
榕-蜂系统中影响传粉榕小蜂繁殖及存活的因素研究
2016-12-22 09:21邓国宾李宗波
生物学杂志 2016年6期
张 媛,邓国宾,李宗波
(1.西南林业大学 云南生物多样性研究院,昆明 650224;2.西南林业大学林学院 云南省森林灾害预警与控制重点实验室,昆明 650224)
榕-蜂系统中影响传粉榕小蜂繁殖及存活的因素研究
张 媛1,邓国宾1,李宗波2
(1.西南林业大学 云南生物多样性研究院,昆明 650224;2.西南林业大学林学院 云南省森林灾害预警与控制重点实验室,昆明 650224)
榕属植物(Ficus)及其传粉昆虫榕小蜂(Agaonidae)是自然界协同进化的经典模型,榕小蜂资源是实现榕-蜂繁殖衔接的重要因素。对分布在西双版纳地区的两种重要榕属植物:鸡嗉子榕(Ficussemicordata)和高榕(Ficusaltissima)进行人工控制性放蜂实验,分别验证季节变化、母代雌蜂年龄、榕果雌花期延长对小蜂后代数量、败育率和后代体长的影响。结果表明:季节变化对小蜂后代数量、败育率无影响,而对小蜂后代体大小有一定影响。而随着母代雌蜂年龄的增加和榕果雌花期的延长,都会对小蜂后代数量产生负面影响,使败育率上升,后代体长变小。这一结果有助于了解榕-蜂系统对互利共生双方繁殖的调节,为进一步研究榕-蜂系统的稳定性提供了依据。
互利共生;传粉榕小蜂;繁殖效率;雌花期;窝卵数
自然界中物种的互利共生关系广泛存在,互利共生双方均可从系统中获得一定的服务或是利益,从而得到生存和繁殖的保障[1]。传粉榕小蜂(Hymenoptera: Agaonidae)及其寄主榕树Ficusspp.的关系,是自然界中最典型的动植物间互利共生关系[2-3]。母代传粉榕小蜂通过榕果苞片进入到雌花期榕果内,为榕果内的雌花传粉,并将卵产在雌花子房内,该部分小花变为瘿花,子代小蜂后代在瘿花中发育成熟后便在榕果果腔内进行交配,然后携带花粉的子代小蜂再从苞片口飞出,寻找其他处于雌花期的榕果,开始下一个生命循环[4]。榕树和传粉榕小蜂之间是少有的一对一关系,即每种榕树通常由一种传粉榕小蜂为其传粉,而传粉小蜂也只能借助寄主榕树才能繁衍后带,二者之间相互依存,一方的崩溃必然带来整个系统的瓦解。榕-蜂之间的这种非常紧密的互惠共生关系使其成为研究物种间互惠共生和协同进化的经典材料[5-6]。榕树从繁殖方式上可以分为雌雄同株和雌雄异株两种:在雌雄同株的榕果内,一部分雌花受精花育为种子,一部分雌花被榕小蜂用来产卵繁殖后代;在雌雄异株的榕果内,种子和小蜂的产生分属在不同性别的榕树果内,雌株果内只能产生种子,而榕小蜂和花粉只出现在雄株果内[7]。
榕属植物一年四季均挂果,榕果为兽类、鸟类等食果动物提供了不间断的食物来源,尤其是在冬天,榕果成为稳定的食物来源,这为维持热带雨林地区的物种多样性起到了重要作用[8-9],使榕树成为热带雨林的关键植物类群[10-11]。研究榕树-榕小蜂的繁殖特征及系统进化稳定机制不但可以丰富物种间协同进化理论,还可为物种多样性的保护提供科学依据[12]。榕属植物虽然在热带亚热带地区较为常见,但是,单种榕树在热带亚热带地区的分布密度一般较低[13],加之榕小蜂成虫寿命又非常短暂(一般为数小时至2 d)[14],这就使得榕-蜂的相遇非常困难。