
休眠期前富士苹果树碳代谢探究
2016-12-22 05:58温志静魏婵婵楚克欢
西北农林科技大学学报(自然科学版) 2016年11期
李 燕,温志静,魏婵婵,楚克欢,梁 俊
(西北农林科技大学 园艺学院,陕西 杨凌 712100)
休眠期前富士苹果树碳代谢探究
李 燕,温志静,魏婵婵,楚克欢,梁 俊
(西北农林科技大学 园艺学院,陕西 杨凌 712100)
【目的】 研究苹果树进入休眠期前的碳代谢规律,探讨碳代谢与进入休眠的关系。【方法】 以富士苹果树1年生枝为试材,分析其韧皮部、木质部和芽中可溶性总糖、还原糖、蔗糖和淀粉的含量及其相关酶(酸性转化酶(AI)、中性转化酶(NI)、蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)和淀粉酶)活性的变化。【结果】 随着休眠期的推进,蔗糖、还原糖和可溶性总糖含量在果树不同部位的分布为韧皮部>芽>木质部,且任意两者糖组分之间均极显著正相关;进入休眠期前,果树木质部中淀粉含量不断积累,其他碳水化合物含量变化不大,在休眠期之后,木质部中淀粉与可溶性总糖含量显著性负相关(相关系数为-0.856*)。树体进入休眠期前后,芽中各酶活性均高于韧皮部;韧皮部和芽中AI活性与蔗糖含量极显著性负相关(相关系数分别为-0.909**,-0.783**);芽中SPS活性与蔗糖含量显著正相关(相关系数为0.649*)。【结论】 苹果在进入休眠期前,木质部开始储藏淀粉,随着休眠期的来临,木质部中积累的大量淀粉被分解使可溶性糖含量升高,与此同时,各部位的蔗糖含量快速上升,这与SPS活性的升高、AI活性的降低有关。
富士苹果树;休眠期前;枝条;芽;碳代谢
苹果树从落叶到萌芽的这一段时间称为休眠期[1]。休眠期是果树重要的物候期,体内储藏的碳水化合物是其抵御冬季低温胁迫、维持最基本的生理代谢、花芽进一步分化和春季萌芽生长的重要物质基础[2]。进入休眠期前各种碳水化合物的积累和相关酶活性的变化,对进入休眠期的果树及后期解除休眠后树体营养的整体状况都起着举足轻重的作用。目前,相关研究主要集中于果树休眠期间[3-7]或休眠解除时期[8]枝条、芽中某几种碳水化合物的变化,以及此期与碳水化合物相关酶活性的变化[2,9]等方面,但关于果树进入休眠期前碳水化合物的积累及相关酶活性变化的研究很少,而对于苹果树的研究尚未见报道。本试验以富士苹果树的枝条和芽为材料,测定了其进入休眠期(即落叶前后)前韧皮部、木质部和芽中的碳水化合物及与之相关的代谢酶活性的变化,研究了苹果枝条和芽在此期间的糖代谢规律,探讨了果树休眠期前碳代谢与进入休眠的关系,以期为苹果树体内有机碳代谢研究、有机碳肥开发奠定基础。
1 材料与方法
1.1 材 料
