
胡麻粗脂肪含量的主基因+多基因遗传分析
2016-12-16 00:35:48化青春王利民张建平党占海
西北农林科技大学学报(自然科学版) 2016年11期
化青春,赵 利,王利民,赵 玮,党 照,张建平,党占海
(甘肃省农业科学院 a 小麦研究所,b 作物研究所,甘肃 兰州 730070)
胡麻粗脂肪含量的主基因+多基因遗传分析
化青春a,赵 利b,王利民b,赵 玮b,党 照b,张建平b,党占海b
(甘肃省农业科学院 a 小麦研究所,b 作物研究所,甘肃 兰州 730070)
【目的】 了解胡麻粗脂肪含量的遗传方式。【方法】 以用DYM×STS构建的包含233个家系的重组自交系群体(F6∶7)和2个亲本为材料,在甘肃定西、宁夏固原和河北张家口3个环境下种植,采用索氏抽提仪测定粗脂肪含量,运用数量性状主基因+多基因混合遗传模型分析方法,进行胡麻粗脂肪含量的遗传模型分析。【结果】 甘肃定西、宁夏固原和河北张家口3个环境条件下家系间胡麻粗脂肪含量均存在广泛变异,表现超亲遗传现象,近似正态分布;粗脂肪含量的遗传模型均为4MG-AI,表现为4对具有加性上位性效应的主基因遗传模型,不存在多基因效应。主基因遗传率分别为92.80%,99.02%和80.71%,环境引起的变异分别为7.20%,0.98%和19.29%。【结论】 胡麻粗脂肪含量受主基因控制,且存在加性上位性效应。在胡麻高油育种中,提高粗脂肪含量不仅要注重主基因的利用,也要考虑基因间的互作效应。
胡麻;粗脂肪;主基因+多基因遗传分析
胡麻(LinumusitatissimumL.)主要分布在我国的甘肃、山西、内蒙古、宁夏、河北和新疆等地区,是当地重要的特色油料作物和经济作物。甘肃省胡麻的种植面积约占全国的40%,总产量约占全国的42%,面积和总产量均居全国首位,对我国胡麻生产和科研起着举足轻重的作用。 随着社会发展和人民生活水平的提高,胡麻油的保健作用越来越受到重视,胡麻的市场需求量大幅上升,加强高产高油胡麻育种便成了育种家追求的重要目标。因此了解胡麻粗脂肪含量的遗传规律,进行胡麻粗脂肪含量的遗传分析尤为重要。盖钧镒等建立的主基因+多基因混合遗传模型常被作为植物数量性状遗传的通用模型[1-5]。这种分析方法不仅能够评估植物基因的总体遗传效应,还可以分析主基因与多基因是否存在及其相应的效应值,使数量性状的表型特征得到较全面的描述,在一定程度上提高了对植物复杂数量性状遗传规律的认识,并可指导育种研究[6-8]。该方法已在玉米[9-10]、向日葵[11]、大豆[12-16]、花生[6-8,17-20]、甘蓝型油菜[21-23]、棉花[24-25]、小麦[26]、白菜[27]、甜瓜[28]、甘蓝[29]等作物的遗传研究中得到了广泛的应用。但在胡麻粗脂肪含量遗传研究上,目前仅有通过遗传力来评价含油率遗传的报道[30-31],而通过数量性状的主基因+多基因混合模型来研究胡麻粗脂肪含量遗传规律的研究尚未见报道。因此,本研究拟以DYM×STS构建的重组自交系群体为材料,在甘肃定西(E1)、宁夏固原(E2)和河北张家口(E3)3个环境下种植,采用索氏抽提仪测定胡麻粗脂肪含量,运用数量性状的主基因+多基因混合遗传模型分析方法,以确定胡麻粗脂肪含量的最适遗传模型和基因作用方式,估计基因的遗传效应大小,揭示基因型效应和环境效应对胡麻粗脂肪含量的影响,为胡麻高油育种提供理论参考。
1 材料与方法
1.1 试验材料
2011年夏季以综合农艺性状好、含油率较低的DYM为母本(P1),以含油率高的STS为父本(P2),在甘肃兰州配制杂交组合,当年冬季在云南元谋种植F1,次年夏季于甘肃兰州种植F2,F2单株通过一粒传的方法经兰州、元谋一年两季连续多代自交,于2014年形成包含233个家系的重组自交系(RIL)群体(F6∶7)。
1.2 试验方法
1.2.1 材料种植 试验采用完全随机区组设计,2次重复,2个亲本及233个家系于2014年春季在甘肃定西、宁夏固原和河北张家口3个环境下种植,RIL家系每系1行,行长2 m,行距20 cm,栽培管理同常规生产大田。
1.2.2 粗脂肪含量测定 成熟时按家系收获,自然晾晒、风干后测定各个家系和亲本的粗脂肪含量,每个样品重复2次。粗脂肪含量采用索氏抽提仪测定,执行检测标准GB/T 14772-1993。
1.3 数据分析

2 结果与分析
2.1 胡麻2个亲本及RIL群体粗脂肪含量的遗传变异
由表1可以看出,在3个不同环境E1、E2和 E3条件下,RIL群体粗脂肪含量均存在一定程度的变异。在E1环境下变幅为34.56%~47.50%,在E2环境下变幅为34.61%~46.60%,在E3环境下变幅为35.83%~46.81%,群体的极值分别为 12.94%,11.99%和10.98%,均出现了粗脂肪含量超低亲和超高亲变异类型,且极值差别较大,表明该RIL群体家系间变异类型比较丰富,适合进行下一步的遗传模型分析。
在E1、E2 和 E3环境中胡麻粗脂肪含量变异系数较为接近,分别是5.06%,4.53%和3.98%,偏度、峰度绝对值均小于或接近1,近似正态分布。对其进行Kolmogorov-Smirnov正态性检验,Sig.值分别为0.044,0.045和0.041,表明该频次分布为偏态分布,由图1~3也可以看出此现象,说明存在主基因控制胡麻粗脂肪含量的情况。

表 1 亲本和RIL群体胡麻粗脂肪含量的变异参数
注:E1.甘肃定西;E2.宁夏固原;E3.河北张家口。下表同。
Note:E1.Dingxi Gansu;E2.Guyuan Ningxia;E3.Zhangjiakou Hebei.The same below.
