
卧龙自然保护区拐棍竹笋生长发育初步研究
2016-12-24 03:30张明春黄金燕李德生周小平何胜山周应敏张和民
西华师范大学学报(自然科学版) 2016年3期
张明春,黄金燕,黄 炎,李德生,刘 巅,周小平,谢 浩,何胜山,周应敏,张和民
(中国保护大熊猫研究中心,四川 卧龙 623006)
卧龙自然保护区拐棍竹笋生长发育初步研究
张明春,黄金燕,黄 炎,李德生,刘 巅,周小平,谢 浩,何胜山,周应敏,张和民
(中国保护大熊猫研究中心,四川 卧龙 623006)
拐棍竹是卧龙国家级自然保护区内大熊猫的主要食物之一。对卧龙三个区域内拐棍竹笋的生长发育研究表明,无论是新笋出土时间(10 cm)、还是到达大熊猫可取食高度(30 cm)和停止取食高度(200 cm)的时间,都是随着海拔的升高而有所推迟。各区域之间同海拔高度下的新笋发育也存在显著差异,大熊猫可取食时间长度(停止取食时间-初始取食时间)在各个海拔高度下差异较小。野生大熊猫在水平方向上的移动能增加对竹笋的选择机会,而通过垂直迁移可以延长取食竹笋的时间。卧龙自然保护区拐棍竹新笋的整体存活率为48.64±8.43%。山体中部(2 000m—2 300m)的新笋基径较粗,之后会随着海拔升高逐渐减小。
拐棍竹;海拔;大熊猫;取食
食物是影响野生动物种群数量发展的一个重要因素[1]。大熊猫(Ailuropodamelanoleuca)虽然以竹为食,但仍然保留着食肉动物较简单的消化道,所以尽管食物资源丰富,但消化率却制约着它们的能量摄入水平[2],大熊猫在进化过程中甚至形成了通过降低活动和代谢水平来减少日常能量消耗的机制[3]。这些特殊的生理结构和生活习性,加上栖息地破碎化和严重的人为干扰,使大熊猫成为了世界上最珍稀濒危的野生动物之一[4]。对大熊猫而言,竹笋是竹子中最容易消化的部位,消化率可达40%[5]。所以一到新笋萌发季节,大熊猫会大量取食竹笋,以补充漫长冬季中食物质量较差所带来的体质消耗[2]。
拐棍竹(Fargesiarobusta)是大熊猫在卧龙地区的主食竹种之一,在新笋萌发的季节,当地的多数大熊猫都活动于拐棍竹林中,追逐撵食当年新发竹笋[6],称之为 “撵笋”现象。秦岭野生大熊猫在当地的巴山木竹(Bashaniafargesii)林内也有类似的活动规律[7]。为深入揭示大熊猫“撵笋”现象的驱动因素,我们对卧龙地区拐棍竹笋的发育规律进行了调查,并将结果报道如下。
1 研究方法
1.1 研究区域自然概况
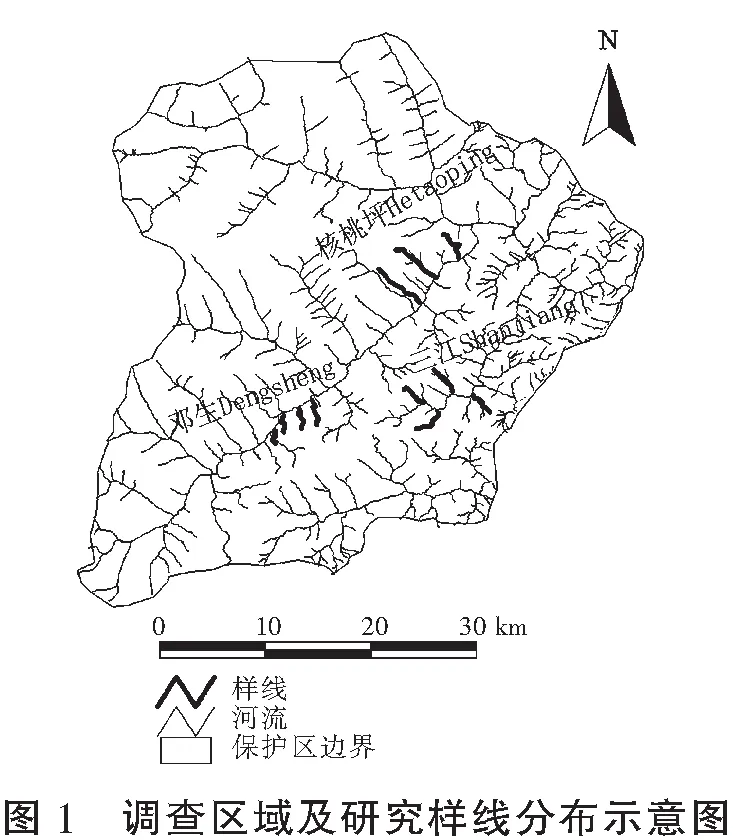