于2014年9月至12月,以西北农林科技大学白水苹果试验站内生长健壮、树势一致的8年生矮化“富士”苹果树为试材,采用随机区组设计,每7株为1个小区,3次重复。在此期间每隔10 d采样1次,每次每株采集长势一致的1年生基生结果枝1枝,共21枝。在试验地立即将叶片清除干净并带回实验室,用水冲洗干净之后迅速将木质部、韧皮部和芽分离。称取一部分韧皮部和芽进行研磨,用于测定糖代谢相关酶活性;剩余的在105 ℃杀青15 min,60 ℃烘干至恒质量,磨碎后过0.175 mm(80目)筛,用于测定各物质含量。
1.2 碳水化合物含量测定
1.2.1 可溶性糖和淀粉的提取 根据高俊凤[10]的方法提取可溶性糖和淀粉。称取枝条、芽干样各 0.5 g和0.25 g于离心管,加入8 mL 体积分数80%的乙醇,80 ℃水浴30 min,3 500 r/min离心15 min,上清液转入25 mL容量瓶中,再向沉淀中加入6 mL体积分数80%乙醇,如上述方法浸提2次,将上清液合并至25 mL容量瓶中,用乙醇定容至25 mL。取8 mL在沸水浴中蒸干,加入20 mL蒸馏水充分溶解后待测。提取后剩余的残渣利用高氯酸法来提取淀粉。即往残渣中加蒸馏水2 mL,沸水浴糊化15 min,冷却后加入9.2 mol/L高氯酸,搅拌15 min,加蒸馏水4 mL,混匀,4 000 r/min离心10 min,上清液转入50 mL容量瓶中;再向沉淀中加入2 mL 4.6 mol/L 高氯酸,搅拌提取15 min,加入5 mL蒸馏水,混匀后离心10 min,合并上清液,用蒸馏水洗涤沉淀2次,每次5~6 mL,合并上清液并用蒸馏水定容至刻度后待测。
1.2.2 可溶性总糖、还原糖含量的测定 可溶性总糖含量用蒽酮比色法[10]在620 nm下测定,根据葡萄糖的标准曲线计算可溶性总糖含量;还原糖含量用3,5-二硝基水杨酸[10]测定520 nm下的光密度值,根据葡萄糖的标准曲线测定还原糖含量。
1.2.3 淀粉含量的测定 淀粉含量用蒽酮比色法[10]测定620 nm下的光密度值,根据葡萄糖的标准曲线计算样品中淀粉含量。
1.2.4 蔗糖含量的测定 蔗糖含量采用间苯二酚比色法[11-12]测定。
1.3 酶液提取及测定
酶液制备基本参照Keller等[13]的方法进行。分别取新鲜的芽、韧皮部各0.5和1.0 g,分3次加7 mL酶提取液,冰浴研磨成匀浆。提取液成分为50 mmol/L HEPES-NaOH (pH 7.5),10 mmol/L MgCl2,1 mmol/L EDTA,2.5 mmol/L DTT,体积分数0.05% Triton-100 和1 g/L BSA。然后将匀浆倒入10 mL离心管中,在2 ℃下11 000 r/min离心20 min,上清液在2 ℃下用透析液(去除Triton-100稀释10倍的提取液)透析24 h,透析后的酶液用于酶活性分析。
