
红椿不同种源种子性状的地理变异
2016-12-20 09:08邓小梅陈晓阳曾昭佳鲁好君
中南林业科技大学学报 2016年5期
陈 叶 ,湛 欣 ,邓小梅 ,陈晓阳 ,曾昭佳 ,鲁好君
(1.华南农业大学 广东省森林植物种质创新与利用重点实验室,广东 广州 510642;2.江西环境工程职业学院,江西 赣州 341000; 3. 广东九连山林场,广东 连平 517100)
红椿不同种源种子性状的地理变异
陈 叶1,2,湛 欣1,邓小梅1,陈晓阳1,曾昭佳3,鲁好君1
(1.华南农业大学 广东省森林植物种质创新与利用重点实验室,广东 广州 510642;2.江西环境工程职业学院,江西 赣州 341000; 3. 广东九连山林场,广东 连平 517100)
为揭示红椿不同种源种子性状地理变异规律,对产自31个不同地区红椿种子的6个种子性状进行观察测定并分析,结果表明:种子成熟期及色泽在不同种源间存在多样性;种长、翅1长、翅2长、翅1宽、翅2宽、千粒重这6个种子性状在种源间差异均达到极显著水平;经、纬度是影响种子性状的主要因素,种子长度、宽度、千粒重呈现出由南向北、由西向东逐渐增大的规律,除千粒重外其它种子性状均与海拔呈显著或极显著负相关,千粒重与无霜期呈显著负相关,除翅2宽外其它种子性状均与年降水量呈显著正相关;根据6个种子性状数据,对红椿31个种源进行聚类,基本可划分为3大类群。
红椿;种子性状;地理变异;种源
红椿Toona cliata,又名红楝子,属楝科Meliaceae香椿属Toona落叶或半落叶乔木,被国家列为Ⅲ级重点保护珍贵树种[1],素有“中国桃花心木”之美称,是我国热带、亚热带地区的珍贵速生用材树种[2],国内主要分布于福建、湖南、广东、广西、四川和云南等省区[3]。红椿是我国南方低山地区重要速生用材树种,同时作为营造针阔混交林进行树种结构调整的首选树种之一[4],大力推广和发展这一珍贵优良阔叶树种,对于发展林业生产有着重要意义[5]。自20世纪70年代发现红椿以来,研究人员对其进行了大量研究,目前国内外对红椿的研究主要集中在其生物学特性、繁殖育苗、栽培管理、资源调查以及开发利用等方面[6-12],对该树种种子性状地理变异的研究尚未见报道。
植物种子性状是较稳定的遗传特征,采用遗传上较为稳定、不易受环境影响的性状研究表型变异,可以揭示种源的遗传规律及变异大小[13]。林木种内地理变异是普遍的客观现象,种子性状的变异是对这种复杂环境的一种适应[14]。红椿分布广泛,可形成大量不同的遗传类型,通过对红椿种子性状地理变异进行研究,揭示红椿种子性状的变异程度及地理变异规律,可为红椿的遗传改良及种源区划提供参考
1 材料与方法
1.1 试验材料
2011年5月至2011年12月在红椿主要自然分布区的10个省31个采种点采集种子用于试验,采种时选择胸径25 cm以上的大树,采种母树间距在50 m以上,单株采种种子不少于100 g。种子自然晾干、清理干净后,分种源混合处理,密封置于4 ℃冰箱贮藏备用。采种点地理位置及生态因子见表1。

表1 红椿31个种源的地理位置和生态因子Table 1 Locations and ecological factors of 31 provenances in Toona ciliata
1.2 性状测定
参照刘军等[13]、岳华峰等[15]对毛红椿、苦槠的表型测定方法进行红椿种子性状测量。每种源随机抽取30粒种子,重复3次,用游标卡尺分别测定种子总长度、翅1(种翅较长的一侧)长、翅2(种翅较短的一侧)长、翅1宽(种翅最宽处)、翅2宽,测量单位精确到0.01 mm。千粒重测定采用千粒重法进行测定,每种源随即抽取1 000粒完整种子称量,重复3次,测量精确到0.01 g。
1.3 数据处理
采用Excel2003及SPSS17.0软件对试验数据进行统计分析。
2 结果与分析
2.1 红椿种子成熟期、色泽多样性
