
高寒草甸土壤微生物群落功能多样性对广布弓背蚁蚁丘扰动的响应
2017-02-15 08:21:33杨有芳字洪标刘敏阿的鲁骥陈焱王长庭
草业学报 2017年1期
杨有芳,字洪标,刘敏,阿的鲁骥,陈焱,王长庭
(西南民族大学生命科学与技术学院,四川 成都610041)
高寒草甸土壤微生物群落功能多样性对广布弓背蚁蚁丘扰动的响应
杨有芳1,字洪标1,刘敏1,阿的鲁骥1,陈焱1,王长庭*
(西南民族大学生命科学与技术学院,四川 成都610041)
为了解蚂蚁在高寒草甸生态系统中的功能和作用,评估和认识高寒草甸生态系统的健康状况和稳定性,利用Biolog-ECO生态板法对蚁丘干扰后高寒草甸土壤理化性质以及土壤微生物功能多样性进行研究分析,结果表明,1)蚁丘扰动后0~10 cm土层土壤有机质、全氮、速效氮、速效磷、速效钾含量均显著高于蚁丘10~20 cm土层(P<0.05),而蚁丘扰动与土层深度的交互作用对理化性质均无显著影响(P>0.05);2)蚁丘干扰后高寒草甸土壤微生物多样性指数(Shannon-Weiner指数、Pielou指数、McIntosh指数和底物碳源利用数)有所增加;3)主成分分析表明蚁丘干扰对土壤微生物功能多样性产生了影响,改变了微生物代谢功能特征,土壤微生物利用的主要碳源是碳水化合物、氨基酸类和羧酸类;4)冗余分析表明土壤养分影响土壤微生物碳源代谢特征和功能。因此,由于蚂蚁的挖掘活动、有机质的累积等改变了蚁丘土壤微环境,进而改变了土壤微生物的碳源利用类型和代谢功能。
蚁丘;Biolog-ECO;土壤微生物群落;高寒草甸
高寒草甸是青藏高原的代表性草地类型,该类型的草甸虽然草层低矮,但分布面积广,草质柔软,营养丰富,适口性好,比较耐牧,是牦牛和藏羊放牧的优良场地[1]。近几十年,因为气候环境的改变以及人为不合理的利用行为,导致高寒草甸生态系统的功能和稳定性逐渐下降。气候变化降低了牧草开花期和籽粒成熟期的发育百分率,影响了牧草的产籽量和次年牧草的返青[2],气候变化通过光、热、水等要素变化影响土壤有机质、土壤微生物的活动和繁殖,从而影响土壤肥力[3]。而持续地超载过牧不但降低草地生态系统的初级生产力,而且明显改变草地植物群落的组成和结构[4],家畜等大型食草动物通过则改变草地地上和地下凋落物的数量和品质,以及排放尿液和粪便来间接影响植被和生态系统特征[5],适度的放牧干扰有利于地表凋落物的分解和输入,增加土壤养分含量,有利于土壤微生物的繁殖[6]。
干扰是自然界无处不在的一种普遍现象,一定程度的干扰可以使生物多样性、群落结构的分化与复杂性及生态位的增加[7-8]。随着人为活动的加剧,干扰越来越受到人们的关注,干扰的生态学作用也在近年来受到广泛的重视[9]。蚂蚁是陆地生态系统中分布最广的生物之一,其活动是草地生态系统中不可忽视的生物干扰因子[10]。它们不仅是土壤动物生物量的主要组成部分,而且通过生物扰动和刺激土壤有机质转化影响土壤养分循环[11]。蚂蚁在筑巢过程中收集大量木质碎屑、昆虫猎物和蜜糖类物质作为食物。因此,活跃的蚁丘比相邻的非蚁丘表层土壤富含有机质和无机营养元素[12]。蚂蚁的活动也可以改变土壤的理化性质、土壤湿度、土壤有机质及N、P、K等各元素的含量[13-14],改变土壤微生物群落结构和其他动物的生物量[15],影响有机质的分解率[16]。国内外关于蚂蚁群落物种多样性、蚂蚁的行为活动与植物的关系等的研究均有报道[8,10,17]。然而,蚂蚁活动形成的蚁丘对高寒草甸土壤微生物群落组成和功能多样性的影响以及土壤微生物群落与地上植被的关系、反馈机制等方面的研究鲜有报道。
土壤微生物通过分解枯枝落叶以及动物残体等参与高寒草甸生态系统的物质循环和能量流动,从而影响地上植被的生长发育,是碎屑养分系统的基本组成成分[18],也是土壤系统的主要组分、判断土壤肥力的重要指标之一。土壤微生物功能多样性可以反映土壤中微生物的生态特征,通过对土壤微生物群落功能多样性的研究,可以反映土壤有效肥力的供应情况,并能够反映某个地区土壤退化或恢复的程度[19]。本研究通过调查川西北高寒草甸上的蚁丘,利用Biolog-ECO板测定蚁丘土壤微生物代谢活动和功能多样性,分析蚁丘对土壤理化性质、土壤微生物功能多样性的影响,为正确评价蚂蚁在高寒草甸生态系统特别是土壤生态系统中的功能和作用提供理论依据。
1 材料与方法
1.1 研究区自然概况
本试验的研究区位于青藏高原东缘四川省阿坝州红原县境内(31°50′-33°22′ N,101°51′-103°23′ E),地势由东南向西北倾斜,平均海拔3500 m,草地面积6293 km2,占全县总面积的76.8%,大部分属于高寒草甸类型;属大陆高原寒温带半湿润季风气候,年降水量650~800 mm,80%集中在5-8月份。年平均气温1.1 ℃,最高温度24.6 ℃,最低温度-10.3 ℃,年平均积温1432.3 ℃,年平均日照2417.9 h。植被平均盖度超过80%,植被高度可达45~60 cm,植被以莎草科和禾本科为主,如四川嵩草(Kobresiasetschwanensis)和高山嵩草(Kobresiapygmaea),四川剪股颖(Agrostisclavata)和垂穗披碱草(Elymusnutans),杂类草中条叶银莲花(Anemonetrullifolia)、钝苞雪莲(Saussureanigrescens)和委陵菜属(Potentillaspp.)比较常见[20]。试验区土壤为亚高寒草甸土,土壤速效氮含量较高,有机质丰富,速效钾含量一般丰富,速效磷含量较少[8],pH值为4.6左右[21]。
1.2 研究方法
1.2.1 实验设计和样品采集 本研究于2011年8月在四川省红原县西南民族大学青藏高原生态保护与畜牧业高科技研发基地试验园区进行。广布弓背蚁(Camponotusherculeanus)是实验样地中最为常见的蚂蚁,喜在落叶和枯枝下营巢[22],在四川省红原县高寒草甸分布较多。设置2个平行样地,选取广布弓背蚁筑巢自然形成的直径大小为30~50 cm,高度为10~20 cm的蚁丘各3个,面积约为0.5 m×0.5 m,共计6个蚁丘,分别编号为Ant-hill 1#、Ant-hill 2#、Ant-hill 3#、Ant-hill 4#、Ant-hill 5#、Ant-hill 6#;与蚁丘样地相距4~5 m处且没有蚁丘扰动的草地作为对照样地,随机选取6个对照样方(0.5 m×0.5 m),分别编号为CK 1#、CK 2#、CK 3#、CK 4#、CK 5#、CK 6#。