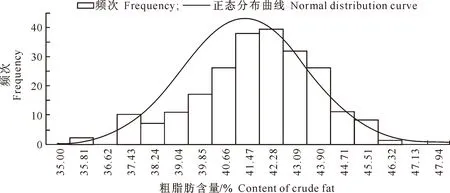
图 1 甘肃定西胡麻粗脂肪含量频次分布

图 2 宁夏固原胡麻粗脂肪含量频次分布
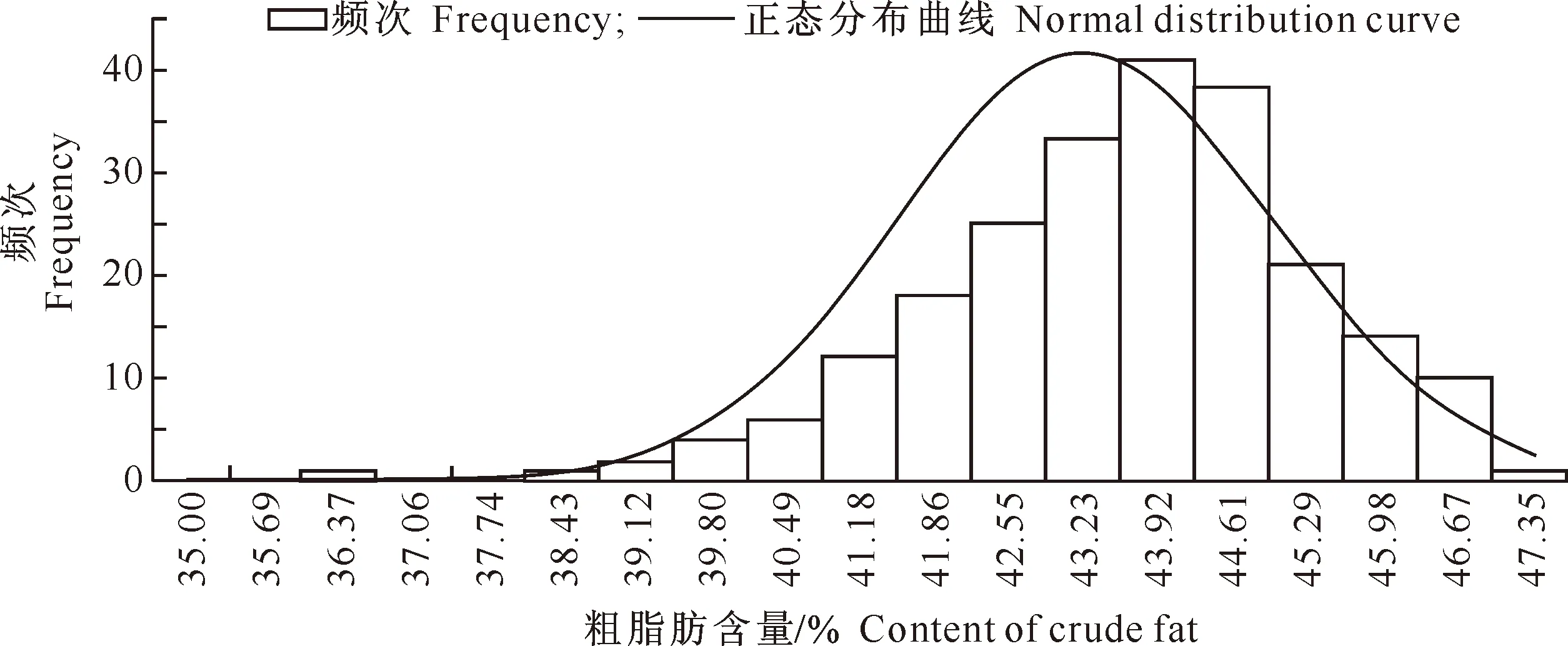
图 3 河北张家口胡麻粗脂肪含量频次分布
2.2 最适遗传模型分析
经遗传模型分析软件G3DH.exe[32]运算,得到不同遗传模型及其AIC值及适合性检验值见表2。根据最佳遗传模型选择的原则,在甘肃定西环境中AIC值最小的3个模型分别为4MG-AI、MX3-AI-AI和3MG-A,统计量达到显著水平的个数分别为1,0和3,模型MX3-AI-AI达到显著水平的个数最少,但模型4MG-AI的AIC值最小,统计量达到显著水平的个数仅为1,因此确定该试验点的最优模型为4MG-AI。在宁夏固原环境中AIC值最小的3个模型分别为4MG-AI、MX2-AI-A和MX3-AI-AI,统计量达到显著水平的个数均为0个,因此直接选取AIC值最小的4MG-AI模型为该试验点的最优模型,即4对主基因遗传模型,无多基因效应,但主基因存在加性上位性效应。在河北张家口环境中AIC值最小的3个模型分别为MX3-AI-AI、4MG-AI和3MG-A,统计量达到显著水平的个数分别为8,7和11个,可见模型4MG-AI达到显著水平的个数最少,表明该模型能够对该环境中胡麻粗脂肪含量的遗传特性作出充分解释,可以作为该试验点胡麻粗脂肪含量遗传的最优模型。由于甘肃定西试验、宁夏固原试验和河北张家口试验的最优模型均为4MG-AI,说明胡麻粗脂肪含量的最优遗传模型为4对具有加性上位性效应的主基因遗传模型,无多基因效应。
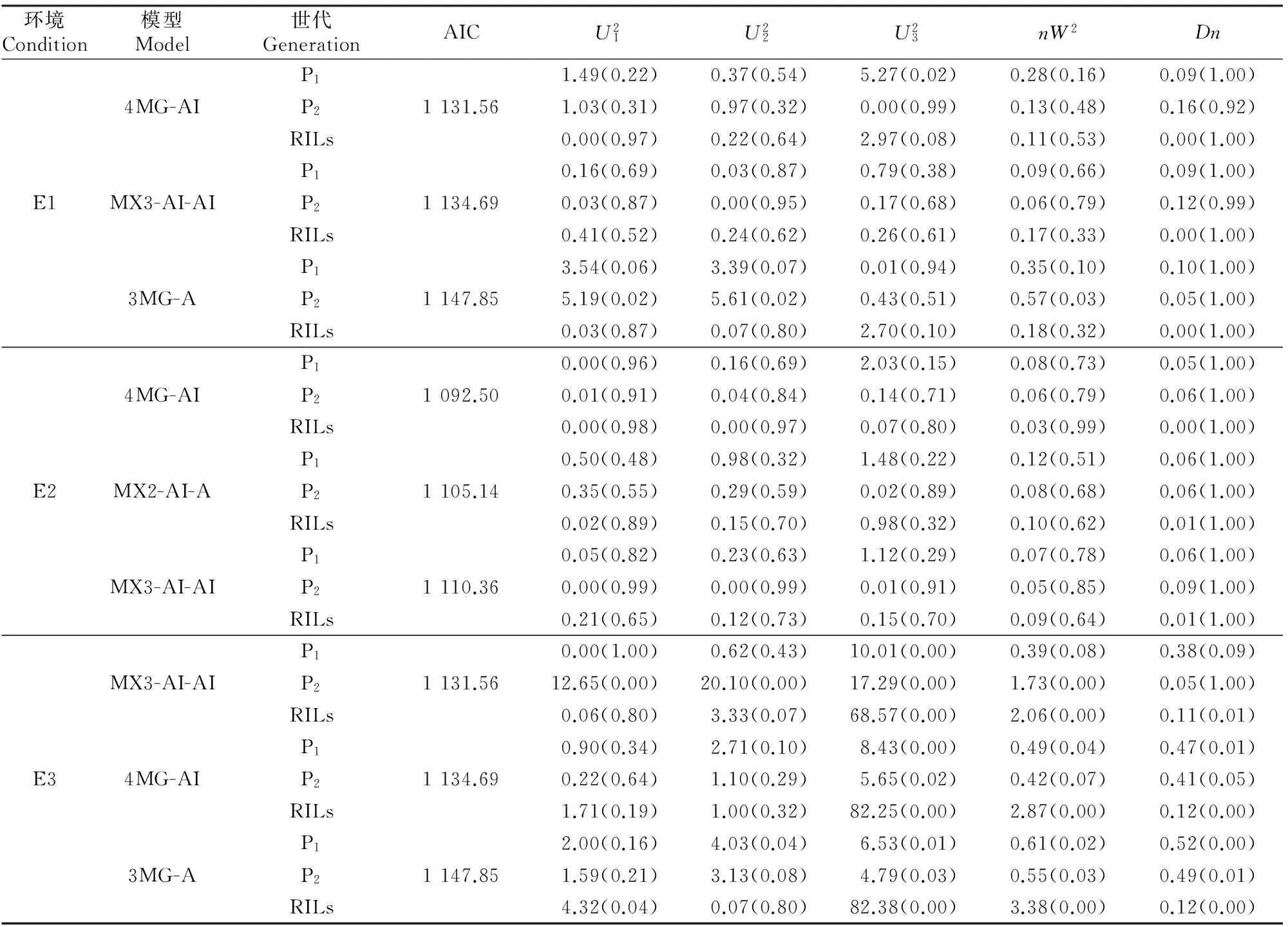
表 2 胡麻RIL群体粗脂肪含量候选模型的AIC值及适合性检验

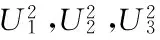
2.3 遗传参数估计