卧龙自然保护区位于四川省阿坝藏族羌族自治州汶川县西南部、邛崃山脉东南坡,始建于1963年,面积20×104hm2。根据全国第四次大熊猫调查结果,保护区内分布着100多只野生大熊猫。与大熊猫同域分布的国家重点保护动物包括金丝猴(Rhinopithecusroxellanae)、羚牛(Budorcastaxicolor)等。该地区属青藏高原气候区东缘,年均气温9.8 ℃,最高气温发生于7月,平均17 ℃,最低气温发生于1月,平均-1.7 ℃,年均降雨量1 800 mm,年均蒸发量874 mm,年日照时数927 h。
随着海拔的升高,研究区域植被由常绿阔叶林(1 150—1 600m)过渡到常绿落叶阔叶混交林(1 600—2 000m),再到针阔混交林(2 000—2 600m),然后是亚高山针叶林(2 600—3 600m),再往上就是高山灌丛草甸和高山流石滩稀疏植被(3 800m以上)。拐棍竹分布在海拔1 800—2 500m(个别地区向下分布至1 600m,向上分布至2 650m) 的林木下层,每年4月中旬出土,70—80 d长成成竹。
1.2 数据收集
在卧龙保护区内,于拐棍竹分布起始海拔不同的核桃坪、邓生和三江3个研究区域,选择人为活动较少的竹林中设置3—4条样线(见图1),样线间距离大于1 km。在海拔整百米处(±5m),于行进道路的两侧各设置5个1m×1m的小样方。从2015年4月中旬开始,待新笋出土约10cm高(因竹林下的枯枝落叶层较厚,低于10cm难于被发现)时,用卷尺测量每根新发竹笋的高度,用数显游标卡尺测量出对应的基径,并标记好每根新笋在样方中的位置,每5 d重复测量一次,直到新笋高度超过大熊猫取食的高度(200 cm)[6,8]。在10月时记录各小样方中存活的老笋数目,其它因病虫害、野生动物包括大熊猫、小熊猫(Ailurusfulgens)取食等原因致死的当年新笋记录为死亡。
1.3 数据分析
计算各样线上当年新笋的存活率,用单因素方差分析(One-Way ANOVA,因数据符合正态分布)比较核桃坪、邓生和三江保护站三个区域间新笋的存活率差异。以4月1日为1,4月2日为2……以此类推,将日期变量数字化,并假设新笋在两次测量间匀速生长,求出各新笋到达10cm、30cm、200cm高度的时间。根据胡锦矗等[6]和秦自生等[8]中的记录,将其分别作为出土高度、大熊猫初始取食高度和停止取食高度,用单因素方差分析(数据呈正态分布)和Kruskal-Wallis H检验(数据不呈正态分布)分析新笋到达这三个高度的时间随着海拔高度和区域改变而发生的变化,在存在显著差异的基础上进一步的多重比较采用Bonferroni分析或者Mannay Wittney U检验。用Spearman相关分析检验这三个时间之间的相关关系。