在每个测定过地上生物量的样方内,用土钻(直径3 cm)采用“V”字形法钻取5钻,按0~10 cm、10~20 cm 土层分别取样,做好标记,其中一部分过0.25 mm土壤筛,风干后用于测定土壤pH值和养分;另一部分拣去石块和杂物等做好标记,迅速放入保鲜盒,带回实验室放入4 ℃冰箱保存,用于土壤微生物功能多样性的测定。蚁丘干扰试验区(ant-hill disturbance experiment)概况如表1所示。

表1 试验区概况Table 1 General properties of the study sites
1.2.2 土壤pH和土壤养分测定 土壤pH采用电极法测定,土壤有机质(soil organic matter, SOM)采用重铬酸钾容量法(外加热法)测定,土壤全氮(total nitrogen, TN)采用凯氏定氮法测定,土壤速效氮(available nitrogen, AN)采用扩散皿法测定[23],土壤全磷(total phosphorus, TP)和土壤速效磷 (available phosphorus, AP)分别采用钼锑抗比色法和碳酸氢钠浸提-钼锑抗比色法测定,土壤全钾(total potassium, TK)和土壤速效钾(available potassium, AK)含量分别采用醋酸铵法和四苯硼钠法测定[24]。
1.2.3 土壤微生物多样性的测定和计算 土壤微生物功能多样性的测定采用Biolog-ECO生态板,每个0~10 cm、10~20 cm土壤样品进行3次重复测定。具体操作方法参照Chaerun等[25]、Biolog生态板每孔的平均颜色变化率(average well color development, AWCD)的计算采用Classen等[26]的方法。AWCD值是一个用于评价土壤微生物对碳源的利用程度,指示土壤微生物的代谢活性的重要指标。Shannon-Weiner指数(H)是反映群落物种及其个体分布均匀程度的综合指标,受群落物种丰富度影响较大,能较全面地测度物种的多样性(公式2);Pielou指数(J)是群落物种丰富度均一性的度量(公式3);McIntosh指数(U)则是反映土壤中微生物群落均匀度的指标(公式4)。各指标计算公式如下:
(1)
(2)
(3)
(4)
式中:Ci为第i孔的590和750 nm下的吸光值之差;R为对照孔的吸光值;n为Biolog微平板的碳源种类数目,n=31;Pi为第i孔的相对吸光度值与整个平板相对吸光度值总和的比率;S为产生颜色变化的孔的数目;ni为第i孔的相对吸光度值[27]。
1.3 数据分析
采用SPSS 19.0对微生物群落多样性指数、底物碳源利用数进行单因素方差分析(one-way ANOVA),对土壤理化数据进行双因素方差分析(two-way ANOVA),最小显著差数法(LSD法)进行多重比较。用Pearson相关系数评价AWCD值、微生物物种多样性指数、6类碳源利用能力与土壤理化性质间的相关关系。用软件CANOCO for Windows 4.5对培养72 h的土壤微生物进行主成分分析 (principal component analysis, PCA)来解释土壤微生物功能特征的变化;使用线性冗余分析(redundancy analysis, RDA)来解释土壤理化性质与土壤微生物功能多样性间的关系(α=0.05)。
2 结果与分析
2.1 蚁丘扰动对高寒草甸土壤理化性质的影响
方差分析表明,蚁丘扰动对TN、TK、AP和AK具有显著影响(P<0.05);不同土层深度对SOM、TN、AN、AP、AK有显著影响(P<0.05);蚁丘扰动与土层深度的交互作用对土壤理化性质均无显著影响(P>0.05)(表2)。SOM、TN、AN、AP、AK在蚁丘扰动土壤中和对照土壤中的含量均表现出0~10 cm>10~20 cm(P<0.05),蚁丘扰动土壤中的SOM、TN、AN、AP和AK含量相对于对照土壤分别增加了30.45 g/kg,1.63 g/kg,89.16 mg/kg,5.49 mg/kg,220 mg/kg,其中,蚁丘扰动与对照间相同土层土壤AK含量以及10~20 cm土层AP含量均表现出蚁丘扰动显著大于对照(P<0.05),对其余理化性质均无显著影响(P>0.05)(图1)。
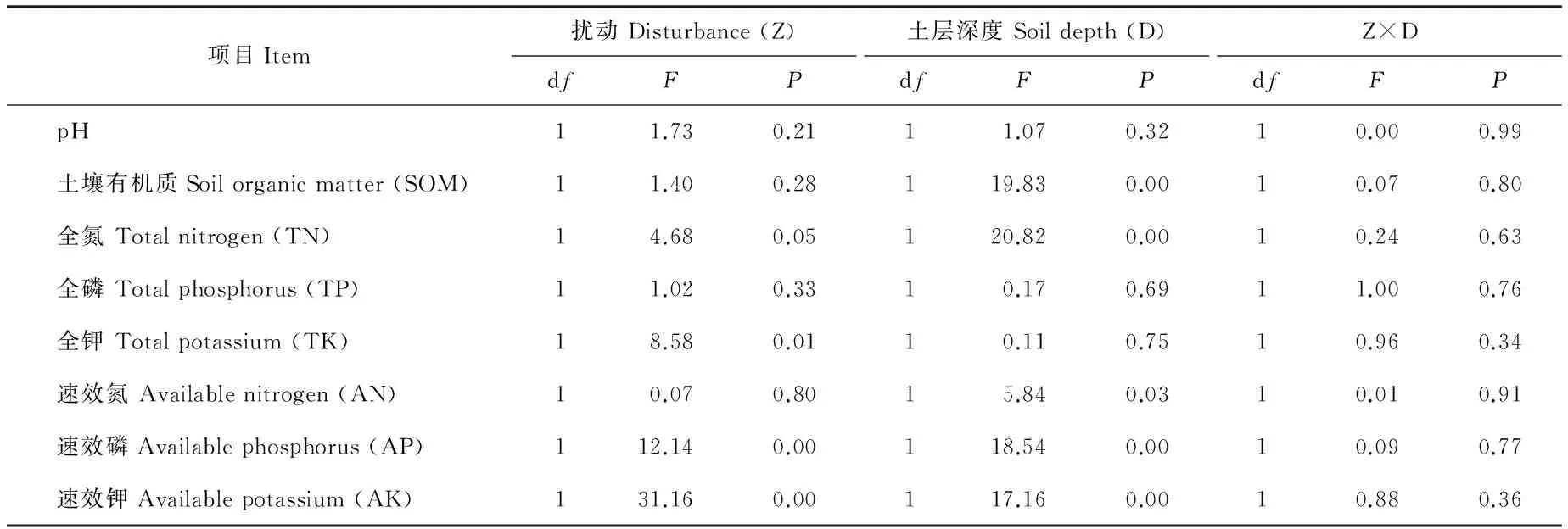
表2 蚁丘扰动不同土壤深度对土壤理化性质的影响的方差分析结果Table 2 Result of two-way analyses of variance showing the effects of ant-hill disturbance and soil depth on soil physicochemical properties (Mean±SE, n=5)
2.2 蚁丘对高寒草甸土壤微生物功能多样性的影响
2.2.1 蚁丘对高寒草甸土壤微生物生长曲线的影响 高寒草甸土壤微生物AWCD值对蚁丘干扰的响应如图2所示,蚁丘干扰后高寒草甸土壤微生物的AWCD值显著增加(P<0.05),各土壤样品从开始到培养24 h,AWCD值为0,表明在24 h内碳源基本未被利用;在24~168 h内,AWCD值近似指数型的方式急剧升高,底物碳源被大幅度消耗利用;168~240 h时间段,AWCD 值缓慢增长,微生物生长进入稳定期,土壤微生物底物利用率降低。
2.2.2 土壤微生物多样性指标 从表3可知,蚁丘干扰后高寒草甸土壤微生物Shannon-Weiner指数、Pielou指数、McIntosh指数和碳源利用数显著高于对照土壤(P<0.05),分别比对照增加了0.06,0.03,0.51和0.67。说明蚁丘对高寒草甸土壤微生物群落的代谢活性有促进作用,有利于其代谢活性的提高。
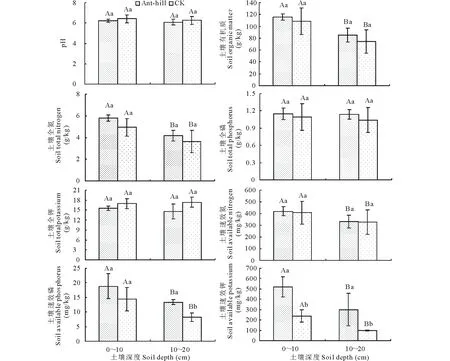
图1 蚁丘扰动对高寒草甸土壤养分的影响Fig.1 Effects of ant-hill on soil nutrients in alpine meadow不同小写字母表示相同土层土壤理化性质显著性差异(P<0.05),不同大写字母表示不同土层土壤理化性质显著性差异(P<0.05)。The different lowercase letters indicated that significant difference of soil physicochemical properties at same soil depth at P<0.05,while different uppercase letters indicated that significant difference of soil physicochemical properties at different soil depth at P<0.05.