蔗糖合成酶(SS)活性测定按照Rufty 等[14]的方法,通过测定从UDPG(尿苷二磷酸葡萄糖)生成的蔗糖来定量;蔗糖磷酸合成酶(SPS)活性测定按照Zhu等[15]的方法,通过测定从UDPG生成的蔗糖来定量;酸性转化酶(AI)、中性转化酶(NI)和淀粉酶活性测定按照Merlo等[16]的方法,测定释放还原糖的量来表示酶活性。
1.4 数据分析
试验数据采用Excel 2003、Origin 7.5、SPSS 19.0软件进行处理与相关数理统计分析。
2 结果与分析
2.1 休眠期前后富士苹果树枝条和芽中碳水化合物含量的变化
2.1.1 还原糖和可溶性总糖含量 如图1所示,随着休眠期的推进,还原糖与可溶性总糖含量在富士苹果树1年生枝条不同部位的分布为韧皮部>芽>木质部,韧皮部和芽中还原糖、可溶性总糖含量较高,而木质部中两者含量均较低。尽管可溶性总糖含量略高于还原糖含量,但大多数情况下两者不存在显著性差异。苹果树在11月8日左右(此时期开始落叶)开始进入休眠期,由图1可知,休眠期前还原糖和可溶性总糖含量在各部位变化不大,进入休眠期后各部位可溶性总糖和还原糖含量都呈现快速增加的趋势。

图 1 休眠期前后富士苹果树各部位还原糖和可溶性总糖含量的变化
2.1.2 蔗糖含量 由图2可以看出,在富士苹果树1年生枝条韧皮部中蔗糖含量最高,其次是芽、木质部,这与可溶性总糖、还原糖的分布规律一致。休眠期前,木质部中蔗糖含量极低,但在进入休眠期后,其含量迅速上升;韧皮部和芽中蔗糖含量在休眠期前比较稳定,之后也呈快速上升的现象。
2.1.3 淀粉含量 由图3可以看出,淀粉含量在富士苹果树1年生枝条各部位的分布基本呈现为木质部>韧皮部>芽,木质部和韧皮部中淀粉含量变化比较剧烈,大致呈现先升后降的趋势,但木质部中其含量是在进入休眠期1周后达到最高值,而韧皮部是在休眠期前即达最高值;芽中淀粉含量较低且变化较小,尤其进入休眠期后,淀粉含量比较稳定。
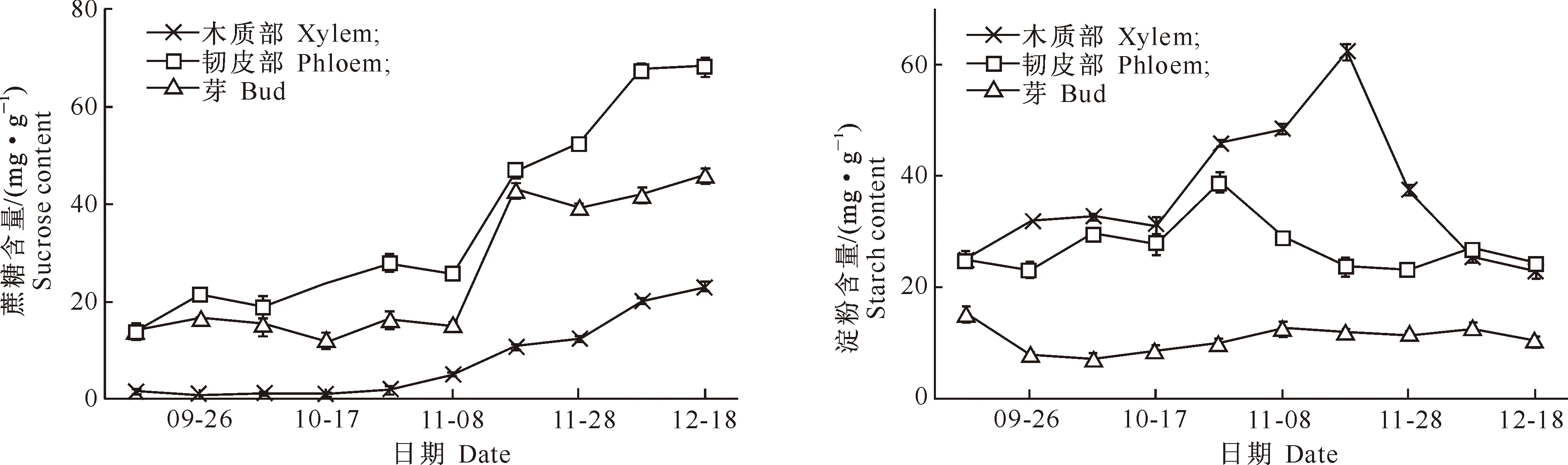
图 2 休眠期前后富士苹果树各部位蔗糖含量的变化

图 3 休眠期前后富士苹果树各部位淀粉含量的变化
2.2 休眠期前后富士苹果树不同部位糖组分之间的相关性
由表1可知,在休眠期前,富士苹果树1年生枝条芽、木质部和韧皮部中的可溶性总糖、蔗糖和还原糖任意两者之间都呈极显著正相关关系。在休眠期以后,对木质部中淀粉与可溶性总糖含量进行了相关分析,两者之间呈显著性负相关(相关系数为-0.856*),但韧皮部、芽中两者无显著相关关系。

表 1 休眠期前富士苹果树不同部位碳水化合物含量之间的相关性分析
注:相关显著水平:*.P<0.05,**.P<0.01。表2同。
Note:Significant correlation level:*.P<0.05,**.P<0.01.The same for table 2.
