology & Biochemistry,2014,68:402—409
Effect of Stoichiometric Ratio of Soil Nutrients on Mineralization and Priming Effect of Glucose in Paddy Soil
TANG Zhenzhu1,2ZHU Zhenke2†SHEN Bingjie2,3HU Yajun2WANG Juan2PANG Jing1†
TONG Chengli2GE Tida2WU Jinshui2
(1 School of Resources and Environmental Science,Hubei University,Wuhan 430062,China)
(2 Key Laboratory of Agro Ecological Processes in Subtropical Region,Institute of Subtropical Agriculture,Chinese Academy of Sciences,Changsha 410125,China)
(3 College of Environmental Science and Technology,Central South University of Forestry and Technology,Changsha 410004,China)
【Objective】The turnover of liable organic carbon(C)sources in soil is a key component of the cycle of soil carbon. Liable C sources(i.e. glucose),derived from C deposition in rhizosphere and other extraneous C inputs,are important sources of the C pool in the soil. Therefore,studying the turnover of liable C sources as affected by stoichiometric ratio of soil nutrients may help reveal mechanisms of the eco-stoichiometric regulation of the turnover of extraneous C in paddy soil. 【Method】In this study,an inlab incubation experiment was conducted using13C-glucose as extraneous organic C typical in paddy soil to explore quantitatively dynamics of decompositing mineralization and priming effect of glucose as affected by C/ N/P/S stoichiometric ratio. 【Result】Results show that glucose was rapidly mineralized in the paddy soil so that 65.5% to 74.6% of the added glucose-C was mineralized after 60 d of incubation. The addition of nutrient elements not only increased glucose mineralization rate,but also raised the proportion of glucose-C available to soil microbes from 58% to 65%,thus accelerating glucose turnover rate in the soil. The amount of nutrient elements added was found significantly and positively related to both proportion of the microbe-available C pool in and mineralization rate(R2= 0.63,p <0.05;R2= 0.83,p<0.05)of glucose C,which suggests that the addition of nutrient elements promoted the microbial utilization of extraneous C in the soil,and stoichiometric ratio of the nutrient elements regulated the dynamic of glucose mineralization. With the addition of glucose-C,the decomposition of the soil native organic matter was inhibited,and the cumulative negative priming effect was -370 to -570 mg kg-1. The higher the amount of nutrient elements added,the stronger the negative priming effect observed,and the two were obviously in negative correlation(R2= 0.66,p<0.05),indicating that the soil microbes preferentially utilize the added glucose-C to sustain their growth. The supply of N,P and S reduced the dependence of microbes on nutrient elements in the soil native organic matter,and thereby slow down the decomposition of soil organic matter. 【Conclusion】Mineralization of liable C in paddy soil is affected by C/N/P/S stoichiometric ratio. The addition of a high ratio of nutrient elements promotes mineralization of liable C in paddy soil,inhibits decomposition of native soil organic matter,and intensify negative priming effect. All the findings in this study demonstrate that application of liable C and fertilizers to paddy soils could effectively inhibit decomposition of soil native organic matter,and stimulate accumulation of soil organic matter,thus building up soil fertility.
Paddy soil;Organic carbon mineralization;Priming effect;Nutrient stoichiometry
S15
A
10.11766/trxb201604260070
(责任编辑:陈荣府)
* 国家自然科学基金项目(41430860,41371304)、湖南省自然科学基金项目(2016JJ3132)和中国科学院战略性先导科技专项(XDB15020401)资助 Supported by the National Natural Science Foundation of China(Nos. 41430860 and 41371304),Hunan Provincial Natural Science Foundation of China(No. 2016JJ3132)and the Strategic Priority Research Program of the Chinese Academy of Sciences(No. XDB15020401)
† 通讯作者 Corresponding authors,E-mail:zhuzhenke@isa.ac.cn;pangjing2286@126.com
汤珍珠(1992―),女,江苏常州人,硕士研究生,主要研究土壤地理学。E-mail:1486430455@qq.com
6;
2016-08-06;优先数字出版日期(www.cnki.net):2016-09-21
猜你喜欢
建材发展导向(2022年24期)2022-12-22 07:44:36
舰船科学技术(2022年11期)2022-07-15 07:51:56
环境卫生工程(2021年4期)2021-10-13 06:52:26
绿色科技(2020年20期)2020-11-20 01:56:34
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
江苏农业科学(2017年6期)2017-05-11 14:35:37
上海农业学报(2017年3期)2017-04-10 12:39:12
林业勘查设计(2017年1期)2017-03-11 16:49:57
丝绸(2014年5期)2014-02-28 14:55:12