2.3 土壤微生物对碳源的利用
用主成分分析方法(PCA)对土壤微生物群落利用碳源情况进行分析,进而了解土壤微生物功能的综合差异。将72 h的AWCD值转化为几个综合变量,以更好地反映微生物群落功能特征对蚁丘干扰的响应,结果如图3所示。前2个主成分的数据变异贡献率为61.4%,其中第1主成分(PC1)的数据变异贡献率为32.9%,第2主成分(PC2)的数据变异贡献率为28.5%。蚁丘土壤(Ant-hill 1#、Ant-hill 2#、Ant-hill 3#、Ant-hill 4#、 Ant-hill 5#、Ant-hill 6#)和CK土壤(CK 1#、CK 2#、CK 3#、CK 4#、 CK 5#、CK 6#)分别位于坐标图的不同区域,两种土壤很好地区分开来。这表明蚁丘干扰对土壤微生物功能多样性产生了影响,改变了微生物代谢功能特征,从而导致了不同的碳源利用图谱。
通过对主成分的因子载荷分析可知(表4),与PC1显著相关的碳源有15种,其中碳水化合物有5种,分别为 β-甲基-D-葡萄糖苷、D-半乳糖酸-γ-内酯、D-半乳糖醛酸、D-纤维二糖、α-D-乳糖;氨基酸类有L-精氨酸、L-苯基丙氨酸、L-丝氨酸、L-苏氨酸和甘氨酰-L-谷氨酸5种;羧酸类有丙酮酸甲酯和α-丁酮酸2种;多聚类有吐温40和α-丁酮酸2种;胺类只有腐胺1种。与PC2显著相关的碳源有14种,其中碳水化合物有5种,分别是D-木糖、i-赤藓糖醇、D-甘露醇、D-葡萄糖胺酸、α-D-葡萄糖-1-磷酸;氨基酸类有4种,分别是L-天门冬氨酸、L-苏氨酸、L-丝氨酸和甘氨酰-L-谷氨酸;酚酸类有2-羟基苯甲酸和4-羟基苯甲酸;羧酸类、多聚物及酚酸类各1种。这一结果表明土壤微生物利用的主要碳源是碳水化合物、氨基酸类和羧酸类,蚁丘扰动土壤和对照土壤微生物群落在对上述碳源的利用能力上存在差异。
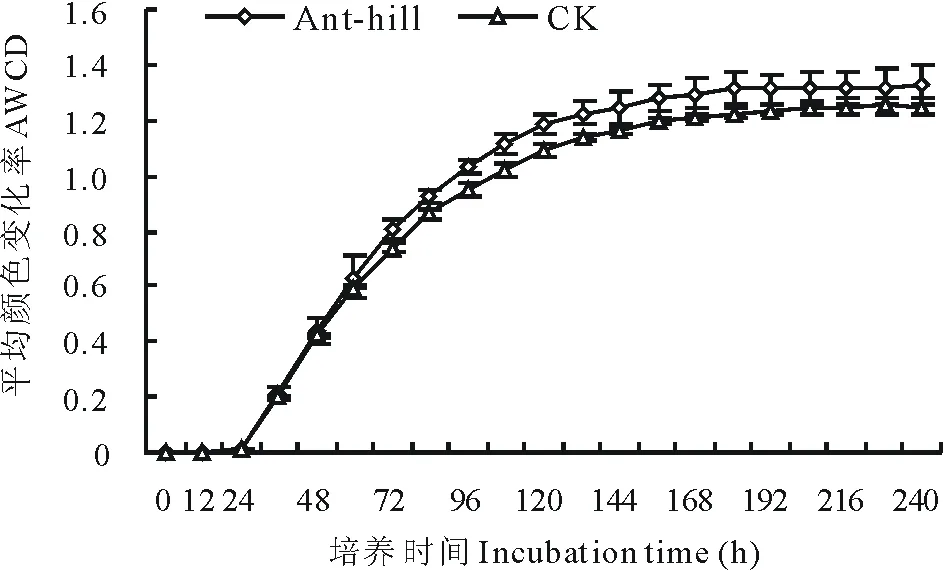
图2 高寒草甸土壤微生物AWCD值对蚁丘的响应Fig.2 Response of AWCD value of alpine meadow soil microbial communities to ant-hill
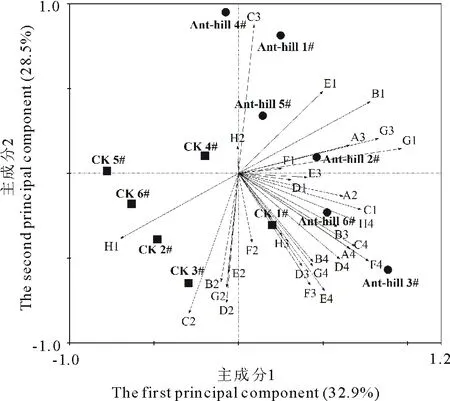
图3 蚁丘试验土壤微生物群落对碳源利用的主成分分析Fig.3 PCA for carbon source utilization of soil microbial communities on the ant-hill experimentA2:β-甲基-D-葡萄糖苷β-methyl-D-glucoside;A3:D-半乳糖酸-γ-内酯D-galactonic acid lactone;A4:L-精氨酸L-arginine;B1:丙酮酸甲酯Pyruvic acid methyl ester;B2:D-木糖D-xylose;B3:D-半乳糖醛酸D-galacturonic acid;B4:L-天门冬酰胺L-asparagine;C1:吐温40 Tween 40;C2:i-赤藓糖醇i-erythritol;C3:2-羟基苯甲酸2-hydroxybenzoic acid;C4:L-苯基丙氨酸L-phenylalanine;D1:吐温80 Tween 80;D2:D-甘露醇D-mannitol;D3:4-羟基苯甲酸4-hydroxybenzoic acid;D4:L-丝氨酸L-serine;E1:α-环式糊精α-cyclodextrin;E2:N-乙酰-D-葡萄糖胺N-acetyl-D-glucosamine;E3:γ-羟基丁酸γ-hydroxybutyric acid;E4:L-苏氨酸L-threonine;F1:肝糖Glycogen;F2:D-葡萄糖胺酸D-glucosaminic acid;F3:衣康酸Itaconic acid;F4:甘氨酰-L-谷氨酸Glycyl-L-glutamic acid;G1:D-纤维二糖D-cellobiose;G2:α-D-葡萄糖-1-磷酸α-D-glucose-1-phosphate;G3:α-丁酮酸α-ketobutyric acid;G4:苯乙胺Phenylethylamine;H1:α-D-乳糖α-D-lactose;H2:D,L-α-磷酸甘油D,L-α-glycerol phosphate;H3:D-苹果酸D-malic acid;H4:腐胺Putrescine.