2.3 休眠期前后富士苹果树枝条和芽中糖代谢相关酶活性的变化
2.3.1 转化酶(AI和NI)活性 如图4所示,富士苹果树1年生枝条芽中AI和NI活性均明显高于韧皮部,且同一器官中两者活性变化趋势相似,AI在此期间活性较高,整体呈现下降的趋势,直到11月28日,其活性降到比较平稳的范围,并保持较低水平;NI在韧皮部和芽中的活性很低,前期活性比较稳定,在休眠期前突然上升并达到最大值,进入休眠期后其活性又迅速下降并降至最低。

图 4 休眠期前后富士苹果树韧皮部和芽中转化酶活性的变化
2.3.2 蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性 休眠期前后富士苹果树韧皮部和芽中SS和SPS活性的变化见图5。

图 5 休眠期前后富士苹果树韧皮部和芽中SS和SPS活性的变化
如图5所示,富士苹果树1年生枝条韧皮部和芽中SS活性的变化比较复杂,基本呈现下降-上升-下降的变化趋势,芽中变幅较大且活性明显高于韧皮部。SPS在韧皮部和芽中活性变化趋势一致,前期比较平稳,但在果树进入休眠期后迅速上升。
2.3.3 淀粉酶活性 休眠期前后富士苹果树韧皮部和芽中淀粉酶活性的变化见图6。

图 6 休眠期前后富士苹果树韧皮部和芽中淀粉酶活性的变化
由图6可知,富士苹果树1年生枝条中淀粉酶活性与上述4种酶类似,在芽中的活性高于韧皮部,淀粉酶活性在韧皮部和芽中大致都呈先升后降的变化趋势,即在休眠期前淀粉酶活性逐渐升高,之后快速下降,而且芽中活性开始下降的时间晚于韧皮部1周左右。
2.4 休眠期前后苹果树不同部位碳水化合物含量与相关代谢酶活性的关系
由表2可以看出,富士苹果树1年生枝条韧皮部中AI与蔗糖、还原糖、可溶性总糖含量呈显著或极显著负相关(相关系数分别为-0.909**,-0.737*,-0.888**),而芽中AI仅与蔗糖含量呈极显著负相关(相关系数为-0.783**)。韧皮部中NI与还原糖和可溶性总糖含量呈极显著负相关(相关系数分别为-0.913**和-0.796**),芽中NI与蔗糖、还原糖、可溶性总糖含量呈显著或极显著负相关(相关系数分别为-0.805**,-0.744**,-0.716*)。韧皮部中SPS与还原糖含量呈显著正相关(相关系数为0.687*),而芽中SPS与蔗糖含量呈显著正相关(相关系数为0.649*)。

表 2 休眠期前后富士苹果树韧皮部和芽中碳水化合物含量与相关酶活性的关系
3 讨论与结论
3.1 休眠期前后苹果树各部位碳水化合物含量变化与休眠的关系
果树在进入休眠期前,碳水化合物作为体内的主要能量来源在此期间会进行一系列的代谢和分配,以保证其生命活动的正常进行。本研究结果显示,果树进入休眠期前,1年生枝条木质部中淀粉含量不断积累,其他碳水化合物含量变化不大,这与前人的研究结果[17-18]相似。进入休眠期之后,木质部中淀粉与可溶性总糖含量显著负相关(相关系数为-0.856*),而韧皮部和芽中的淀粉含量与可溶性总糖含量没有显著相关性,这表明韧皮部、芽中可溶性总糖含量的升高很可能是由于木质部中淀粉的水解,即果树进入休眠期前,木质部是储藏淀粉的主要部位,此后随着外界环境温度的降低,为树体进入休眠期做准备,淀粉迅速被水解为可溶性糖以适应树体自身内部的需要。李秀珍等[2]在对欧李的研究中就提出,休眠期韧皮部和芽中淀粉含量与可溶性总糖含量之间没有相关性,可溶性糖的来源需要进一步探讨,本研究结果恰好解释了这一现象。本研究还发现,在进入休眠期后果树各部位中可溶性总糖含量迅速升高,原因是可溶性总糖可提高低温下果树的渗透调节作用,进而增强植株抵抗低温的能力,这与前人对果树抗性研究的结果[19-21]一致。蔗糖含量在进入休眠期前很低,在之后明显升高,这与王慧等[8]对油桃的研究结果相似。