种源收集时发现红椿种子成熟期存在多样性,表现在不同地理种源的差异性,如云南景洪关平种子成熟期为5月上旬,保山为6月上旬,而广西百色、广东云浮、贵州望谟等地为6月底至7月上旬,江西、湖南、湖北、安徽为11月份。红椿物候多样性也表现在同一地理位置出现的差异,如福建南平部分单株种子成熟期为10月份,部分单株成熟期在12月份;同是广东韶关的曲江县红椿种子成熟时间为7月中旬,而乐昌种子成熟期则在11月底(各种源种子成熟期见表2)。

表2 红椿种子成熟期Table 2 Seed maturity of Toona ciliata
红椿表型特征丰富,主要表现在果实大小、形状,种子大小、色泽、千粒重等。红椿不同种源的种子色泽有很大区别,初步将其分为灰白、浅棕、深棕、浅褐、深褐5种色泽。浙江临安种源为灰白色;浙江其它种源、江西和湖南种源以棕色为主,如浙江丽水、江西官山和井冈山种子为浅棕色,湖南常德、邵阳种源种子为深棕色;广东、广西、云南、贵州地区种源以褐色为主,如广东云浮、广西田林种源种子为浅褐色,云南普洱、贵州册亨种源种子为深褐色。
2.2 红椿种子性状变异
对红椿种子形态性状进行方差分析(见表3)表明:红椿种子的形态指标种长、翅1长、翅2长、翅1宽、翅2宽、千粒重在种源间的差异均达到极显著水平,说明红椿种子性状存在丰富的遗传变异。
种子形态和千粒重的统计结果表明,31个产地红椿的平均种长为19.46 mm,翅1长为8.21 mm,翅2长为4.55 mm,翅1宽为4.56 mm,翅2宽为3.75 mm,千粒重为4.40 g。6个种子性状变异系数的幅度为0.72% ~ 19.77%,其中种子长度的变化幅度大于宽度变化幅度,说明红椿种子长短变异大于宽度变异。相对极差的变化趋势与变异系数不一致,说明种子性状稳定性差。

表3 红椿不同种源种子性状方差分析†Table 3 Variation in seed traits and variance analysis among different provenances in Toona ciliata
由表4可知,种长较长的为安徽、湖南、浙江等华中、华东地区的种源,较小的为广西、云南和贵州等西南、华南地区种源,其中种长最长的湖南湘西种源是最短的贵州安龙种源的1.88倍。翅1长与千粒重最大的均为江西马头山种源,最小的均为贵州安龙种源,千粒重最大为最小的4倍之多。翅2长与翅2宽较大的是湖南和安徽地区种源,较小的是贵州和云南省的种源,翅2长最长的为安徽泾县种源,翅2宽最大的为广东乐昌种源,最小的均为贵州安龙种源。翅1宽较大的为浙江、江西、湖南省种源,最大的是广东乐昌种源,最小的是云南楚雄种源。红椿种子大小基本可概括为华中、华东地区种源的种子较大且重,西南、华南地区种源种子较小且轻,可见随着纬度和经度的减小,种子性状有变小的趋势。 对31个种源种子性状进行多重比较,结果见表4。

表4 红椿31个种源种子性状的表现及多重比较†Table 4 The mean value and standard deviation of traits of 31 provenances in Toona ciliata
2.3 红椿种子性状与地理、气候因子的相关性
种子形态指标在产地间的差异性在一定程度上反映不同生长环境对植物生长的影响。红椿种子性状与地理、生态因子间的相关分析见表5。结果表明,红椿4个种子性状均与经、纬度呈极显著正相关,种子长度、宽度、千粒质量呈现出由南向北、由西向东逐渐增大的规律。种子种长、翅1长、翅2长、翅1宽、翅2宽与海拔呈显著或极显著负相关,随着海拔的降低,种子有变大的趋势;除翅2宽外其它种子性状均与年降水量呈显著正相关,说明随着水分的增加,能够使种子更大且更饱满;千粒重还与无霜期显著负相关,无霜期越短,种子越饱满,如无霜期较短的江西马头山和湖南邵阳种源种子千粒重大。
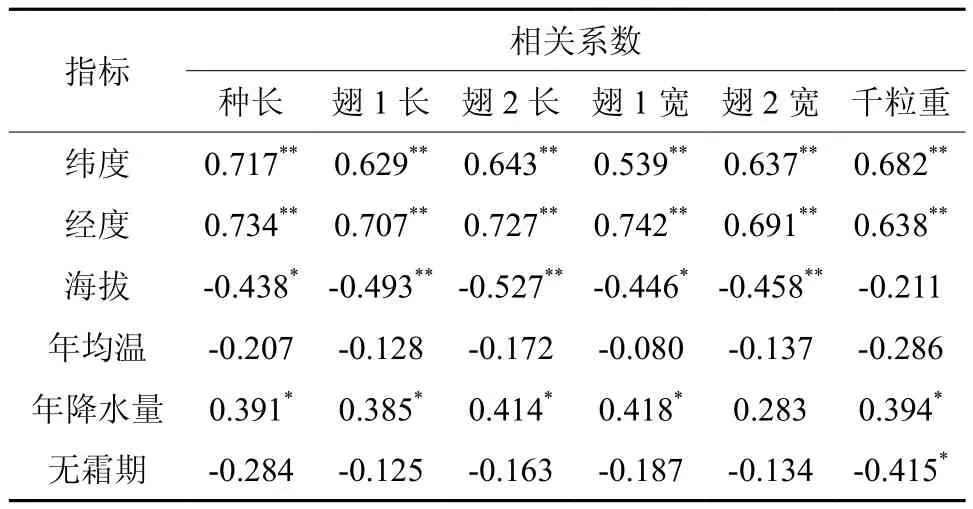
表5 红椿种子性状与地理生态因子间的相关分析Table 5 Correlative analysis between seedling traits and geographic climate factors in Toona ciliata