由表3可以看出,在E1环境中,胡麻粗脂肪含量4个主基因的加性效应值分别为-1.84,-2.65,4.37和1.12,4对主基因的作用不等,因|dc|>|db|>|da|>|dd|,说明第3对主基因的加性作用最大,第4对主基因的加性作用最小。4对主基因的加性效应中虽然有2对可以增强胡麻粗脂肪含量的正向效应和2对减弱胡麻粗脂肪含量的负向效应,但由于正向效应相对较大,所以加性效应能使胡麻粗脂肪含量小幅增加;4对基因间两两上位性互作效应分别为-6.56,4.94,-2.64,4.95,-3.15和 5.16,表现为3对正向效应和3对负向效应,但由于正向效应较大,能使胡麻粗脂肪含量增加,因此主基因遗传率为92.80%,仅有7.20%的变异由环境因素决定。同时,|da|+|db|+|dc|+|dd|<|iab|+|iac|+|iad|+|ibc|+|ibd|+|icd|,说明在这个试验点,粗脂肪含量的上位性效应要大于加性效应。
由表3可以看出,在E2环境中,胡麻粗脂肪含量4个主基因加性效应值分别为-3.54,-4.37,5.15和2.78,说明4对主基因的作用不等,因|dc|>|db|>|da|>|dd|,说明第3对主基因的加性作用最大,第4对主基因的加性作用最小。其中dc和dd为正向效应,可以使胡麻粗脂肪含量增加,da和db为负向效应,可以使胡麻粗脂肪含量减弱,正负效应基本抵消了粗脂肪含量的增加;4对基因间两两上位性互作效应分别为-3.61,4.48,2.68,4.95,2.77和-3.81,有4对可以使粗脂肪含量增加的正向效应和2对减弱粗脂肪含量的负向效应,互作效应使粗脂肪含量得到大幅增加,主基因遗传率为99.02%,属于高等遗传率。表明控制粗脂肪含量的遗传是主基因作用的结果,仅有0.98%由环境因素决定。同时,|da|+|db|+|dc|+|dd|<|iab|+|iac|+|iad|+|ibc|+|ibd|+|icd|,说明在这个试验点,胡麻粗脂肪含量的上位性效应要大于加性效应。