将测得的新笋基径数据,分区域和海拔进行成对样本t检验(Paired-Samples T Test,数据呈正态分布),或进行相关样本检验(Two-Related-Samples Test,数据不呈正态分布)。对数据初步检测发现第1、2次的测量数据因新笋未完全出土而变化较大,最后两次测量的直径也因基部箨壳掉落而有所减小,中间几次测量相互间差别不明显(P>0.05),故取第5次的测量结果代表新笋基径大小,数据经检验符合正态分布,因此用单因素方差分析检验各区域新笋基径随着海拔的升高而发生的变化,以海拔为控制变量,用协方差分析(General Linear Model:Univariate)检验各区域间新笋基径大小的差异,并用Bonfferoni多重比较检验两两之间的差异。用到达停止取食高度的时间减去到达初始取食高度的时间得到大熊猫对该新笋的可取食时间长度,将海拔分为低海拔(1 700—1 900m)、中间海拔(2 000—2 300m)和高海拔(2 400—2 600m)三个部分,对核桃坪和邓生区域用单因素方差分析(数据呈正态分布),用Kruskal-Wallis H检验分析三江区域(数据不呈正态分布)的新笋取食时间长度在这三个区域内的三个海拔范围间的差异,并用Mannay Wittney U检验分析三江区域三个海拔范围间两两之间的差异。
2 结 果
2015年4月至6月,我们调查了三个区域的11条样线,共测量新发竹笋2 627株,存活并纳入分析的竹笋1 259株,整体存活率为48.64±8.43%。存活率在三个区域间没有太大的差异(F=3.79,P=0.07)。
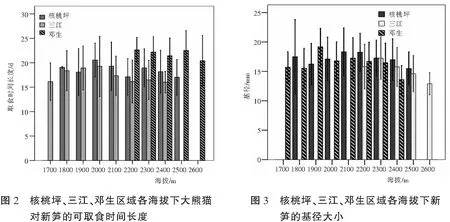
除了三江区域海拔1 800m的竹笋发育有所延迟以外,无论是新笋出土时间,还是达到大熊猫初始取食高度和停止取食高度,都是随着海拔的升高而逐渐延迟。相关分析结果显示这三个时间彼此之间的相关性较大(r>0.5,P<0.001),三个地区的当年新笋,在这三个时间上的海拔差异都极为显著(P<0.001),从各地区的海拔最低点到最高处,竹笋的发育时间延迟约10天左右(表1);各个区域同海拔高度的竹笋发育之间也存在着显著性差异(P<0.05),只是在海拔2 500 m处,核桃坪与邓生区域间的新笋到达初始取食高度的时间没有太大的差别(F=3.38,P=0.07)。大熊猫对竹笋的可取食时间长度方面,核桃坪区域(F=2.34,P=0.10)和邓生区域(F=1.22,P=0.27)在三个海拔范围内差别不大,但在三江区域差别较大(x2=8.68,df=2,P=0.01) (图2)。Mannay Wittney U检验显示低海拔范围内的竹笋与中、高海拔处有显著差异(P<0.05),但中、高海拔部分之间无显著差异(Z=1.22,P=0.22)。
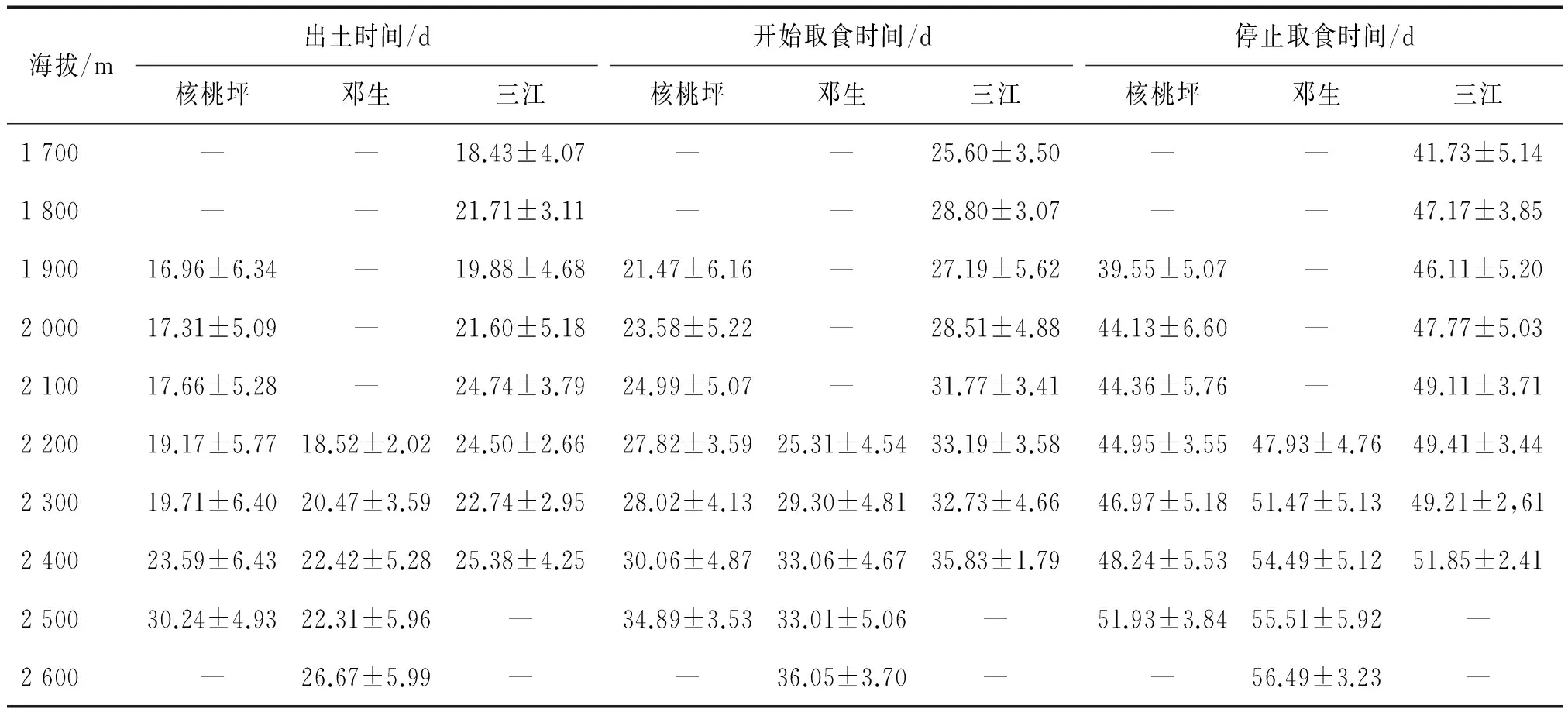
表1 核桃坪、邓生及三江区域各海拔下的拐棍竹新笋出土时间、大熊猫开始取食时间及停止取食时间