但在自然界中,大约只有1%的传粉榕小蜂可以成功找到处于雌花期的寄主榕树,榕-蜂之间依靠榕果释放的挥发性化学物质实现通讯[15-16],大部分传粉榕小蜂在找到合适时期的寄主榕树之前就死去[17]。最近的研究发现:榕果所处的时期,小蜂年龄、繁殖季节等都会对榕果的繁殖资源分配产生影响[18-21],但这些因素是通过何种途径对榕-蜂繁殖进行调配仍不明确。本研究将以分布在西双版纳地区的两种榕树为实验材料,研究繁殖季节以及榕果和传粉母代年龄对传粉榕小蜂繁殖及存活产生的影响,实验结果将为我们研究榕树-榕小蜂繁殖稳定机制提供依据。
1 材料与方法
1.1 研究区概况
试验地为中国科学院西双版纳热带植物园及周边。该植物园位于云南省西双版纳州勐腊县勐仑镇,地处印度马来热带雨林区北缘,终年受西南季风控制,属低山宽谷型地貌,热带季风气候。西双版纳热带植物园的年均气温21.8℃,6月最热,平均气温为25.7℃,最冷1月,平均气温为16.0℃,终年无霜[22]。根据降雨量可分为旱季(11月—次年4月)和雨季(5月—10月),旱季还可分为雾凉季(11月—次年2月)和干热季(3月—4月),年均相对湿度为85%,全年日照时间为1859 h[23]。榕属植物是该西双版纳地区的关键类群之一[24]。
1.2 研究材料
高榕(Ficusaltissima),雌雄同株,为桑科榕属榕亚属(Urostigma)的半附生的高大乔木,高度可达40 m以上。高榕唯一的传粉榕小蜂为Eupristinaaltissima,传粉类型为主动传粉,隶属于膜翅目(Hymenoptera),小蜂总科(Chalcidoidea),榕小蜂科(Agaonidae),雌雄异型,雌蜂有翅,雄蜂无翅。蜂源为采集野外雄花期榕果,将其置于纱网贷,待其自然出蜂获得。
鸡嗉子榕(Ficussemicordata)隶属于荨麻目(Urticales)桑科(Moraceae)榕属聚果榕亚属(Sycomorus)。雌雄异株榕树,小乔木,高3~10 m。鸡嗉果的唯一传粉昆虫为Ceratosolengravelyi,传粉类型为主动传粉,隶属膜翅目(Hymenoptera)小蜂总科(Chalcidoidea)榕小蜂科(Agaonidae)。雌雄异型,雌蜂有翅,雄蜂无翅。蜂源为采集野外雄花期榕果,将其置于纱网贷,待其自然出蜂获得。
1.3 季节对榕小蜂繁殖及存活的影响
试验选取分布在中国科学院西双版纳热带植物园周边的3株鸡嗉子榕雄树,2年时间里,每周对样树进行物候观测。当发现榕果接近雌花期时,改为每周观测2次,鸡嗉子榕果到达雌花期时苞片会明显松动,变化肉眼可见,开始单果引入传粉榕小蜂,根据传粉榕小蜂对果的行为反应判断该果是否到达雌花期。具体方法是:将单头当天出蜂的活力较好的传粉榕小蜂引到鸡嗉子榕果果壁上,如果榕小蜂在5 min内没有表现任何进入顶生苞片的行为,就判断榕果为还未到达雌花开花期,并继续用纱网袋套住,第2天用同样的方法进行测定,如果5 min内,传粉榕小蜂表现出钻入顶生苞片的行为,就认为此榕果已经到达雌花期,将榕果标记为雌花期第1天。在干热季、雨季、雾凉季时分别为雌花期第1天的榕果放入1头传粉小蜂,每个处理30个榕果,放入小蜂后的榕果用纱网袋重新套上防止其他昆虫的干扰。待榕果发育到雄花期时,单果分装于纱网袋,带回实验室统计每果内小蜂后代数量和败育瘿花数量,败育率为发育不良瘿花数与总瘿花数(小蜂与发育不良瘿花数之和)的比值。可得到自然状态下,小蜂在不同季节的繁殖变化情况。