由表3还可以看出,在E3环境中,4个主基因的加性效应值分别为5.75,5.75,3.81和4.54,|da|=|db|>|dd|>|dc|,说明第1对和第2对主基因的加性作用相等且均大于第4对和第3对的加性效应,4对主基因的加性效应均为正向效应,都能使胡麻粗脂肪含量增加, 4对主基因间两两上位性互作效应分别为-0.76,-1.33,-5.41,-1.33,-5.03和-3.47,互作效应均为负效应,降低了胡麻粗脂肪含量,因此主基因遗传率仅为80.71%,属于中等偏上的遗传率,有19.29%的变异由环境因素决定。因该点|da|+|db|+|dc|+|dd|>|iab|+|iac|+|iad|+|ibc|+|ibd|+|icd|,说明该试验点胡麻粗脂肪含量的加性效应要大于上位性效应。
由于胡麻RIL群体在3个试验点中主基因效应决定了胡麻粗脂肪含量表型变异的92.80%,99.02%和80.71%,仅有7.20%,0.98%和 19.29% 是由环境因素决定,主基因的遗传力非常高,即胡麻粗脂肪含量主要由遗传效应控制,环境效应对其影响不大。3个试验点间的主基因加性效应和主基因间的上位性互作效应差异较大,也说明了胡麻粗脂肪含量的数量性状特征。
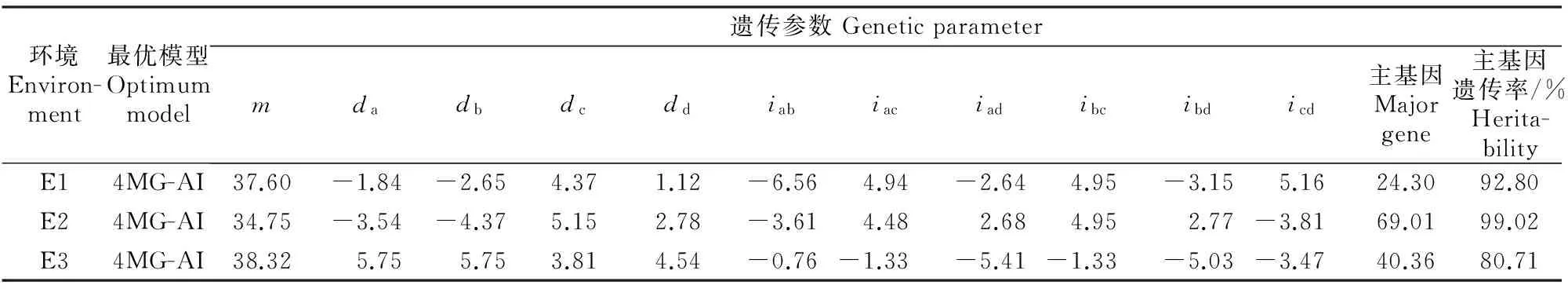
表 3 胡麻RIL群体的粗脂肪含量最优模型及遗传参数估计
注:m.群体均值;d.主基因加性效应值;a、b、c、d.主基因序号;ab、ac、ad、bc、bd、cd.分别为主基因之间的互作;i.上位效应值。
Note:m.Population mean;d.Additive effect in major gene;a,b,c,d.Number of major-gene additive effect of the major gene;ab,ac,ad,bc,bd,cd.Interaction between major genes;i.Epistasis value.