协方差分析结果显示,三个区域间的竹笋基径存在着显著差异(F=8.88,P=0.00),且其与海拔因素之间存在着交互作用(F=9.89,P=0.00)。进一步的多重比较显示核桃坪区域的竹笋基径(17.30±3.60)略大于三江区域(16.78±3.47),但差异并不显著(P>0.05),它们比邓生区域的新笋(15.32±3.40)基径更大(P<0.05)。三个区域的竹笋大小均具有海拔差异(P<0.05),低海拔部分的新笋基径与其它部分的差别不大,略小于中间部分的,又略大于高海拔部分的(P>0.05),海拔到了2 400m后,新笋直径随着海拔的升高逐渐减小(图3),这部分的竹笋基径显著小于中间海拔部分(P<0.05)。
3 讨 论
拐棍竹笋的发育不仅与生长环境中的光照和土壤肥力有关[9],还受地震、动物取食等干扰因素的影响[10],但决定其萌发和生长速度的最关键因子还是环境中的温度高低[8]。随着海拔的升高,气温会逐渐降低,所以无论是新笋的出土时间,还是大熊猫的开始取食时间和停止取食时间,海拔较高处都会晚于低海拔地区。坡度较大的地区不是大熊猫最喜欢选择的生境或取食场所[11,12],但尽管拐棍竹分布区域的坡度往往较大,当地的野生大熊猫在新笋长到一定高度时就会进入拐棍竹林中取食。卧龙地区海拔1 800—1 900m属于常绿落叶阔叶林混交林带[13],拐棍竹林中光照较差,山势险峻,土壤有机质受雨水冲刷流失严重而肥力较差,这可能是造成其基径比中间部位海拔处还小的原因。又因为这一带接近河谷地区带,人为干扰严重,采笋和放牧都会破坏拐棍竹的生长,所以拐棍竹分布比较稀疏,大熊猫取食竹笋的活动往往发生在海拔2 000m以上,这些区域不仅人为干扰较少,而且周围的竹笋质量也较好。大熊猫的家域面积较大[6,14],所以即使它们每天的摄食量超过40kg,取食的竹笋也不会超过当地所发新笋总量的10%[7],大熊猫对竹笋拥有较充分的选择机会。
三个受调查区域的竹林均属正常发育的成熟林,竹笋存活率保持在50%左右,与秦自生等[8]的研究结果相似。三个区域所处环境各异,水分和土壤肥力等也有所差别,所以竹笋的生长和发育在相同海拔下也有较大的差异。实际上,即使处于同一个小样方中,不同时间萌发的竹笋也可能有不同的发育速度,初始发育相近的一批竹笋在后期可能继续相似,也可能发育程度迥异,发生这些变化的原因还需要进一步的研究进行确定。但从整体而言,这些变化没有海拔跨度方向上明显而有规律。利用竹笋发育在海拔跨度上的差异,大熊猫最初可以在它们家域内的较低海拔处往来搜寻新笋,直到新笋长高而且变得不再适口时,它们又会迁移到海拔较高的区域,取食此时已经长高了的新笋。所以栖息地内较大的海拔跨度可以延长大熊猫取食竹笋的时间长度,帮助大熊猫增强体质,考虑到拐棍竹发笋期与大熊猫发情期时间相近[15],大量取食竹笋或许可以促进其增加繁殖的成功率。
[1] 孙儒泳.动物生态学原理(第三版)[M].北京:北京师范大学出版社,2001.