测量后代传粉榕小蜂个体大小以确定季节对小蜂后代发育的影响,选取不同进蜂时期的雄株果,每个年龄的雄株果为5个,每个榕果取10只雌蜂,将其浸泡在80%的乙醇中,乙醇中加入一滴丙三醇以保证小蜂不变形,然后在奥林巴斯 SZX12显微镜下测量每只小蜂的体长(不包括产卵器长度)。
1.4 母代雌蜂年龄对榕小蜂繁殖及存活的影响
在前期的工作中,我们发现仅有高榕传粉小蜂的寿命可达2 d,故选用高榕来测试母代雌蜂年龄对繁殖的影响。首先用纱网袋套住处于雌花前期的榕果,以防止传粉榕小蜂传粉或非传粉小蜂产卵的影响,每天观察榕果,一旦发现榕果到达雌花期(确定方法同上),我们就将单只小蜂引入雌花期第1天的单个榕果内,引蜂试验分两种组合:一种是用当天出果的传粉榕小蜂进行引蜂试验,另一种是将刚出果的传粉榕小蜂放入15℃的恒温箱内保存24 h,第2天将其引入榕果。放入15℃恒温箱的目的是让传粉榕小蜂在一个相对较低的温度下,减少活动,以保持其活力。两类准备好的传粉榕小蜂都引入雌花期第1天的榕果,以保证榕果质量一致。将传粉榕小蜂放入榕果后,又重新用纱网袋套住榕果,直到其发育到雄花期,共收获27个第1天寿命的传粉榕小蜂授粉榕果和30个第2天寿命的传粉榕小蜂授粉榕果。单果分装于纱网袋内,统计传粉榕小蜂后代数量和发育不良瘿花数。此外,我们还测量了不同组合小蜂雌性后代大小,方法见1.3。
1.5 榕果发育时期对榕小蜂繁殖及存活的影响
用1.3中的方法确定雌花期的第1天,在鸡嗉子雄果处于雌花期1、4、12 d时分别单果引入单只当天出蜂的活力较好的传粉榕小蜂,然后将纱网袋重新套上榕果,让榕果正常发育。待榕果发育到雄花期时,将榕果单果采集分装,统计传粉小蜂后代数,发育不良的瘿花数。同样测量不同处理的小蜂后代体长,方法同1.3。
1.6 数据整理与分析
采用单因素方差分析 (ANOVA) 分析母代雌蜂年龄对后代数量、败育率及后代体大小的影响;采用单因素方差分析 (ANOVA) 中的多重比较分析榕果时期、季节变化对小蜂后代数量、败育率及后代体大小的影响,在满足方差齐性时采用ANOVA中的LSD检验,在不满足方差齐性时采用ANOVA 种的Tamhane’s T2检验。P< 0.05时差异显著。采用Excel进行数据整理和作图,采用SPSS Statistic 17.0 软件包程序进行数据分析。
2 结果与分析
2.1 季节对榕小蜂繁殖及存活的影响
如图1-A所示,干热季、雨季和雾凉季,在放一头鸡嗉子榕小蜂时,后代数量在季节之间无显著差异(ANOVA:F=0.57,P>0.05);小蜂后代的败育率在3个季节之间同样无明显差异(ANOVA:F=1.01,P>0.05) ,图1-B;而小蜂后代体大小在不同季节差异显著(ANOVA:F= 4.89,P<0.05),其中干热季的小蜂后代体长显著大于雨季和雾凉季(Tamhane:P<0.05),雨季和雾凉季之间小蜂体大小差异不显著(Tamhane:P>0.05),图1-C。