3.2 休眠期前后苹果树韧皮部和芽中碳水化合物含量变化与相关代谢酶活性的关系
本研究结果显示,富士苹果树1年生枝条韧皮部中各酶活性均低于芽中,相关性分析表明,韧皮部和芽中AI活性与蔗糖含量极显著性负相关(相关系数分别为-0.909**和-0.783**),而韧皮部NI活性与蔗糖含量无显著相关性;进入休眠期后SPS活性突然增大,这与蔗糖含量在此时快速增加相一致。通常认为SPS是促进蔗糖合成的一种重要酶[22]。相关性分析认为,芽中SPS活性与蔗糖含量显著正相关(相关系数为0.649*),可以认为芽中SPS是合成蔗糖的关键酶。SS是蔗糖双向作用酶类,其变化复杂,与蔗糖含量的变化无相关性,但是SS活性的变化与蔗糖含量变化有时相反有时一致,这恰恰体现了该酶的特性。由以上分析可知,果树休眠期前蔗糖调控的关键酶是AI和SPS,NI和SS起调节作用。
综上所述,苹果树在进入休眠期前,木质部开始储藏淀粉,随着休眠期的到来,木质部中积累的大量淀粉开始分解,使各部位可溶性糖含量升高,以此来适应逐渐变冷的外界环境,与此同时,各部位的蔗糖含量快速上升,这与SPS活性的升高、AI活性的降低有密切关系。
[1] 杨鲜明.苹果树休眠期的管理 [J].甘肃农业科技,1996(6):27.
Yang X M.The management of dormant apple trees [J].Gansu Agricultural Science and Technology,1996(6):27.
[2] 李秀珍,陈苏丹,李天忠.休眠期欧李碳水化合物代谢与休眠关系的分析 [J].中国农业大学学报,2012,17(4):75-80.
Li X Z,Chen S D,Li T Z.Relation of bud dormancy to carbohydrate metabolism inCerasushumilis(Beg.) Sok [J].Journal of China Agricultural University,2012,17(4):75-80.
[3] 高东升,夏 宁,王兴安.休眠桃树枝条中碳水化合物的含量变化和外源生长调节剂对破除休眠的效应 [J].植物生理学通讯,1999,35(1):10-12.
Gao D S,Xia N,Wang X A.Changes of carbohydrate content and effect of exogenous growth regulators on dormancy-breaking during dormant period of peach shoots [J].Plant Physiology Communications,1999,35(1):10-12.
[4] 申海林,海 江,徐继忠.休眠期杏花芽若干生理指标变化的研究 [J].河北农业大学学报,2004,27(6):41-43.
Shen H L,Hai J,Xu J Z.Study on some physiological indexes during dormant period of apricot bud [J].Journal of Agricultural University of Hebei,2004,27(6):41-43.
[5] 张 双.梨休眠期芽内营养变化及呼吸代谢研究 [D].河北保定:河北农业大学,2009.