[2] 胡锦矗.大熊猫研究[M].上海:上海科技教育出版社.2001.
[3] NIE Y G,JOHN R,SPEAKMAN J R,et al.Exceptionally low daily energy expenditure in the bamboo-eating giant panda[J].Science,2015,349:171-174.
[4] IUCN.The IUCN Red List of Threatened Species[EB/OL].(2015-04-13)[2015-10-25].http:∥www.iucnredlist.org.
[5] SCHALLER G B,HU J C,PAN W S,et al.The giant pandas of Wolong[M].Chicago:The University of Chicago Press.1985.
[6] 胡锦矗.卧龙的大熊猫[M].成都:四川科技出版社,1985.
[7] 潘文石,吕 植,朱小健,等.继续生存的机会[M].北京:北京大学出版社,2001.
[8] 秦自生,TYLER A,蔡绪慎,等.卧龙大熊猫生态环境的竹子与森林动态演替[M].北京:中国林业出版社.1993.
[9] 周世强,黄金燕,张亚辉,等.野化培训大熊猫采食和人为砍伐对拐棍竹无性系种群更新的影响[J].生态学报,2009.29(9):4804-4814.
[10] 廖丽欢,徐 雨,冉江洪,等.汶川地震对大熊猫主食竹—拐棍竹竹笋生长发育的影响[J].生态学报.2012,32(10):3001-3009.
[11] ZHANG Z J,ZHAN X J,YAN L,et al.What determines selection and abandonment of a foraging patch by wild giant pandas (Ailuropoda melanoleuca) in winter? [J].Environ Sci Pollut Res,2009,16:79-84.
[12] 杨春花,张和民,周小平,等.大熊猫(Ailuropoda melanoleuca)生境选择研究进展[J].生态学报,2006,26(10):3442-3453.
[13] 张科文,胡锦矗,余志伟,等.卧龙自然保护区动植物资源及保护[M].成都:四川科学技术出版社,1990.
[14] ZHANG Z J,SHEPPARD J K,SWAISGOOD R R,et al.Ecological scale and seasonal heterogeneity in the spatial behaviors of giant pandas[J].Integrative Zoology,2014,9(1):46-60.
[15] 魏辅文,张泽钧,胡锦矗.野生大熊猫生态学研究进展与前瞻[J].兽类学报,2011,31(4):412-421.
Variations of the Fargesia robusta Shoots at Different Elevations in Wolong Nature Reserve,China
ZHANG Mingchun,HUANG Jinyan,HUANG Yan,LI Desheng,LIU Dian,ZHOU Xiaoping,XIE Hao,HE Shengshan,ZHOU Yingming,ZHANG Hemin
(China Conservation and Research Center for the Giant Panda,Wolong Sichuan 623006,China)
Fargesiarobustais one of the most important foods for the giant panda.The research for the shoots in three areas of Wolong Nature Reserve showed that time of the new shoot for sprouting (10cm),initial feeding(30cm) and finishing feeding(200cm) by the giant panda would delay with the elevation increasing.The difference was also significant in different areas at the same elevation.The period of the giant panda feeding to the shoots (time of finishing feeding-time of initial feeding) had little difference in different elevations.Giant pandas had more chances to choose shoots via wandering at horizontal direction,and had longer period to fetch shoots via Vertical migrations.Survival rates of the shoots were 48.64±8.43%, the basal diameter was the biggest at middle part (2 000—2 300m) of all areas, and then decreased with the increase of the elevation.
Fargesiarobusta;elevation;giant panda;feeding
1673-5072(2016)03-0249-04
2015-10-29 基金项目:川港卧龙持续合作计划-香港特别行政区资助项目(HK04);大熊猫国际合作资金项目(SD1115);卧龙大熊猫俱乐部自然基金青年项目(WPC-20(15-18)-03)
张明春(1973—),男,四川营山人,高级工程师,主要从事生态学研究。
张和民(1963—),男,四川营山人,教授,主要从事大熊猫保护研究。E-mail: wolong_zhm@126.com
Q958.1
A
10.16246/j.issn.1673-5072.2016.03.002
猜你喜欢
水泵技术(2021年2期)2021-07-31
养生保健指南(2019年10期)2019-12-16
四川林业科技(2019年5期)2019-07-07
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
小猕猴学习画刊(2018年12期)2018-01-19
小小说月刊(2017年7期)2017-07-11
湖北林业科技(2017年2期)2017-05-25
故事大王(2017年2期)2017-05-04
辽金历史与考古(2017年0期)2017-02-06