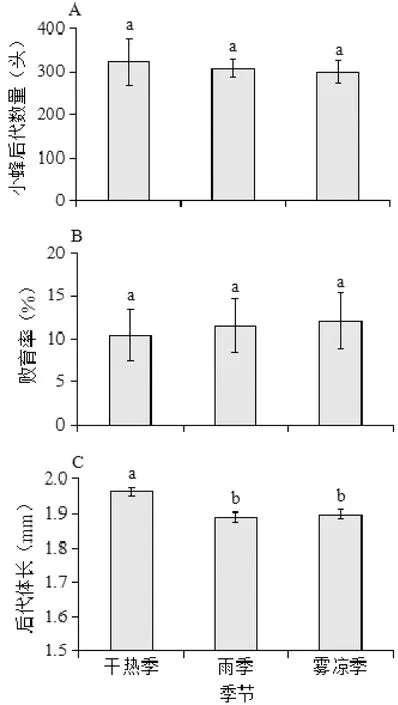
图1 季节对鸡嗉子榕传粉小蜂后代数量(A)、败育率(B)及后代体长(C)的影响
Fig 1 The impact of season on total number (A),abortion rate (B) andbody size (C) of pollinating fig wasps offspring inFicussemicordata
注:柱状图上的不同字母代表差异显著;下同
2.2 母代雌蜂年龄对榕小蜂繁殖及存活的影响
当高榕母代传粉榕小蜂的年龄增加24 h后,高榕内一头雌蜂所繁殖的后代数量显著降低(ANOVA:F= 32.40,P< 0.05),图2-A,且榕果内小蜂败育率上升(ANOVA:F= 4.34,P< 0.05),图2-B。此外,繁殖雌蜂的年龄可显著降低后代个体大小(ANOVA:F= 38.95,P< 0.05),图2-C。
2.3 榕果发育时期对榕小蜂繁殖及存活的影响
在鸡嗉子榕果雌花期第1天进果的传粉榕小蜂,能产生最多的后代数量(322±53.8)头,小蜂子代数量随着母代进果时期的推迟而显著减少(ANOVA:F= 4.88,P< 0.05),第12天进蜂的榕果所产生的后代数量显著少于第1天和第4天的后代数量(LSD:P< 0.05),图3-A。榕果内小蜂后代的败育率也随着榕果雌花期的延长而上升(ANOVA:F= 9.21,P< 0.05),同样是第12天时败育率显著高于第1天和第4天(LSD:P< 0.05),图3-B。此外,小蜂后代体大小也随着母代雌蜂进果时间推迟而下降(ANOVA:F= 19.23,P< 0.05),3个处理间均有显著差异(LSD:P< 0.05),图3-C。

图2 母代高榕雌蜂年龄对传粉小蜂后代数量(A)、败育率(B)及后代体长(C)的影响
Fig 2 The impact of foundress age on total number (A),abortion rate (B)and body size (C) of pollinating fig wasps offspring inFicusaltissima
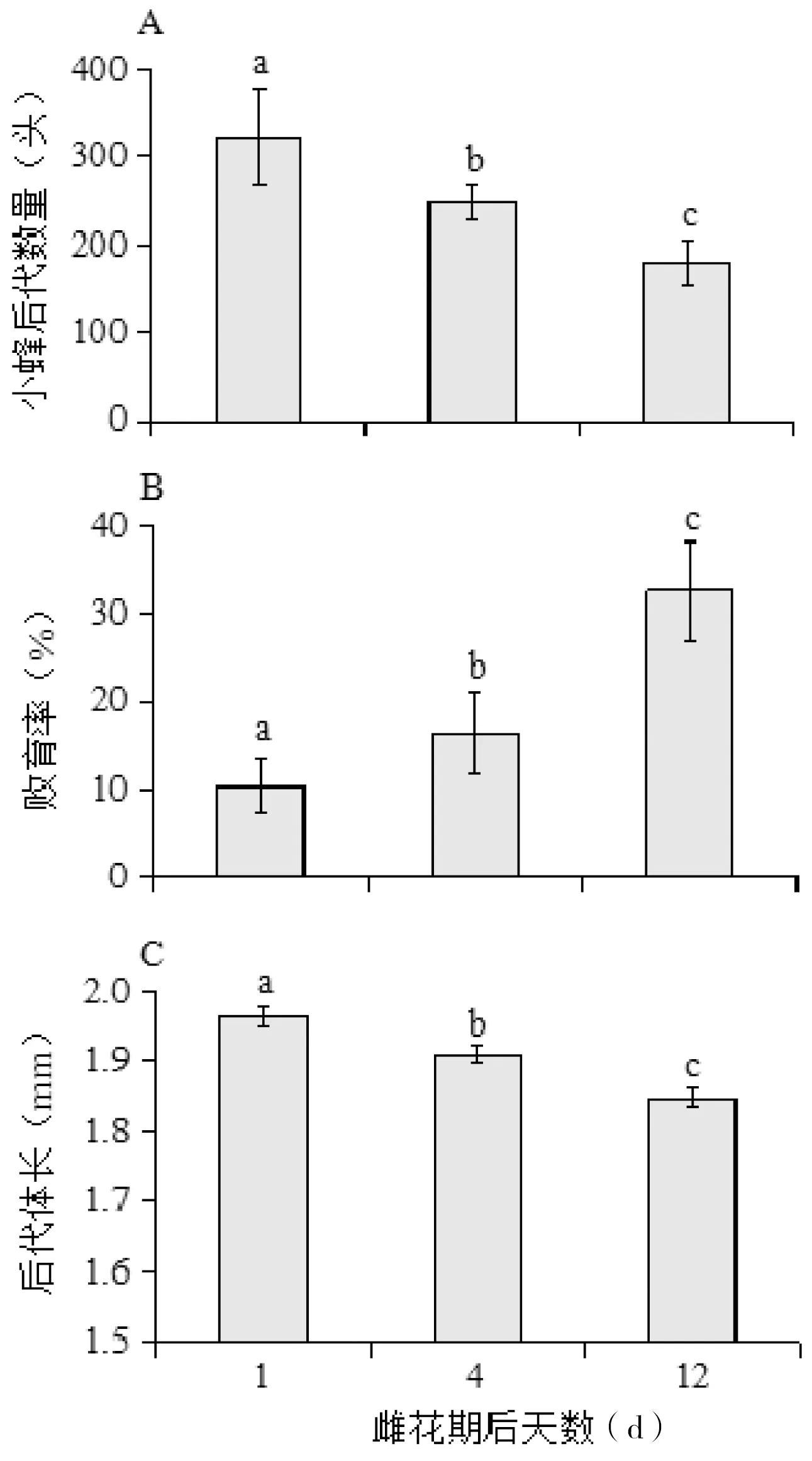
图3 鸡嗉子榕雌花期延长对传粉小蜂后代数量(A)、败育率(B)及后代体长(C)的影响
Fig 3 The impact of season on total number (A),abortion rate (B) andbody size (C) of pollinating fig wasps offspring inFicussemicordata
3 讨论