Zhang S.Study on nourishment changes and respiratory metabolism of dormancy pear buds [D].Hebei,Baoding:Hebei Agricultural University,2009.
[6] 田莉莉,方金豹,王力荣,等.华光油桃解除休眠过程中几项生理指标的变化 [J].果树学报,2006,23(1):121-124.
Tian L L,Fang J B,Wang L R,et al.Changes of several physiological indexes during dormancy releasing in nectarine cultivar Huaguang [J].Journal of Fruit Science,2006,23(1):121-124.
[7] 郑鹏华,刘国琴,Hussain S,等.‘翠冠’梨花芽休眠期碳水化合物变化及其相关基因表达研究 [J].园艺学报,2013,40(2):325-332.
Zheng P H,Liu G Q,Hussain S,et al.Studies on changes in carbohydrate contents and related gene expression in floral buds of ‘Cuiguan’ pear during dormancy [J].Acta Horticulturae Sinica,2013,40(2):325-332.
[8] 王 慧,李 玲,谭 钺,等.休眠期间油桃花芽碳水化合物代谢及其相关基因的表达变化 [J].植物生理学报,2011,47(6):595-600.
Wang H,Li L,Tan Y,et al.Changes in carbohydrate metabolism related gene expression in nectarine floral buds during dormancy [J].Plant Physiology Journal,2011,47(6):595-600.
[9] 唐洪梅.温度处理对欧李一年生休眠枝中淀粉酶及相关产物的影响 [D].河南洛阳:河南科技大学,2012.
Tang H M.Effects of temperature on the activity of amylase its related products in annual dormant branches ofCerasushumilis(Beg.) Sok [D].Luoyang,Henan:Henan University of Science and Technology,2012.
[10] 高俊凤.植物生理学实验指导 [M].北京:高等教育出版社, 2005:144-148.
Gao J F.Experimental guidance of plant physiology [M].Beijing:Higher Education Press,2005:144-148.
[11] 韩振海,陈昆松.实验园艺学 [M].北京:高等教育出版社,2006:389-392.
Han Z H,Chen K S.Experimental horticulture [M].Beijing: Higher Education Press,2006:389-392.
[12] 肖世远.间苯二酚光度法测定蔗糖的适宜条件 [J].四川师范学院学报(自然科学版),1998,19(3):57-59.
Xiao S Y.The suitable conditions of measuring cane sugar by using resorcinol-photometric method [J].Journal of Sichuan Teachers College (Natural Science),1998,19(3):57-59.
[13] Keller F,Ludlow M M.Carbohydrate metabolism in drought-stressed leaves of pigeonpea (Cajanuscajan) [J].Journal of Experimental Botany,1993,44(265):1351-1359.
[14] Rufty T W J,Huber S C.Changes in starch formation and activities of sucrose phosphate synthase and cytoplasmic fructose-1,6-bisphosphatase in response to source-sink alterations [J].Plant Physiol,1983,72:474-480.
[15] Zhu Y J,Komor E,Moore P H.Sucrose accumulation in the sugarcane stem is regulated by the difference between the activities of soluble acid invertase and sucrose phosphate synthase [J].Plant Physiol,1997,115:609-616.
[16] Merlo L,Passera C.Changes in carbohydrate and enzyme levels during development of leaves ofPrunuspersica,a sorbitol synthesizing species [J].Physiologia Plantarum,1991,83:621-626.
[17] Yoshioka H,方金豹.苹果树体内碳水化合物代谢的年变化 [J].国外农学(果树),1989(2):5-8.
Yoshioka H,Fang J B.The annual changes of carbohydrate metabolism in apple trees [J].Foreign Agriculture (Fruit Tree),1989(2):5-8.
[18] 夏国海,罗新书.苹果幼树秋季14C同化物贮藏及再利用 [J].园艺学报,1990,17(2):96-102.