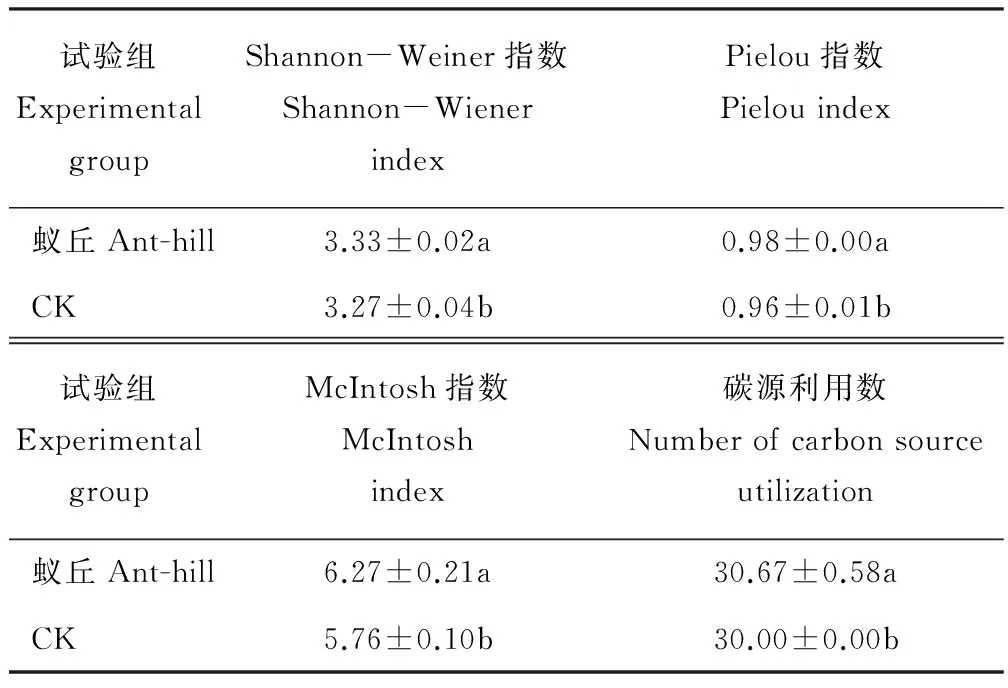
表3 蚁丘对高寒草甸土壤微生物群落功能多样性的影响Table 3 Effect of ant-hill on the alpine meadow soil microbial functional diversity
同列不同字母表示处理间在P<0.05水平上存在显著差异。
Different lowercase letters in the same column indicate significant differences atP<0.05 level.
2.4 蚁丘扰动高寒草甸土壤微生物功能冗余分析
为了探讨环境因子对土壤微生物功能多样性的影响,选择冗余分析(RDA)对环境梯度进行直接排序分析。排序结果表明,第1主成分轴和第2主成分轴分别解释总变异的81.3%和18.6%,前2轴的解释变异达到了99.9% (图4),说明土壤理化性质对土壤微生物碳源代谢特征和功能多样性有较大影响。全钾对第1排序轴有显著负相关关系(P<0.05),速效磷和速效钾与第1排序轴呈极显著正相关(P<0.01);有机质、全氮、全磷和速效氮与第2排序轴存在极显著正相关(P<0.01),而存在显著负相关的为土壤pH(P<0.05)(图4,表5)。
将31种碳源分为碳水化合物(carbohydrate)、氨基酸类(amino acid)、羧酸类(carboxylic acid)、多聚物(polymer)、酚酸类(phenolic acid)和胺(amine)六大类。土壤理化性质对土壤微生物碳源代谢特征具有显著性影响,但对单独6类碳源无显著性影响(表5,表6)。
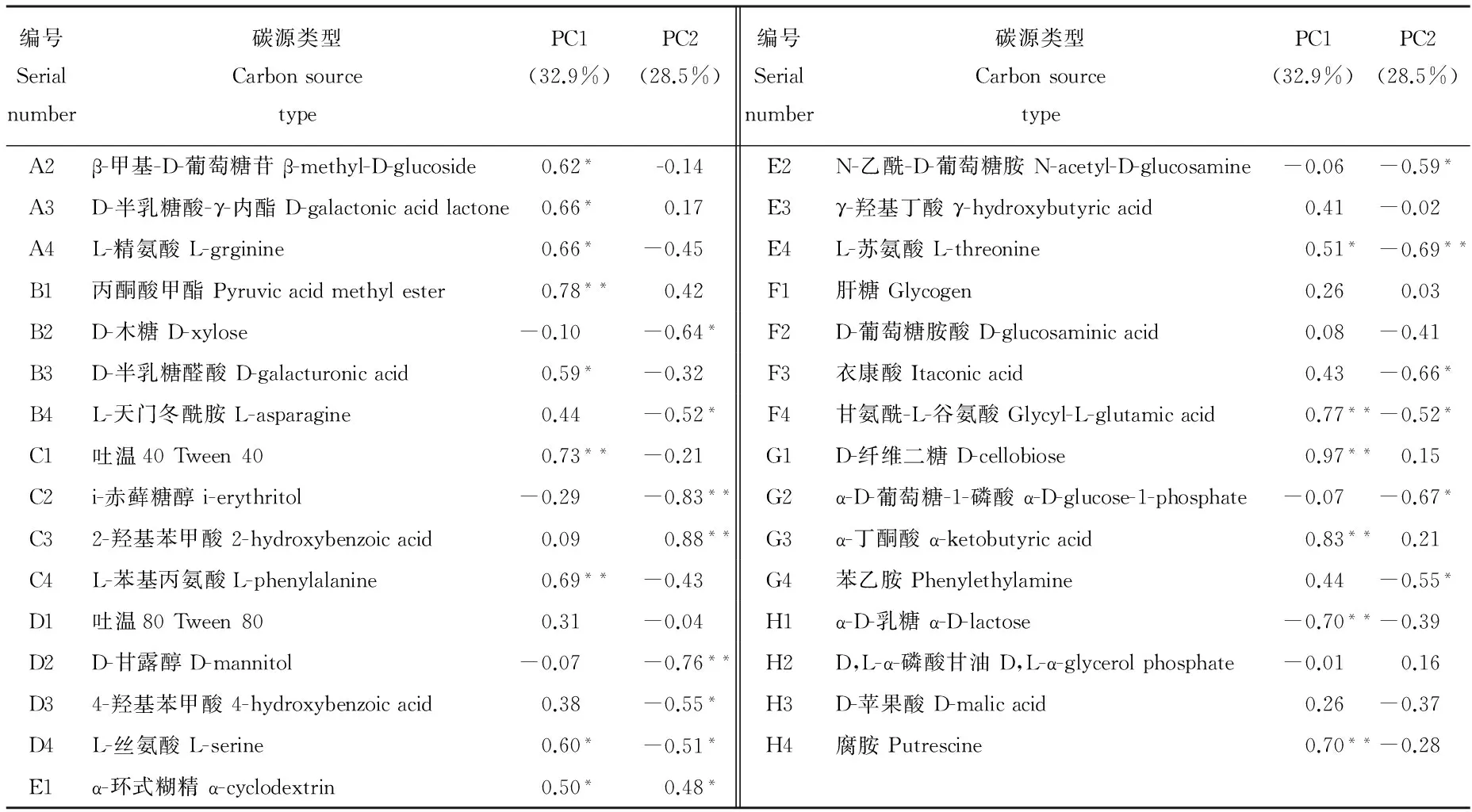
表4 主成分载荷因子Table 4 Loading factors of principle components
PC1: 主成分1 The first principal component;PC2:主成分2 The second principal component. *,**:分别表示在0.05和0.01水平下显著相关(双尾检验)。Significant correlations at the 0.05 and 0.01 levels(2-tailed test).