在热带亚热带地区,榕属植物的密度比较大,但就单种榕树来说,密度比较低,加之榕-蜂之间这种严格的一对一的传粉和寄生关系,以及小蜂短暂的寿命,有99%的小蜂都会在寻找到合适榕果前死去[17],所以小蜂要找到合适时期的榕果并非易事[25],而榕树-榕小蜂系统这种严格一对一的协同进化关系使得任何一方的繁殖失败都会导致系统的崩溃,所以对于榕-蜂系统来说,传粉小蜂资源是榕树和榕小蜂实现繁殖稳定的限制因子。因此,研究传粉榕小蜂繁殖的影响因素,是了解榕-蜂繁殖系统稳定的必要条件。
我们的实验针对自然界可能影响传粉小蜂繁殖的因素,采用人工控制性放蜂实验分别验证了季节变化、母代雌蜂年龄以及榕果所处时期对传粉小蜂后代数量,后代败育率以及后代体大小的影响。结果显示:在母代雌蜂数相同的情况下,季节变化对小蜂后代数量没有显著影响,虽然在西双版纳地区,干热季、雨季和雾凉季的水热条件不同,但由于榕果的间花期(即小蜂发育时间)也不同,在水热条件较差的雾凉季,榕果发育时间较长,所以弥补了水热条件的不足,完成了相同的有效积温,故对小蜂的死亡率也无显著影响。我们之前在聚果榕上进行的研究表明:在自然条件下,不同季节单果小蜂后代数量存在显著差异[19],但本次实验是人工控制性放蜂实验,不同于自然进峰的条件的后代数量,说明在自然条件下,小蜂群落存在的数量变化,并不是由于母代雌蜂的产卵数量造成的,而可能是榕果的物候对其进行调节[26]。另外,随着母代雌蜂年龄的延长以及榕果雌花期的延长,小蜂后代数量均呈现显著下降,在自然界,母代雌蜂进果时的年龄以及榕果时期都会存在不同[27]。这种延长虽然会降低小蜂后代的繁殖数量,但无疑也增加了榕-蜂相遇的概率,所以这种降低可以看作是对系统维持的一个权衡。
后代数量是反映榕果内小蜂繁殖的一个指标,而败育率反映了母代存活后代数量与产卵量的一个比值,除了可知道母代的产卵数量,还可以知道卵的质量或榕果质量是否适合小蜂后代发育。不同季节对小蜂败育率无影响,说明在不同季节,榕果间的质量并无显著差异,均适合小蜂发育,水热条件的不同也不会影响小蜂后代发育。自然条件下不同季节单果繁殖的差异可能只是进蜂量的差异造成的[19,26],而传粉母代寿命和榕果时期的延长都会导致小蜂后代的败育率增加,前者可能是因为卵质量的下降,后者是因为榕果质量下降造成的。
小蜂后代体长同样是反映小蜂繁殖状况的一个重要指标,体型较小的小蜂,寿命较短,飞行距离也随之缩短[28],进而影响榕-蜂系统的维持和稳定。我们的实验证明,在3个季节,雨季和雾凉季的小蜂后代体大小无差异,但要小于干热季,说明水热条件可能在某种程度上影响小蜂后代的大小,前期在聚果榕上的研究表明,季节对榕果大小会产生影响,同样是干热季的榕果较大,这与本次得到的小蜂体大小的规律一致。
人工控制性放蜂实验从自然界可能有变化的3个方面研究了其对传粉榕小蜂繁殖的影响,便于认识在自然条件下,榕-蜂繁殖系统是如何随着外界因素的改变而改变,这一结果有利于进一步了解这一互利共生系统的繁殖维持和稳定机制。
[1]WEST S A,GRIFFIN A,GARDNER A.Social semantics: altruism,cooperation,mutualism,strong reciprocity and group selection [J].J Evol Biol,2007,20(2): 415-432.
