
Prenatal progesterone exposure of male rats induces morphometric and histological changes in testes
2016-11-30 12:33SamyIsmailAhmedAdilSalimElsheikhTahirOsmanAliGamalAliAttia
Samy Ismail Ahmed, Adil Salim Elsheikh, Tahir Osman Ali, Gamal Ali Attia
1Department of Anatomy Faculty of Medicine Najran University, KSA
2Department of Applied Medical sciences, Community College Najran, KSA
3Dean of the Faculty of Graduate Studies & Scientific Research, National Ribat University, Sudan
Prenatal progesterone exposure of male rats induces morphometric and histological changes in testes
Samy Ismail Ahmed1, Adil Salim Elsheikh2*, Tahir Osman Ali3, Gamal Ali Attia1
1Department of Anatomy Faculty of Medicine Najran University, KSA
2Department of Applied Medical sciences, Community College Najran, KSA
3Dean of the Faculty of Graduate Studies & Scientific Research, National Ribat University, Sudan
ARTICLE INFO
Article history:
Received
Received in revised form
Accepted
Available online
Prenatal
Objective: To investigate the effects of prenatal progesterone exposure of male rats on testicular morphometry and histology. Methods: Twenty male Wizard rats born to dams treated with hydroxyprogesterone hexanoate (HPH) during the fi rst 2 weeks of gestation were employed to evaluate the ef fects of HPH on testicular morphometry, histology and cells quantities. Twenty male rats born to untreated dams were used as control. The rats were kept until they reached 90 days old, sacrif iced, testes dissected, weighed and their dimensions recorded. Histological sections were prepared and stained with H&E. The diameter of the seminiferous tubule (ST), height of germinal epithelium, and thickness of the interstitial space (IS) were measured microscopically. Furthermore testicular cells were quantif ied. Results: The results showed that experimental rats had signifi cantly (P < 0.001) diff erent testicular morphometry, histology and cells quantities compared to the control. The histological sections taken from testes of experiment group showed ST with unusual conf iguration, detached and/or folded basal lamina, thin germinal layer, wide IS with few LC, Sertoli cells experienced varying degrees of apoptosis and the lumen of ST contained cells debris and very rare few sperms. Conclusions: Prenatal HPH exposure of male rats adversely af fects their testicular morphometry and histological structure and presumably has crucial negative eff ects on their future fecundity.
1. Introduction
Recent data suggest deterioration of male reproductive health and this deterioration by no means is attributed to xenobiotic chemicals[1-3]. Xenobiotic hormones are widely used in human medicine. The xenobiotic hormones given to women during pregnancy are considered among the endocrine disrupting chemicals capable of modulating and disrupting the endocrine system of the male off -springs leading to deleterious eff ects on their reproductive health[4]. Prenatal exposure to steriod chemicals is known to induce testicular developmental disorders and consequently reduction in the fertility of adult males[5,6]. Irreversible testicular growth alteration, reduced sperm production and suppressed steroidogenesis have been reported after exposure to steroids[7, 8].
Synthetic progesterone (P4) is widely used in fertility clinics for many therapeutic purposes. It is traditionally used as contraceptive[9], ovulation inducer in primary infertility cases[10], and pregnancy support for threatened miscarriages during the fi rst trimester, abnormal uterine bleeding inhibitor[2] and as luteal supporter for transplanted IVF embryos[11]. This situation entails that many women around the world are exposed to higher dose of xenobiotic P4. These hormones are accused of eliciting adverse eff ects on reproductive health and fertility of future generations[9]. Prenatal exposure to P4reduced the body weight, testicular weight, total sperm count and damaged the ST and devoid them of sperm[12]. Histopathological study following in utero P4administration revealed severe testicular damage, decrease in seminiferous tubules (STs) diameters, reduction in number of spermatogenic cells, widening of tubular lumen and interstitial spaces with increment of necrozoospermia[12,13]. Xenobiotic agents interfere with synthesis, secretion, transport, metabolism, binding, elimination of natural blood hormones causing dis×order of homeostasis and consequently hinder reproduction and development[14].
Exposure of embryos to the xenobiotic HPC reduced the circulating testosterone levels, sperm cell count and increased necrozoospermia; indicating a probable inhibition of androgen synthesis[2,15]. In the light of the above mentioned studies; the current study investigated the eff ects of prenatal HPH exposure of male rats on testicular morphometry, histology and cells quantities.