Xia G H,Luo X S.Studies on the characters of storage metabolism and reuse of14C-assimilates synthesized in autumn in young apple trees [J].Acta Horticulturae Sinica,1990,17(2):96-102.
[19] 刘 艳,赵虎成,李 雄,等.梨枝条中淀粉、还原糖及脂类物质动态变化与抗寒性的关系 [J].内蒙古农业大学学报(自然科学版),2002,23(1):57-60.
Liu Y,Zhao H C,Li X,et al.Dynamics of starch grain sugar materials and lipids on pear shoots and its relationship to cold resistance [J].Journal of Inner Mongolia Agricultural University (Natural Science Edition),2002,23(1):57-60.
[20] 沙广利,郭长城,睢 薇,等.梨抗寒性遗传的研究 [J].果树科学,1996,13(3):167-170.
Sha G L,Guo C C,Sui W,et al.Studies on inheritance of cold-hardiness of pear [J].Journal of Fruit Science,1996,13(3):167-170.
[21] Tabaei-Aghdaei S R,Pearce R S,Harrison P.Sugars regulate cold-induced gene expression and freezing-tolerance in barley cell cultures [J].Journal of Experimental Botany,2003,54:1565-1575.
[22] Chapman G W,Horvat R J.Changes in nonvolatile acids,sugars,pectin and sugar composition of pectin during peach (Cv.Monroe) maturation [J].J Agric Food Chem,1990,38(2):383-387.
Carbon metabolism of Fuji before dormancy
LI Yan,WEN Zhijing,WEI Chanchan,CHU Kehuan,LIANG Jun
(CollegeofHorticulture,NorthwestA&FUniversity,Yangling,Shaanxi712100,China)
【Objective】 This study investigated the carbon metabolism of apple tree before dormancy period.【Method】 Annual shoots of Fuji were selected to analyze soluble sugar,reducing sugar,sucrose,starch contents and changes in activities of related enzymes (AI,NI,SS,SPS and amylase) in xylem,phloem and bud.【Result】 Along with the advancement of dormancy,the distribution of sucrose,reducing sugar and total soluble sugar content in different parts of trees was in the order of phloem>bud>xylem,and the correlation between any two was significantly positive.Before dormancy,starch content in xylem continued to accumulate,while other carbohydrate contents barely changed.At early dormancy starch and soluble sugar contents in the xylem had significant negative correlation with coefficient of -0.856*.During the experiment,enzyme activities in bud were higher than in phloem.AI and sucrose content had significant negative correlation in phloem and bud with coefficients of -0.909**and -0.783**,respectively.SPS and sucrose content in bud was significantly positively related with coefficient of 0.649*.【Conclusion】 Before dormancy,the xylem began to store starch,which was hydrolyzed during dormancy to form soluble sugar.At the same time,sucrose content in each organ increased rapidly relating to increase of SPS activity and decrease of AI activity.
Fuji apple;before dormancy;shoot;bud;carbon metabolism
时间:2016-10-09 10:08
10.13207/j.cnki.jnwafu.2016.11.018
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20161009.1008.036.html
2015-05-14
“十二五”国家科技支撑计划项目(2014BAD10B02);陕西省科技统筹创新工程计划项目(2011KTZB02-02-04)
李 燕(1990-),女,山东新泰人,在读硕士,主要从事苹果品质改良研究。E-mail:liyan901112@163.com
梁 俊(1963-),男,陕西高陵人,教授,博士,硕士生导师,主要从事果树生理与生态研究。 E-mail:strongca@163.com
S661.1
A
1671-9387(2016)11-0127-06
猜你喜欢
植物研究(2021年2期)2021-02-26
蔬菜(2018年12期)2018-01-16
科技与创新(2016年22期)2017-03-30
科技视界(2016年20期)2016-09-29
特产研究(2016年3期)2016-04-12
天津农业科学(2015年11期)2015-12-03
安徽农学通报(2014年7期)2014-04-29
中国果业信息(2014年11期)2014-01-23