3 讨 论
粗脂肪是胡麻的重要品质性状,研究粗脂肪含量的遗传规律是胡麻品质育种的必要前提,同时也可为合理的世代选择提供理论依据。本研究以DYM×STS获得的F6∶7重组自交系群体RIL为研究对象,研究胡麻粗脂肪的遗传。由于RIL 家系间是纯合稳定的,无显性和显性互作的干扰,且受环境影响较小,因此能较准确地估计主效基因的加性效应值、加性和加性互作效应值[26],适合进行数量性状遗传分析。本研究结果表明,胡麻粗脂肪含量在3个不同地区的最适模型相同,均为4MG-AI,表现为4 对具有加性上位性效应的主基因遗传模型,且主基因遗传率均在80%以上,均较大,说明内在主基因之间的互补作用或加性作用对胡麻粗脂肪含量变异的贡献率高,在宁夏固原和甘肃定西试验点上位性效应大于加性效应,而在河北张家口试验点加性效应大于上位性效应,而非遗传因素如环境等对其影响较小,说明胡麻粗脂肪含量在不同环境中的遗传方式相对较稳定。
包海柱[11]研究认为,向日葵含油率受加性、显性和上位性遗传效应的控制,加性效应较显性效应重要。宋启建等[12]研究认为,大豆油脂含量主要受基因加性效应控制,并存在一定的显性和上位性效应。赵刚等[9-10]研究认为,玉米籽粒含油率符合2对主基因+多基因遗传模型。郑永战等[13]研究认为,大豆脂肪含量受2对加性互补主基因+多基因控制;王永军[14]研究表明,大豆油分的遗传属于2对连锁主基因+多基因的模型;郭志刚[15]研究认为,大豆油脂的遗传为1对主基因+多基因或多基因遗传模型;刘顺湖等[16]研究认为,大豆油脂含量为2~3对主基因+多基因模型。张洁夫等[21]认为,甘蓝型油菜种子含油量由1对加性-显性主基因+加性显性-上位性多基因控制。刘华等[20]研究认为,花生脂肪含量的遗传受多基因加性遗传和环境作用。陈四龙等[7]认为,花生含油量基因遗传模型存在较大差异,既有多基因加性遗传模型,也有主基因+多基因遗传模型。上述在不同作物中的研究均认为,含油率属于主基因+多基因遗传模型,而本研究认为3个试验点中胡麻粗脂肪含量均为主基因遗传,没有多基因效应,与前人在玉米、向日葵、大豆、甘蓝型油菜和花生等作物含油率上的研究结果不同,这可能与各研究采用的试验材料、分析方法以及不同环境有关。
虽然RIL群体的胡麻粗脂肪含量主要受遗传控制,但控制粗脂肪含量的遗传因素比较复杂,如在甘肃定西和宁夏固原环境中4个主基因的加性效应值均为2负2正,而在河北张家口环境中4个主基因的加性效应值均为正值,两基因间的互作效应值3个点分别为全负、4正2负和3负3正,从而导致不同试验点主基因遗传率差别较大。同时4个主基因两两间的上位性互作效应在不同地点表现不同,宁夏固原点和甘肃定西点的上位性效应要大于加性效应,而河北张家口点则相反,为加性效应大于互作效应。这些充分说明胡麻粗脂肪含量遗传的复杂性及其数量性状特征。由于胡麻粗脂肪含量受4对主基因控制,且主基因遗传率较高,说明可能含有较大的主效QTL,因此在胡麻高油育种中,提高脂肪含量要注重主基因的利用,重视主效基因的聚合,同时要重视基因间的互作效应。可以通过粗脂肪含量高的品种(系)杂交、回交和系统选择等方法对性状进行遗传改良;或者通过基因组测序数据构建胡麻遗传连锁图谱,通过QTL 基因定位寻找与这些基因紧密连锁的分子标记,通过分子标记辅助选择来加速育种进程。
[1] 盖钧镒,章元明,王建康.QTL混合遗传模型扩展至2 对主基因+多基因时的多世代联合分析 [J].作物学报,2000,26(4):385-391.