[2]WEIBLEN G D.How to be a fig wasp [J].Annu Rev Entomol,2002,47: 229-230.
[3]CRUAUD A,R NSTED N,CHANTARASUWAN B,et al.An extreme case of plant-insect co-diversification: figs and fig-pollinating wasps [J].Syst Biol,2012,61(6): 1029-1047.
[4]JANZEN D H.How to be a fig [J].Ann R Ecol,1979,10: 13-51.
[5]XIAO J H,YUE Z,JIA L Y,et al.Obligate mutualism within a host drives the extreme specialization of a fig wasp genome [J].Genome Biol,2013,14(12): R141.
[6]YU H,NASON J D.Nuclear and chloroplast DNA phylogeography ofFicushirta: obligate pollination mutualism and constraints on range expansion in response to climate change [J].New Phytol,2013,197(1): 276-289.
[7]ANSTETT M C.Unbeatable strategy,constraint and coevolution,or how to resolve evolutionary conflicts: the case of the fig/wasp mutualism [J].Oikos,2001,95(3): 476-484.
[8]SHANAHAN M,SO S,COMPTON S G,et al.Fig-eating by vertebrate frugivores: a global review [J].Biol Rev Camb Philos Soc,2001,76(4): 529-572.
[9]SCHATZ B,KJELLBERG F,NYAWA S,et al.Fig wasps: a staple food for ants onFicus[J].Biotropica,2008,40(2): 190-195.
[10]许再富.榕树—滇南热带雨林生态系统中的一类关键植物[J].生物多样性,1994,2(1): 21-23.
[11]LAMBERT F R,MARSHALL A G.Keystone characteristics of birds-dispersedFicusin a Malaysian lowland rain forest [J].J Ecol,1991,79(3): 793-809.
[12]LIU M,ZHANG J,CHEN Y,et al.Contrasting genetic responses to population fragmentation in a coevolving fig and fig wasp across a mainland island archipelago[J].Mol Ecol,2013,22(17): 4384-4396.
[13]MAWDSLEY N A,COMPTON S G,WHITTAKER R J.Population persistence,pollination mutualisms,and figs in fragmented tropical landscapes [J].Conserv Biol,1998,12(6): 1416-1420.
[14]HERRE E A,JAND R K C,MACHADO C A.Evolutionary ecology of figs and their associates: recent progress and outstanding puzzles [J].Annu Rev Ecol Evol Syst,2008,39: 439-458.
[15]PROFFIT M,SCHATZ B,BESSI RE J M,et al.Signalling receptivity: comparison of the emission of volatile compounds by figs ofFicushispidabefore,during and after the phase of receptivity to pollinators [J].Symbiosis,2008,45(1): 15-24.
[16]GU D,YANG D R.Utilisation of chemical signals by inquiline wasps in entering their host figs [J].J Insect Physiol,2013,59(10): 1065-1068.
[17]HERRE E A.Coevolution of reproductive characteristics in 12 species of New World figs and their pollinator wasps [J].Experientia,1989,45(7): 637-647.