3 讨论与结论
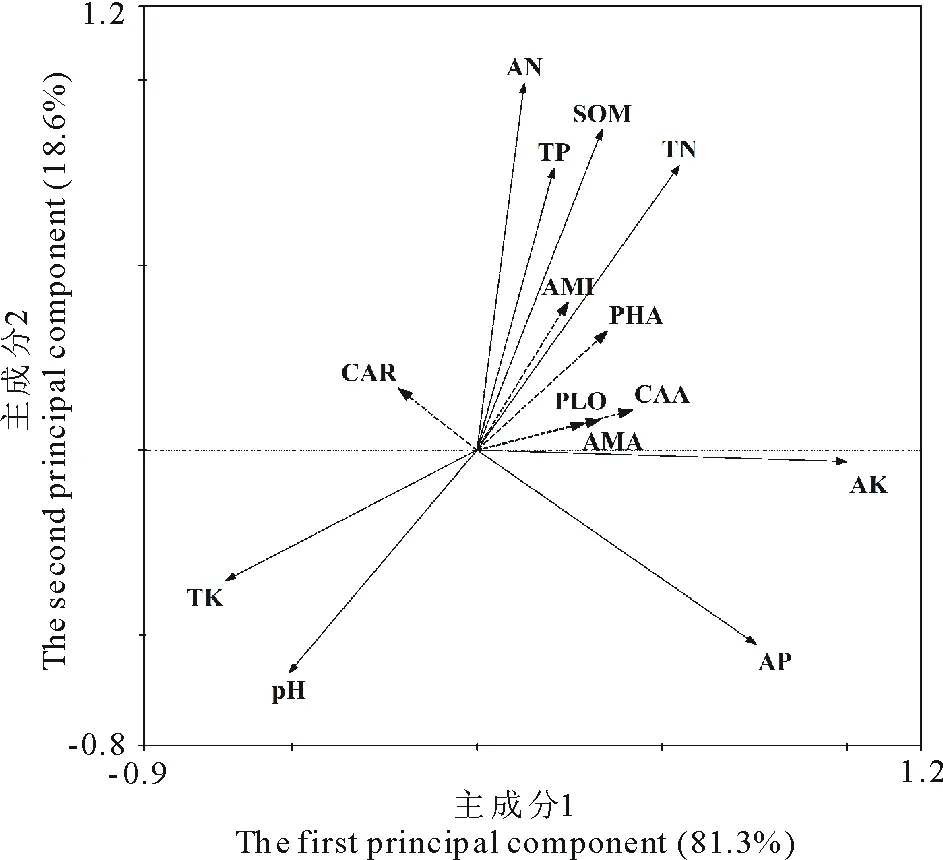
图4 蚁丘扰动高寒草甸土壤理化性质与土壤微生物群落功能特征的RDA分析 Fig.4 RDA for soil physicochemical properties and soil microbial function diversity in alpine meadow with ant-hill disturbanceCAR: 碳水化合物Carbohydrate; AMA: 氨基酸类Amino acid; CAA: 羧酸类 Carboxylic acid; PLO: 多聚物Polymer;PHA: 酚酸类Phenolic acid; AMI: 胺Amine; SOM: 有机质Organic matter; TN: 全氮 Total nitrogen; TP: 全磷Total phosphorus; TK: 全钾Total potassium; AN: 速效氮Available nitrogen; AP: 速效磷Available phosphorus; AK: 速效钾Available potassium.
3.1 蚁丘扰动对高寒草甸土壤理化性质的影响
蚂蚁可以将土壤中的营养成分“搬运”到土壤表层。已有研究表明,与邻近非蚁丘土壤相比,蚁丘扰动土壤的有机质、氮、磷和钾的含量都增加[28];刘任涛等[29]对科尔沁沙地流动沙丘掘穴蚁(Formicacunicularia) 蚁丘的研究表明,蚁丘土壤有机质和全氮增加;本研究中蚁丘干扰后,0~10 cm土层有机质、全氮、速效氮、速效磷、速效钾含量均显著高于10~20 cm土层(P<0.05),高寒草甸土壤养分发生明显改变,这与鱼小军等[30]就蚂蚁对东祁连山高寒草地生态系统的影响的研究结果相似。蚁丘扰动土壤中有机质、全氮、速效氮、速效磷、速效钾含量均升高可能与蚂蚁的粪便及排泄物、动物尸体和蚜虫蜜露等在巢内聚集以及蚂蚁在筑丘过程中巢内土壤覆盖了表层结皮和有机质,导致土壤微生物结皮和凋落物的腐烂分解有关[31]。蚁丘扰动对pH、全氮、全钾含量的影响不明显。蚂蚁使其活动范围内植物群落物种组成发生改变,引起植物产物的改变和凋落物有机质组分的变化,进而通过下行效应影响回归到土壤中养分的含量和组成。通过累积和分解,地上枯落物对陆地生态系统有物理、化学和生物作用[32]。土壤母质、养分本身特性等也影响着土壤的化学性质[33]。
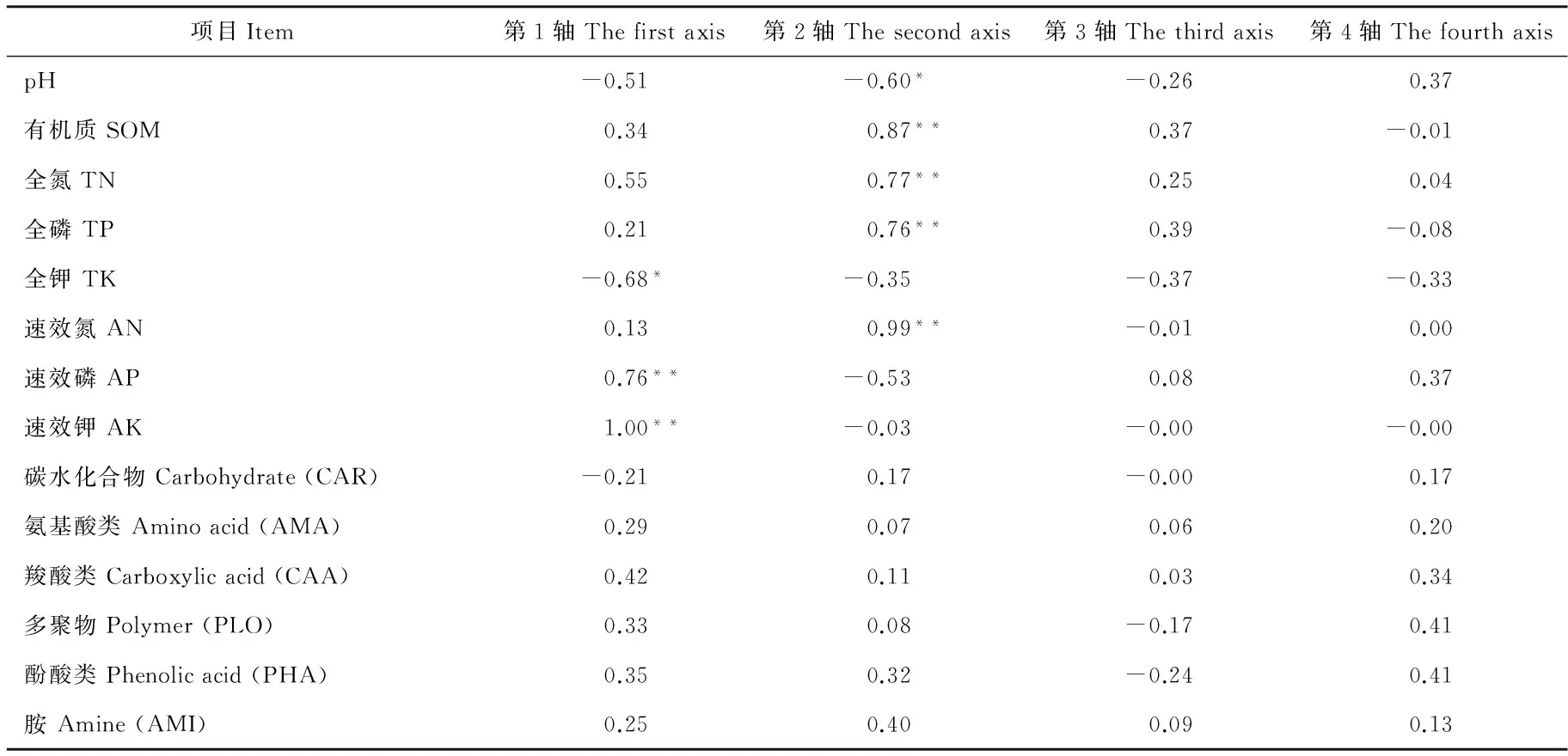
表5 RDA分析排序轴特征值及变异解释Table 5 Eigenvalues and variance percentage of soil environment factors explained by RDA
*,**分别表示在0.05和0.01水平下显著相关。
*,**:Significant correlations at the 0.05 and 0.01 levels.
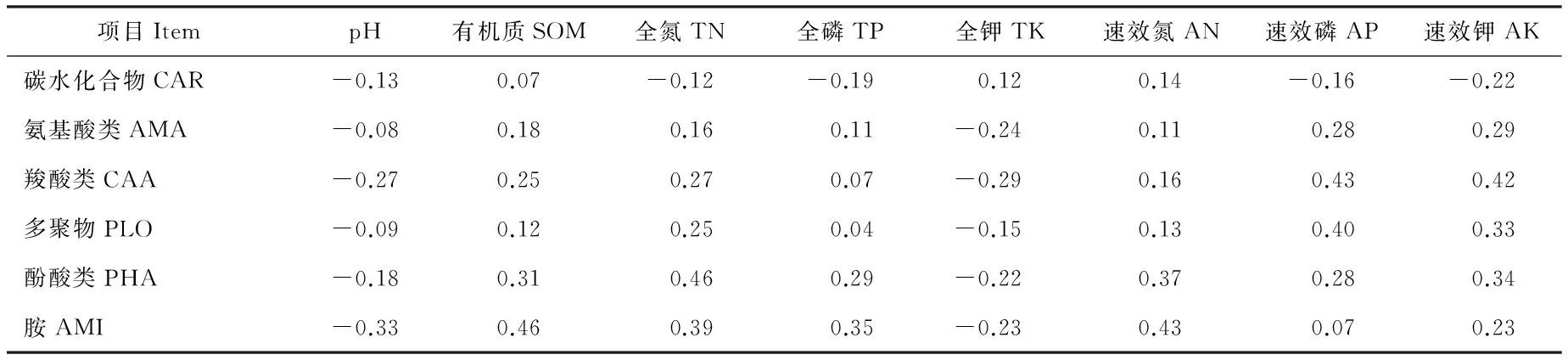
表6 蚁丘干扰下土壤理化性质与6类碳源的相关性分析Table 6 The Pearson correlation between soil physicochemical properties and six kinds of carbon resources