Gai J Y,Zhang Y M,Wang J K.A joint analysis of multiple generations for QTL models extended to mixed two major genes plus polygene [J].Acta Agronomica Sinica,2000,26(4):385-391.
[2] 章元明,盖钧镒.利用DH 或RIL 群体检测QTL 体系并估计其遗传效应 [J].遗传学报,2000,27(7):634-640.
Zhang Y M,Gai J Y.Idenitification of mixed major genes and polygenes inheritanee model of quantitative traits by using DH or RIL populaiton [J].Acta Genetica Sinica,2000,27(7):634-640.
[3] 盖钧镒,章元明,王建康.植物数量性状遗传体系 [M].北京: 科学出版社,2003:145-156.
Gai J Y,Zhang Y M,Wang J K.Genetic system of quantitative traits in plants [M].Beijing:Science Press,2003:145-156.
[4] Gai J Y,Wang J K.Identification and estimation of QTL model and effects [J].Theor Appl Genet,1998,97:1162-1168.
[5] Wang J K,Gai J Y.Mixed inheritance model for resistance to agromyzid beanfly(MelanagromyzasojaeZehntner) in soybean [J].Euphytica,2001,122:9-18.
[6] 张新友,韩锁义,徐 静,等.花生蛋白质含量的主基因+多基因遗传分析 [J].中国油料作物学报,2011,33(2):118-122.
Zhang X Y,Han S Y,Xu J,et al.Genetic analysis of protein using major gene plus polygene methods in peanut(ArachishypogaeaL.) [J].Chinese Journal of Oil Crop Science,2011,33(2):118-122.
[7] 陈四龙,李玉荣,程增书,等.花生含油量杂种优势表现及主基因+多基因遗传效应分析 [J].中国农业科学,2009,42(9):3048-3057.
Chen S L,Li Y R,Cheng Z S,et al.Heterosis and genetic analysis of oil content in peanut using mixed model of major gene and polygene [J].Scientia Agricultura Sinica,2009,42(9):3048-3057.
[8] 刘恩生.花生蛋白质、脂肪含量及其它农艺性状的配合力和遗传参数分析 [J].华北农学报,1987,2(3):18-26.
Liu E S.Diallel analysis for yield,protein and oil content of peanut [J].Acta Agriculturae Boreali-Sinica,1987,2(3):18-26.
[9] 赵 刚,昂 沃,郝小琴,等.微胚乳超高油玉米籽粒含油率的主基因+多基因遗传分析 [J].中国油料作物学报,2010,32(1):53-56.
Zhao G,Ang W,Hao X Q,et al.Major gene plus polygene inheritance analysis of seed oil content in microendosperm super-high oil corn [J].Chinese Journal of Oil Crop Science,2010,32(1):53-56.
[10] 赵 刚.微胚乳超高油玉米几个性状的主基因+多基因遗传研究 [D].南宁:广西大学,2007.
Zhao G.Stduies on the major gene plus polygene inheritance of several traits in microendosperm super-high oil corn [D].Nanning:Guangxi University,2007.
[11] 包海柱.基于混合线性模型的油用向日葵农艺性状及产质量遗传研究 [D].呼和浩特:内蒙古农业大学,2013.
Bao H Z.The genetic study on agronomic traits,yield and qualities traits of oil sunflower based on mixed linear model [D].Hohhot:Inner Mnngolia Agricultural University,2013.
[12] 宋启建,盖钧镒,马育华.大豆品种蛋白质、油分含量的遗传特点 [J].中国农业科学,1989,22(6):24-29.
Song Q J,Gai J Y,Ma Y H.A study on genetic property of protein and oil content in soybean [J].Scientia Agricultura Sinica,1989,22(6):24-29.
[13] 郑永战,盖钧镒,周瑞宝,等,大豆脂肪及脂肪酸组分含量的遗传分析 [J].大豆科学,2007,26(6):801-806.
Zheng Y Z,Gai J Y,Zhou R B,et al.Inheritance of fat and fatty acid composition contents in soybean [J].Soybean Science,2007,26(6):801-806.
[14] 王永军.大豆重组自交系群体的构建与调整及其在遗传作图、抗花叶病毒基因定位和农艺性状及品质性状的QTL分析中的应用 [D].南京:南京农业大学,2001.
Wang Y J.Establishment and adjustment of RIL population and its application to map construction,mapping genes resistant to SMV and QTL analysis of agronomic & quality traits in soybeans [D].Nanjing:Nanjing Agricultural University,2001.
[15] 郭志刚.数量性状主基因+多基因混合遗传分析的应用研究 [D].南京:南京农业大学,1998.