2.4 红椿不同种源种子性状的聚类分析
由于红椿各产地间差异大,因此通过种子性状进行聚类分析对于种源区划十分有必要。现根据6个种子性状数据,对红椿31个种源进行聚类(见图1),基本可将31个红椿种源划分为3大类群。第一大类属于西南、华南地区,如云南保山、广西隆林、贵州兴义等种源;第二类除湖南怀化、浙江临安、福建南平3个种源外也主要为西南、华南地区种源,其中湖南怀化地处湖南西部,临近贵州,福建南平年均温高,年降水量大,这些因素可能是造成这两个种源归属此类原因,浙江临安种源推测其可能从华南或西南地区引种而来;第三大类属于华中、华东地区,如湖南湘西、江西官山、湖北恩施、浙江丽水、安徽泾县等种源,据调查广东乐昌种源为原生种,其地处广东北部,临近湖南、江西,故其被划为此类。整体上看红椿种子性状变异基本随地理分布呈连续性变化,具有明显地理区域特征。

图1 红椿种子性状聚类分析Fig.1 Cluster diagram based on seed morphological traits in Toona ciliata
3 结论与讨论
3.1 不同种源的红椿种子性状变异
同一物种不同产地形态上的差异是遗传型和环境因子共同作用的结果,而且差异越大,可能存在的遗传变异也越大[16]。31个红椿种源的种子性状在不同产地间均存在极显著差异,说明红椿种源间存在极大的遗传变异,为优良红椿种源的选择提供了基础条件。这与国内对其它树种如乌桕、桤木等种子性状的种源变异研究基本类似[17-18]。翅宽的变异系数比翅长的小,说明种子纵向的变异程度小于横向的长短变异程度,6个种子性状的相对极差与变异系数的变化规律不一致,说明种子性状的稳定性差。西南部种源种子较小,东南部种源种子相对大一些,呈一定地理分布规律。
3.2 红椿种子性状地理变异规律
探讨种子性状之间的相关关系,可以用来指导选择育种和林木种质资源保存中样本策略的制定。通过对不同种源种子性状进行相关分析,利用各性状间的内在联系对容易测定的性状进行预测和间接选择,可以降低改良成本,提高选择效率[19]。红椿不同种子性状间存在极显著差异,说明可根据这6个种子性状进行种源选择。
通过对红椿种子性状与地理、生态因子的相关性分析,可以了解不同环境条件对红椿种子性状的影响。总体上看种子性状基本可概括为在采样分布区东北部的群体较大,西南部群体较小,即随着纬度的减小和经度的减小,种子性状有变小的趋势。相关分析表明,经纬度均与红椿种子性状呈显著正相关,这与苦槠种子形态性状与经纬度负相关不一致[15]。海拔与种子性状呈极显著负相关,说明海拔越高,种子性状越小,这一结论与刘军等[13]、舒枭等[20]对毛红椿和厚朴的变异研究不一致。
3.3 红椿种子性状聚类分析
通过聚类分析可将红椿31个种源划分为3大类,前两类主要为西南、华南地区种源,第三类主要为华中、华东地区种源,进一步分析可知湖南怀化、福建南平种源地理上属于东北部,而聚类上却划分为西南、华南地区一类,广东乐昌种源划分到华中、华东地区一类,这可能是由于生态因子所致,而浙江临安被划为华南、西南地区,猜测其是否由华南、西南地区引入,对此还需进一步证实。
[1]中国科学院植物研究所.中国植物红皮书——稀有濒危植物(一)[M].北京:科学出版社, 1992.
[2]中国树木志编委会.中国主要树种造林技术[M].北京:中国林业出版社, 1981: 613-615.
[3]中国科学院中国植物志编辑委员会.中国植物志:第43卷[M].北京:科学出版社, 1997:40.
[4]马献良.红椿播种育苗初步研究[J].安徽林业科技, 2005, (4):7-8.
[5]程冬生,崔同林.珍贵速生树种红椿的利用价值及栽培技术[J].中国林副特产,2010, (4):39-40.
[6]赵汝玉,李光友,徐建民.红椿育苗及造林技术[J].广西林业科学,2005, 34(3):155-156.