3.2 蚁丘对高寒草甸土壤微生物碳源利用代谢活性的影响
土壤微生物在土壤活跃成分中扮演着非常重要的角色,其代谢活性的变化可以反映土壤质量、土壤肥力的演变,是评价土壤质量的生物指标之一[34]。张萍等[35]的研究发现土地利用方式的不同对土壤微生物生物量和微生物多样性的影响也不同。AWCD值可以用来评价土壤微生物利用单一碳源能力的大小来指示土壤微生物的代谢活性[24]。本研究中AWCD值随着培养时间的延长呈指数型增长,蚁丘干扰后高寒草甸土壤微生物群落的代谢活性和代谢多样性增加,这可能是因为蚂蚁在不停地挖掘活动中将其排泄物和某些动物的尸体进行掩埋,而微生物则通过对动物尸体以及粪便的进一步分解,增加了微生物群落多样性[8,10],而植物根系本身和分泌物均是土壤微生物的主要碳源[33],这使土壤中的碳源含量和分布都有所增加,微生物对碳源的利用能力也就相应提高。土壤速效钾含量与微生物活性呈极显著正相关[36],在本研究中,蚁丘样地土壤速效钾含量显著高于对照样地,这说明土壤速效养分也是改变微生物代谢活性的原因之一。
土壤微生物多样性指数反映了土壤微生物多样性的不同方面,多样性指数可用于评价不同土壤的微生物群落多样性水平[37]。蚁丘扰动下的土壤微生物对碳源的利用率,Shannon-Weiner、Pielou和McIntosh指数均比未受蚁丘干扰的要高,因为蚁丘群落植物多样性和种子库多样性指数均显著高于对照[8,10,38-39]。
3.3 蚁丘扰动对高寒草甸土壤微生物功能多样性的影响
通过蚁丘干扰样地和对照样地土壤微生物对第1主成分轴和第2主成分轴碳源利用量及利用能力的比较发现,蚁丘干扰下的土壤与未受干扰的土壤微生物在底物碳源的利用能力上存在差异,土壤微生物利用的主要碳源是碳水化合物、氨基酸类和羧酸类。不同的多样性指标反映土壤微生物群落组成的不同方面,综合反映了土壤微生物群落的功能多样性[24]。此外,土壤容重、土壤孔隙度等都能够通过影响微生物的生存条件而影响其结构和功能[34]。
冗余分析表明,第1主成分轴和第2主成分轴解释总变异达到99.9%,土壤理化性质中有机质、全钾、全磷、全氮、速效钾、速效磷和速效氮是影响土壤微生物碳源代谢特征和功能的主要因素。土壤微生物对碳源利用的选择性可能与土壤微生物的组成结构和多样性的变化有关,从而影响土壤微生物碳源代谢活性和功能多样性的变化[20]。
总之,在草地生态系统中不同物种组成的群落可能存在着植物—土壤微生物特定的关系。例如:植物物种的共存可能会受到微生物群落组成和功能多样性的影响,反过来土壤微生物组成和多样性也会受到作为碳源基质的植物凋落物和土壤养分的质量和数量的影响。植物凋落物的化学特性较地上植物群落组成、物种丰富度、物种均匀度能更好地解释土壤的内部过程[40]。尽管蚂蚁在高寒草甸生态系统中的作用已有相关报道[8],但蚁丘形成过程中植物多样性与土壤微生物组成、多样性之间的关联性与反馈机制仍不清楚,值得更深入的研究。
References:
[1] Chen L Z, Wang Z W. The Impact of Human Activities on Ecosystem Diversity[M]. Hangzhou: Zhejiang Science and Technology Press, 1999. 陈灵芝, 王祖望. 人类活动对生态系统多样性的影响[M]. 杭州: 浙江科学技术出版社, 1999.