[18]张 媛,杨大荣,彭艳琼.高榕的三类雌花形态及其繁殖特征[J].植物生态学报,2010,34(11): 1303-1309.
[19]张 媛,杨大荣,陈欢欢,等.季节变化对聚果榕-榕小蜂互利共生系统生长与繁殖的影响[J].广西植物,2015,35(4): 500-506.
[20]PENG Y Q,COMPTON S G,YANG D R.The reproductive success ofFicusaltissimaand its pollinator in a strongly seasonal environment: Xishuangbanna,Southwestern China [J].Plant Ecol,2010,209(2): 227-236.
[21]ZHANG Y,PENG Y Q,COMPTON S G,et al.Premature attraction of pollinators to inaccessible figs ofFicusaltissima: a search for ecological and evolutionary consequences [J].PloS ONE,2014,9(1): e86735.
[22]刘文杰,李红梅.西双版纳旅游气候资源[J].自然资源,1997,19(2): 62-66.
[23]赵俊斌,张一平,宋富强,等.西双版纳热带植物园引种植物物候特征比较[J].植物学报,2009,44(4): 464-472.
[24]杨成云,王瑞武,赵桂仿,等.[J].动物学研究,2005,26(4): 379-385.
[25]WARE A B,COMPTON S G.Dispersal of adult female fig wasps: 1.arrivals and departures [J].Entomol Exp Appl,1994,73(3): 221-229.
[26]PATEL A.Variation in a mutualism: phenology and the maintenance of gynodioecy in two Indian fig species[J].J Ecol,1996,84(5): 667-680.
[27]ZHANG Y,YANG D R,PENG Y Q,et al.Costs of inflorescence longevity for an Asian fig tree and its pollinator [J].Evol Ecol,2012,26(3): 513-527.
[28]DUNN D W,YU D W,RIDLEY J,et al.Longevity,early emergence and body size in a pollinating fig wasp-implications for stability in a fig pollinator mutualism [J].J Anim Ecol,2008,77(5): 927-935.
Research on effecting factors for reproduction and survivabilityof pollinating fig wasps in fig-fig wasps mutualism
ZHANG Yuan1,DENG Guo-bin1,LI Zong-bo2
(1.Yunnan Academy of Biodiversity;2.Key Laboratory of Forest Disaster Warning and Controlin Yunnan Province,College of Forestry,Southwest Forestry University,Kunming 650224,China)
The interaction between pollinating fig wasps (Agaonidae) and their host fig trees (Ficus) is a striking example of an obligate pollination mutualism.Agaonidae resource is the key factor which hinges fig-fig wasps reproduction.To test the influences of seasonal change,length of female phase and foundress age on the wasps offspring numbers,abortion ratio and body length of wasps offspring,we performed manually-controlling experiments on two importantFicusspecies distributed in Xishuangbanna,FicussemicordataandFicusaltissima.The experimental results revealed that seasonal change had no significant influence on wasps offspring numbers and abortion ratio,while it had significant impact on offspring body size.Furthermore,prominent negative influences can be found either along with the increase in age of foundress,or with the extension of female phase of figs,such as lower offspring numbers,higher abortive percentage and shorter body length of offspring.
mutualism; pollinating fig wasps; reproductive efficiency; female phase; cluth size
2016-01-04;
2016-02-01
收稿日期:国家自然科学基金项目(31560116);云南省应用基础研究计划项目(2013FD023);西南林业大学科研启动基金项目
张媛,博士,助理研究员,主要从事进化生态学研究,E-mail: zygogo@163.com
邓国宾,博士,高级实验师,主要从事植物学相关研究,E-mail: dqw1230@163.com
10.3969/j.issn.2095-1736.2016.06.043
Q948.12+2.5
A
2095-1736(2016)06-0043-05
猜你喜欢
中国科学探险(2022年8期)2022-12-31
西北农业学报(2022年7期)2022-07-22
疯狂英语·新悦读(2022年12期)2022-03-08
广西林业科学(2021年5期)2021-11-05
环境昆虫学报(2021年4期)2021-08-13
昆虫学报(2021年5期)2021-07-07
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
浙江大学学报(农业与生命科学版)(2019年4期)2019-09-19
华南农业大学学报(2015年5期)2015-12-04
生物灾害科学(2015年3期)2015-03-11