[7]吴际友,程 勇,王旭军,等.红椿无性系嫩枝扦插繁殖试验[J].湖南林业科技,2011,38(4) :5-7.
[8]卢海啸,李家洲,莫花浓,等.红楝子提取物对小鼠记忆和耐力的影响[J].中国野生植物资源, 2008,27(4):55-58.
[9]李家洲,卢海啸,吴华县.红椿不同提取物药理活性初探[J].安徽农业科学, 2009, 37(29):14164-14166.
[10]梁瑞龙,廖仁雅,戴 俊.红椿濒危原因分析及保护策略[J].广西林业科学, 2011, 40(3):201-203.
[11]刘 球,陈彩霞,吴际友,等.红椿无性系幼苗叶片抗氧化酶指标对干旱胁迫的响应[J].中南林业科技大学学报,2013,33(11): 73-76.
[12]陈彩霞,王瑞辉,吴际友,等.持续干旱条件下红椿无性系幼苗的生理响应[J].中南林业科技大学学报,2013,33(8):46-49.
[13]刘 军,张海燕,姜景民,等.毛红椿种实和苗期生长性状地理种源变异[J].南京林业大学学报:自然科学版, 2011, 35(3):55-59.
[14]Shu X, Yang Z L, Yang X, et al. Variation in seed characters of Magnolia of fi cinalis from different locations[J]. Forest Research,2010, 23(3): 457-461.
[15]岳华峰,邵文豪, 井振华,等.苦槠种子形态性状的地理变异分析[J].林业科学研究, 2010, 23(3): 453-456.
[16]Roach D A, Wulff R D. Maternal effects in plants[J]. Annual Review of Ecology and Systematics, 1987, 18: 209-235.
[17]金雅琴.乌桕不同种源种子性状的地理变异[J].西南林业大学学报, 2011,31(4):44-48.
[18]王军辉,顾万春,万 军,等.桤木不同种源球果及种子性状的遗传变异[J].东北林业大学学报, 2006,34 (2): 1-4.
[19]魏志刚,高玉池,杨传平,等.引种盐松不同种源种子表型性状和发芽特性[J].东北林业大学学报, 2009,37(11):7-10.
[20]舒 枭,杨志玲,杨 旭,等.不同产地厚朴种子性状的变异分析[J].林业科学研究, 2010, 23(3): 457-461.
Geographic variation in seed traits of Toona cliata from different locations
CHEN Ye1,2, ZHAN Xin1, DENG Xiao-mei1, CHEN Xiao-yang1, ZENG Zhao-jia3, LU Hao-jun1
(1. Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Agricultural University of South China, Guangzhou 510642, Guangdong, China; 2. Jiangxi Environmental Engineering Vocational College, Ganzhou 341000, Jiangxi,China; 3. Guangdong Jiulianshan Forest Farm, Lianping 517100, Guangdong, China)
In order to investigate geographic variation in seed traits of Toona cliata, 6 seed morphologic characters were measured in 31 seed collect areas in natural distribution of China. Results show that extremely signi fi cant differences were found among different provenances in seed length, wing 1 length, wing 2 length, wing 1 width, wing 2 width and thousand seed weight. The latitude and longitude were the main factors that affected the size of seed. From southwest to northeast, the size of seed gradually increasing, There was signi fi cant negative correlation between all seed traits and elevation except thousand seed weight, The thousand seed weight was signi fi cant negative associated with frost-free period, There was signi fi cant positive correlation between all seed traits and altitude except wing 2 width. According to cluster analysis on 6 seed characters mean values including seed size and quality and geographical climatic factors, the Toona cliata from 27 natural distribution regions could be divided into 3 provenance region.
Toona cliata; seed traits; geographic variation; provenances
S722.3+1
A
1673-923X(2016)05-0052-06
10.14067/j.cnki.1673-923x.2016.05.010
2015-01-14
林业公益性行业科研专项项目(201004020)
陈 叶,硕士研究生 通讯作者:邓小梅,教授 博士;E-mail:dxmei2006@scau.edu.cn
陈 叶,湛 欣,邓小梅,等. 红椿不同种源种子性状的地理变异[J].中南林业科技大学学报,2016, 36(5): 52-57.
[本文编校:谢荣秀]
猜你喜欢
特产研究(2022年6期)2023-01-17
四川蚕业(2022年2期)2022-11-19
绿色科技(2021年21期)2021-11-26
辽宁林业科技(2021年1期)2021-03-16
趣味(数学)(2020年4期)2020-07-27
世界科学技术-中医药现代化(2020年2期)2020-07-25
四川蚕业(2020年3期)2020-07-16
支部建设(2020年15期)2020-07-08
农家科技中旬版(2020年4期)2020-07-06
百科知识(2015年18期)2015-09-10