Guo Z G.Studies on apply using major gene and polygene mixed inheritance model of quantitative traits [D].Nanjing:Nanjing Agricultural University,1998.
[16] 刘顺湖,周瑞宝,盖钧镒.大豆蛋白质有关性状遗传的分离分析 [J].作物学报,2009,35(11):1958-1966.
Liu S H,Zhou R B,Gai J Y.Segregation analysis for inheritance of protein related traits in soybean [Glycinemax(L) merr] [J].Acta Agronomica Sinica,2009,35(11):1958-1966.
[17] 夏友霖,敬昱霖,毛金雄.花生晚斑病抗性遗传分离分析 [J].中国油料作物学报,2015,37(2):134-140.
Xia Y L,Jing Y L,Mao J X.Segregation analysis of resistance to late leaf spot in peanut [J].Chinese Journal of Oil Crop Science,2015,37(2):134-140.
[18] 李兰周,刘风珍,万勇善,等.花生荚果和籽仁相关性状的主基因+多基因混合遗传模型分析 [J].华北农学报,2013,28(5):116-123.
Li L Z,Liu F Z,Wan Y S,et al.Genetic analysis of pod and kernel characters by major gene plus polygene mixed inheritance model in peanut [J].Acta Agriculturae Boreali-Sinica,2013,28(5):116-123.
[19] 黄冰艳,张新友,苗利娟,等.花生油酸和亚油酸含量的遗传模式分析 [J].中国农业科学,2012,45(4):617-624.
Huang B Y,Zhang X Y,Miao L J,et al.Inheritance analysis of oleic acid and linoleic acid content ofArachishypogaeaL. [J].Scientia Agricultura Sinica,2012,45(4):617-624.
[20] 刘 华,张新友,崔党群,等.花生蛋白质和脂肪含量的主基因+多基因遗传分析[J].江苏农业科学,2011(2):127-130.
Liu H,Zhang X Y,Cui D Q,et al.Genetic analysis of protein and fat using major gene plus polygene methods in peanut(ArachishypogaeaL.) [J].Jiangsu Agricultural Science,2011(2):127-130.
[21] 张洁夫,戚存扣,浦惠明,等.甘蓝型油菜含油量的遗传与QTL 定位 [J].作物学报,2007,33(9):1495-1501.
Zhang J F,Qi C K,Pu H M,et al.Inheritance and QTL identification of oil content in rapeseed (BrassicanapusL.) [J].Acta Agronomica Sinica,2007,33(9):1495-1501.
[22] 李洪戈,余坤江,郭婷婷,等.甘蓝型油菜无花瓣性状的主基因+多基因遗传分析 [J].江苏农业学报,2014,30(2):253-258.
Li H G,Yu K J,Guo T T,et al.Genetic analysis of apetalous trait inBrassicanapususing mixed model of major gene and polygene [J].Jiangsu Journal of Agricultural Science,2014,30(2):253-258.
[23] 丛 野,程 勇,邹崇顺,等.甘蓝型油菜发芽种子耐湿性的主基因+多基因遗传分析 [J].作物学报,2009,35(8):1462-1467.
Cong Y,Cheng Y,Zou C S,et al.Genetic analysis of waterlogging tolerance for germinated seeds of rapeseed (BrassicanapusL.)with mixed model of major gene plus polygene [J].Acta Agronomica Sinica,2009,35(8):1462-1467.
[24] 董 娜,李成奇,王清连,等.不同生态环境下短季棉早熟及相关性状的混合遗传 [J].棉花学报,2010,22(4):304-311.
Dong N,Li C Q,Wang Q L,et al.Mixed inheritance of earliness and its related traits of short-season cotton under different ecological environments [J].Cotton Science,2010,22(4):304-311.
[25] 殷剑美,武耀廷,朱协飞,等.陆地棉产量与品质性状的主基因与多基因遗传分析 [J].棉花学报,2003,15(2):67-72.
Yin J M,Wu Y T,Zhu X F,et al.Genetic analysis of yield traits and fiber qualities by using major gene plus polygene mixed inheritance model in upland cotton(G.hirsutumL.) [J].Cotton Science,2003,15(2):67-72.
[26] 李斯深,陈茂学,王洪刚.利用重组自交系(RILs) 群体进行质量-数量性状的遗传分析:遗传模型和小麦产量性状遗传 [J].作物学报,2001,27(6):896-904.
Li S S,Chen M X,Wang H G.Genetic analysis on qualitative-quantitative traits by using populations of recombinant inbred lines (RILs):genetic models and inheritance of yield traits in wheat [J].Acta Agronomica Sinica,2001,27(6):896-904.
[27] 林婷婷,王 立,张 琳,等.不结球白菜叶绿素含量的主基因+多基因混合遗传分析 [J].南京农业大学学报,2014,37(5):34-40.
Lin T T,Wang L,Zhang L,et al.Genetic analysis of chlorophyll content using mixed major gene plus polygene inheritance model in non-heading Chinese cabbage [J].Journal of Nanjing Agricultural University,2014,37(5):34-40.