[2] Zhang G S, Li L, Li X L,etal. Effects of climatic changes of south Qinghai plateau on the alpine meadow. Acta Prataculturae Sinica, 1999, 8(3): 1-10. 张国胜, 李林, 李希来, 等. 青藏高原气候变化及其对高寒草甸牧草生长影响的研究. 草业学报, 1999, 8(3): 1-10.
[3] Zhou Y, Tan Z H, Bao G. Impacts of climate change on agriculture and its responses. Chinese Agricultural Science Bulletin, 2011, 27(32): 299-303. 周义, 覃志豪, 包刚. 气候变化对农业的影响及应对. 中国农学通报, 2011, 27(32): 299-303.
[4] Pierre H. Effects of grazing on plant species composition and spatial distribution in grassland of the Sahel. Plant Ecology, 1998, 138: 191-202.
[5] Persson I L, Danell K, Bergstrom R. Different moose densities and accompanied changes in tree morphology and browse production. Ecology Application, 2005, 15(4): 1296-1305.
[6] Xiao H Y, Liu H, Li B,etal. A study on the influence on grazing disturbance on soil fauna communities in subalpine meadows. Acta Prataculturae Sinica, 2012, 21(2): 26-33. 肖红艳, 刘红, 李波, 等. 放牧干扰对亚高山草甸土壤动物群落影响的研究. 草业学报, 2012, 21(2): 26-33.
[7] Li B. Ecology[M]. Beijing: Higher Education Press, 2004. 李博. 生态学[M]. 北京: 高等教育出版社, 2004.
[8] Meng F Q, Gao X M, Sun S C. Plant community succession on ant-hills of a sub-alpine meadow in Northwestern Sichuan, China: species composition and diversity. Plant Diversity and Resources, 2011, 33(2): 191-199. 蒙凤群, 高贤明, 孙书存. 川西北高寒草甸蚁丘植物群落演替:植物组成与物种多样性. 植物分类与资源学报, 2011, 33(2): 191-199.
[9] Mao Z H, Zhu J J. Effects of disturbances on species composition and diversity of plant communities. Acta Ecologica Sinica, 2006, 26(8): 2695-2701. 毛志宏, 朱教君. 干扰对植物群落物种组成及多样性的影响. 生态学报, 2006, 26(8): 2695-2701.
[10] Hou J H, Zhou D W, Jiang S C. The effect of hill-building activities of ants on the species diversity of plant communities in Songnen grassland. Acta Phytoecologica Sinica, 2002, 26(3): 323-329. 侯继华, 周道玮, 姜世成. 蚂蚁筑丘活动对松嫩草地植物群落多样性的影响. 植物生态学报, 2002, 26(3): 323-329.
[11] Folgarait P J. Ant biodiversity and its relationship to ecosystem functioning: a review. Biodiversity and Conservation, 1998, 7(9): 1221-1244.
[12] Kristiansen S M, Amelung W. Abandoned anthills ofFormicapolyctenaand soil heterogeneity in a temperate deciduous forest: morphology and organic matter composition. European Journal of Soil Science, 2001, 52(3): 355-363.
[13] Folgarait P J, Perelman S, Gorosito N,etal. Effects ofCamponotuspunctulatusants on plant community composition and soil properties across land-use histories. Plant Ecology, 2002, 163(1): 1-13.
[14] Dean W R J, Milton S J, Klotz S. The role of ant nest mounds in maintaining small scale patchiness in dry grasslands in central Germany. Biodiversity and Conservation, 1997, 6: 1293-1307.
[15] Laakso J, Setala H. Nest mounds of red wood ants (Formicaaquilonia): hot spots for litter-dwelling earthworms. Oecologia, 1997, 111(4): 565-569.
[16] Petal J, Kusinska A. Fractional composition of organic matter in the soil of anthills and the environment of meadows. Pedobiologia, 1994, 38: 493-501.
[17] Silvia C, Susana L S, Florencia F C. Response of ants to grazing disturbance at the central Monte Desert of Argentina: community descriptors and functional group scheme. Journal of Arid Land, 2014, 6(1): 117-127.
[18] Fierer N, Schimel J P, Holden P A. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry, 2003, 35: 167-176.
[19] Cao C Y, Yao J D, Han X S,etal. Soil microbes functional diversity in sand-fixingCaraganamicrophyllacommunities in Horqin Sandy Land. Chinese Journal of Applied Ecology, 2011, 22(9): 2309-2315. 曹成有, 姚金冬, 韩晓姝, 等. 科尔沁沙地小叶锦鸡儿固沙群落土壤微生物功能多样性. 应用生态学报, 2011, 22(9): 2309-2315.
[20] Hu L, A De L J, Zi H B,etal. Effects of plateau zokor disturbance and restoration years on soil nutrients and microbial functional diversity in alpine meadow. Chinese Journal of Applied Ecology, 2015, 26(9): 2794-2803. 胡雷, 阿的鲁骥, 字洪标, 等. 高原鼢鼠扰动及恢复年限对高寒草甸土壤养分和微生物功能多样性的影响. 应用生态学报, 2015, 26(9): 2794-2803.
[21] Wang R. Desertificates and Changes Remote Sensing Analysis on Meadow of Hongyuan[D]. Chongqing: Southwest Agricultural University, 2005. 王润. 红原草地荒漠化变化遥感分析[D]. 重庆: 西南农业大学, 2005.
[22] Ma Y L, Xin M, Song L Y,etal. A survery of ants (Hymenoptera: Formicidae) species and distribution in Ningxia. Journal of Agricultural Sciences, 2008, 29(1): 35-38. 马永林, 辛明, 宋伶英, 等. 宁夏蚁科昆虫重量累积分布调查. 农业科学研究, 2008, 29(1): 35-38.
[23] Bao S D. Soil Agricultural Chemistry Analysis[M]. Beijing: China Agriculture Press, 2008. 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2008.
[24] Xiang Z Y, Zhang L, Zhang Q F,etal. Soil nutrients and microbial functional diversity of different stand types in Qinghai Province. Scientia Silvae Sinicae, 2014, 50(4): 22-31. 向泽宇, 张莉, 张全发, 等. 青海不同林分类型土壤养分与微生物功能多样性. 林业科学, 2014, 50(4): 22-31.
[25] Chaerun S K, Pangesti N P, Toyota K,etal. Changes in microbial functional diversity and activity in paddy soils irrigated with industrial wastewaters in Bandung, West Java Province, Indonesia. Water Air and Soil Pollution, 2011, 217: 491.
[26] Classen A T, Boyle S I, Haskins K E,etal. Community-level physiological profiles of bacteria and fungi: plate type and incubation temperature influences on contrasting soils. FEMS Microbiology Ecology, 2003, 44(3): 319-328.
[27] Fang H, Yu Y, Chu X,etal. Degradation of chlorpyrifos in laboratory soil and its impact on soil microbial functional diversity. Journal of Environmental Sciences, 2009, 21: 380-386.