[28] 高美玲,袁成志,王玉书.甜瓜叶面积RIL群体主基因+多基因混合遗传模型分析 [J].东北农业大学学报,2014,45(1): 46-51.
Gao M L,Yuan C Z,Wang Y S.Genetic analysis of leaf area in melon (CucumismeloL.) with major gene plus polygene mixed genetic model by using recombinant inbred lines (RILs) population [J].Journal of Northeast Agricultural University,2014,45(1):46-51.
[29] 缪体云,刘玉梅,方智远,等.一个结球甘蓝DH群体主要农艺性状的遗传效应分析 [J].园艺学报,2008,35(1):59-64.
Miao T Y,Liu Y M,Fang Z Y,et al.Genetic analysis of the main agronomic traits of DH population inBrassicaoleraceavar.capitata[J].Acta Horticulturae Sinica,2008,35(1):59-64.
[30] 夏 征,陈甫堂,赵林英,等.亚麻主要性状遗传力和遗传相关的初步研究 [J].中国麻作,1985(2):37-41.
Xia Z,Chen F T,Zhao L Y,et al.Studies on the main trait heritability and genetic releated in flax [J].China’s Fiber Crops,1985(2):37-41.
[31] 王玉富,颜忠峰,乔广君,等.亚麻种子的含油率及其遗传与相关的研究 [J].黑龙江农业科学,1993(2):14-17.
Wang Y F,Yan Z F,Qiao G J,et al.Studies on the main trait heritability and genetic releated in flax [J].Heilongjiang Agricultural Sciences,1993(2):14-17.
[32] 曹锡文,刘 兵,章元明.植物数量性状分离分析Windows软件包SEA的研制 [J].南京农业大学学报,2013,36(6):1-6.
Cao X W,Liu B,Zhang Y M.SEA:a software package of segregation analysis of quantitative traits in plants [J].Journal of Nanjing Agricultural University,2013,36(6):1-6.
Major gene plus polygene inheritance analysis of crude fat content in oil flax (LinumusitatissimumL.)
HUA Qingchuna,ZHAO Lib,WANG Liminb,ZHAO Weib,DANG Zhaob,ZHANG Jianpingb,DANG Zhanhaib
(aWheatResearchInstitute,bCropResearchInstitute,GansuAcademyofAgriculturalSciences,Lanzhou,Gansu730070,China)
【Objective】 This study aimed to reveal the genetic mechanism of oil flax crude fat content.【Method】 A total of 233 recombinant inbred lines (RILs) derived from DYM×STS (F6∶7) along with their P1and P2were planted in three environments including Dingxi Gansu,Guyuan Ningxia,and Zhangjiakou Hebei.Crude fat contents were analyzed with Soxhlet Auto analyzer and mixed major gene plus polygene inheritance model was used to analyze P1,P2and RILs.【Result】 The crude fat contents appeared wide variation and transgressive inheritance,and the frequency distribution fitted normal distribution.The genetic model of crude fat content was 4MG-AI in three different environments.The crude fat content was controlled by 4 major genes with additive epistatic effect but without polygenic effect.The main gene heritability values of E1,E2,and E3 were 92.80%,99.02%,and 80.71%,while their environmental effects were 7.20%,0.98% and 19.29%,respectively.【Conclusion】 The oil flax crude fat content was controlled by major gene with additive epistatic effect.Attention should be paid to the use of main genes and consideration of interaction among genes for breeding oil flax with high oil content.
oil flax;crude fat;major gene plus polygene mixed inheritance
时间:2016-10-09 10:08
10.13207/j.cnki.jnwafu.2016.11.012
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20161009.1008.024.html
2016-04-13
国家自然科学基金项目“油用亚麻遗传图谱构建及亚麻酸含量的QTL定位研究”(31160291),“胡麻抗盐相关性状选择及QTL定位研究”(31560347)
化青春(1968-),女,甘肃会宁人,经济师,主要从事农作物新品种选育研究。E-mail:huaqch@163.com
赵 利(1973-),女,陕西武功人,研究员,博士,硕士生导师,主要从事亚麻遗传育种、种质资源及生理生化研究。 E-mail:zhyj0801@126.com
S565.903
A
1671-9387(2016)11-0083-07
猜你喜欢
黑龙江农业科学(2024年3期)2024-04-12 00:00:00
阜阳师范大学学报(自然科学版)(2022年1期)2022-04-02 12:35:16
食品工业科技(2020年24期)2020-12-09 07:36:18
农家科技中旬版(2020年7期)2020-08-10 08:52:47
系统管理学报(2018年3期)2018-08-13 01:05:28
系统管理学报(2018年2期)2018-08-13 01:04:38
新课程·下旬(2017年10期)2018-01-02 08:05:46
化工管理(2017年20期)2017-08-16 03:11:02
石家庄铁路职业技术学院学报(2017年4期)2017-05-25 13:26:34
中国工程咨询(2017年8期)2017-01-31 03:00:32