[28] Li Q X, Sheng L X, Zhou D W,etal. Effect of theLasisusflavusin Songnen grassland on structure and biomass of theLeymuschinensiscommunity. Chinese Journal of Ecology, 2003, 22(6): 49-52. 李庆新, 盛连喜, 周道玮, 等. 松嫩草原黄墩蚁对羊草群落组成与生物量的影响. 生态学杂志, 2003, 22(6): 49-52.
[29] Liu R T, Zhao H L, Zhao X Y,etal. Nest building activities and effect ofFormicacuniculariaon soil properties on mobile dune of Horqin sandy land. Journal of Desert Research, 2010, 30(1): 135-139. 刘任涛, 赵哈林, 赵学勇, 等. 科尔沁沙地流动沙丘挖掘蚁(Formicacunicularia)筑丘活动及其对土壤的作用. 中国沙漠, 2010, 30(1): 135-139.
[30] Yu X J, Pu X P, Huang S J,etal. Effects of ants (Tetramoriumsp. ) on eastern Qilian Mountains alpine grassland ecosystem. Acta Prataculturae Sinica, 2010, 19(2): 140-145. 鱼小军, 蒲小鹏, 黄世杰, 等. 蚂蚁堆东祁连山高寒草地生态系统的影响. 草业学报, 2010, 19(2): 140-145.
[31] Levan M A, Stone E L. Soil modifications by colonies of black meadows ants in a New York old field. Soil Science Society of America Journal, 1983, 47: 1192-1196.
[32] Li Q, Zhou D W, Chen X Y. The accumulation, decomposition and ecological effects of above-ground litter in terrestrial ecosystem. Acta Ecologica Sinica, 2014, 34(14): 3807-3819. 李强, 周道玮, 陈笑莹. 地上枯落物的积累、分解及其在陆地生态系统中的作用. 生态学报, 2014, 34(14): 3807-3819.
[33] Zheng H, Oyang Z Y, Wang X K,etal. Effects of forest restoration types on soil quality in red soil eroded region, Southern China. Acta Ecologica Sinica, 2004, 24(9): 1994-2002. 郑华, 欧阳志云, 王效科, 等. 不同森林恢复类型对南方红壤侵蚀区土壤质量的影响. 生态学报, 2004, 24(9): 1994-2002.
[34] Song C Q, Wu J S, Lu Y H,etal. Advances of soil microbiology in the last decade in China. Advances in Earth Science, 2013, 28(10): 1097-1105. 宋长青, 吴金水, 陆雅海, 等. 中国土壤微生物研究十年回顾. 地球科学进展, 2013, 28(10): 1097-1105.
[35] Zhang P, Dao Z L, Guo H J,etal. Influence of land use on numbers and diversity of soil microorganisms in Gaoligong Mountains. Acta Botanica Yunnanica, 1999, (Suppl, XI): 84-89. 张萍, 刀志灵, 郭慧军, 等. 高黎贡山不同土地利用方式对土壤微生物数量和多样性的影响. 云南植物研究, 1999, (Suppl, XI): 84-89.
[36] Hou X J, Wang J K, Li S P. Effects of different fertilization and plastic mulching on functional diversity of soil microbial community. Acta Ecologica Sinica, 2007, 27(2): 655-661. 侯晓杰, 汪景宽, 李世朋. 不同施肥处理与地膜覆盖对土壤微生物群落功能多样性的影响. 生态学报, 2007, 27(2): 655-661.
[37] Klose S, Acosta-Martínez V, Ajwa H A. Microbial community composition and enzyme activities in a sandy loam soil after fumigation with methyl bromide or alternative biocides. Soil Biology and Biochemistry, 2006, 38: 1243-1254.
[38] Chen Y W, Li X R, Su Y G,etal. Study on the eco-functions ofFormicacunicularia(Hymenoptera: Formicidae) in a revegetated area on the southeast fringe of Tengger Desert, North China. Acta Ecologica Sinica, 2007, 27(4): 1508-1514. 陈应武, 李新荣, 苏延桂, 等. 腾格里沙漠人工植被区挖掘蚁(Formicacunicularia)的生态功能. 生态学报, 2007, 27(4): 1508-1514.
[39] Belnap J, Lange O L. Biological Soil Crusts Structure, Function and Management[M]. Berlin: Springer, 2003.
[40] Meier C L, Bowman W D. Links between plant litter chemistry, species diversity, and below-ground ecosystem function. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(50): 19780-19785.
Responses of soil microbial community functional diversity to Camponotus herculeanus ant-hill disturbance in alpine meadows
YANG You-Fang1, ZI Hong-Biao1, LIU Min1, A DE Lu-Ji1, CHEN Yan1, WANG Chang-Ting*
CollegeofLifeScienceandTechnology,SouthwestUniversityforNationalities,Chengdu610041,China
To understand the function of the ants in the health and stability of an alpine meadow ecosystem we analyzed the effects of ant-hill disturbance on soil physical and chemical properties and microbial functional diversity in an alpine meadow using the Biolog-ECO microplate method. The results showed that while organic matter, total nitrogen, available nitrogen, available phosphorus and available potassium levels within the 0-10 cm soil depth were significantly higher than those in the 10-20 cm soil depth as expected (P<0.05), and that ant-hill disturbance had no significant effect on soil physicochemical properties (P>0.05). Ant-hill disturbance significantly improved soil microbial diversity index (specifically, values of Shannon-Weiner, Pielou, McIntosh and carbon utilization efficiency) and principal component analyses showed that ant-hill disturbance influenced soil microbial diversity and changed the microbial metabolism characteristics of soil microorganisms, to a status where carbohydrates, amino acids and carboxylic acid were the main carbon sources for the soil microbial community. Redundancy analyses indicated that metabolic rate and microbial functional diversity of soil microbial community was affected by soil nutrient levels. Therefore, the mining activities of the ants and associated accumulation of organic matter changed the soil microenvironment under the ant hills, resulting in changes to the soil microbial carbon source utilization and metabolism.
ant-hill; Biolog-ECO; soil microbial community; alpine meadows
10.11686/cyxb2016066
http://cyxb.lzu.edu.cn
2016-03-02;改回日期:2016-04-07
国家自然基金项目(31370542),国家重点基础研究发展计划(973计划)(2013CBA01807),中央高校基本科研业务费专项资金项目(2014NZYTD01)和西南民族大学研究生创新型科研项目(CX2016SZ070)资助。
杨有芳(1993-),女,青海互助人,在读硕士。E-mail:yangruobing123@163.com*通信作者Corresponding author. E-mail:wangct@swun.edu.cn
杨有芳, 字洪标, 刘敏, 阿的鲁骥, 陈焱, 王长庭. 高寒草甸土壤微生物群落功能多样性对广布弓背蚁蚁丘扰动的响应. 草业学报, 2017, 26(1): 43-53.
YANG You-Fang, ZI Hong-Biao, LIU Min, A DE Lu-Ji, CHEN Yan, WANG Chang-Ting. Responses of soil microbial community functional diversity toCamponotusherculeanusant-hill disturbance in alpine meadows. Acta Prataculturae Sinica, 2017, 26(1): 43-53.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
环境工程技术学报(2022年3期)2022-06-05 07:20:20
昆钢科技(2021年6期)2021-03-09 06:10:20
活力(2019年21期)2019-04-01 12:17:10
现代园艺(2017年21期)2018-01-03 06:41:42
绿色科技(2016年16期)2016-10-11 06:53:51
电源技术(2016年9期)2016-02-27 09:05:25
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