2. Materials and methods
2.1. Animals’ management
Eight to ten weeks old females (n = 14) and males (n = 4) of albino Wistar rats were obtained from the animal house of the Faculty of Pharmacy, Jazan University, Kingdom of Saudi Arabia. They were transferred to the animal house of the Department of Anatomy, Faculty of Medicine, Najran University and grouped into two groups. Each group consisted of seven females and two males. They were kept at 12:12 h light/dark cycle, (28±7) °C temperature, fed on commercial pellet and off ered water ad libitum. The female rats were examined 3 times a day for the presence of cervical plugs and those proved mated were kept separated from males. The pregnant females were grouped into two groups and kept separate away from any stress in (90 cm× 45 cm× 15 cm) in sterilized polypropylene cages lined with woody husk. Group I served as control and groups II as the experimental group. The females in the experiment group were subcutaneously injected with 10 mg/kg BW of proluton depot (Hydroxyprogesterone hexanoate; Schering AG; Germany) on the 1st, 7thand 14thof gestation. The dam body weights were taken on each day of injection to adjust the dose. The prescribed dose of P4is within the range of the regular clinical dose given to pregnant women[12]. The females in the control group were injected with a placebo.
2.2. Tissue collection and preparation
Animals were anesthetized with chloroform and sacrifi ced with cervical dislocation. The testes were dissected and left testes and epididymides were weighed then fixed immediately in aqueous Bouin’s solution for 18 h, dehydrated into 70%, 90% and 100% ascending grades of alcohol, cleared with xylene and embedded in paraffi n wax. Sections were cut at 5 μm thickness by using an American Optical microtome (A0-821. USA). The sections were mounted onto glass slides, deparaffi nized and stained with H&E[16]. The testes were separated from their adjacent epididymdes and their diameters and lengths were measured. Relative testes and epididymides weights were then calculated per fi nal body weight[16].
2.3. Histometry
Morphometric measurers of testicles were done according to Thienpot et al.[17] and Batra et al.[18]. Longitudinal sections of each rat testis were stained with H&E. Then 10 round or nearly round ST were chosen randomly to measure their diameters, the height of germinal epithelium (GE), the thickness of interstitial space (IS) and the number of GE cells using Olympus BX-40 light microscope supported with an image Pro Plus program (100×). Two tubular diameters for each tubule were mapped and their mean recorded. The thickness of IS was measured by measuring three dimensions of spaces assumed to connect the centre of each space to the basement membrane of the surrounding ST. The mean of the three dimensions was calculated and multiplied by 2 to obtain the whole thickness of the IS. The GE height was obtained for the same tubules used to determine tubular diameter. GE epithelium was assumed from the basement membrane to the latest stage of GE (spermatids).
2.4. Cells quantitation
The testes of each rat were prepared for light microscopy. After animal scarification, testes were fixed by perfusion with Bouin’s fixative for 30 min. The testis was then cut into three vertical longitudinal slices, the middle slice including the mediastinum. After immersion and fi xation in Bouin’s for another 1.5 h, the slices were dehydrated in ethanol, cleared with xylene and embedded in paraffi n wax. From each testis slice fi ve sections of 5 μm were cut, thus 15 sections were obtained and mounted individually onto slides. LC of 15 IS of each rat testis were counted under Olympus BX-40 microscope supported with an Image Pro Plus program. The mean counts of LC were calculated for each group. Also mean counts of the SC and GE cells (spermatogonia type A & B and primary spermatocytes cells) were also recorded[19,20].
2.5. Experimental design
This experiment is a one factorial design to investigate the eff ects of prenatal exposure to HPH on pubertal rats’ testicular morphometry, histology and cells quantities. Forty puppies born to the experiment group (n = 20 puppies) and control group (n = 20 puppies) were allowed to grow for 90 d where they reached maturity. The testes were collected and prepared as above. Morphometry, histometry and cell quatitation were carried as described above.
2.6. Statistical analysis
Statistical analyses for all obtained parameters were performed by using SPSS-16.020 (Chicago, USA). Data were subjected to one way ANOVA and are expressed as mean±SD. The level of signifi cance was set at P < 0.05.
3. Results
3.1. Morphometric aspects
3.1.1. Testicular and epididymal weights
The experimental rats group showed a significant (P < 0.001) reduction in testicular and epididymal weights (Figure 1) compared to control group. The mean testicular weights of the control and experimental groups were (2.11±0.33) and (1.60±0.14) g and the mean epididymal weights were (0.67±0.10) and (0.43±0.06) g in the same respective. The relative testicular weight for the control and experimental rats were (0.01±0.01) and (0.01±0.01) g and the mean relative epididymal weight were (4.0±0.0) and (2.0±0.0) mg, respectively.
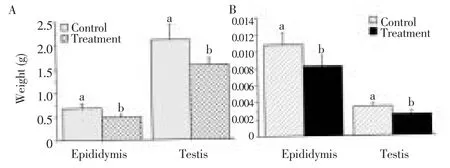
Figure 1. Mean testicular and epididymal weights (A) and relative weights (B).
3.1.2. Testicular dimensions
Figure 2 shows that the testicular dimensions of the control and the experiment rats are quite diff erent (P < 0.001). The mean length of the right testis of the control was (17.85±0.04) mm and that of the experiment rats was (15.57±0.04) mm while the width was (10.87±0.04) and (8.49±0.02) mm in the same respective.
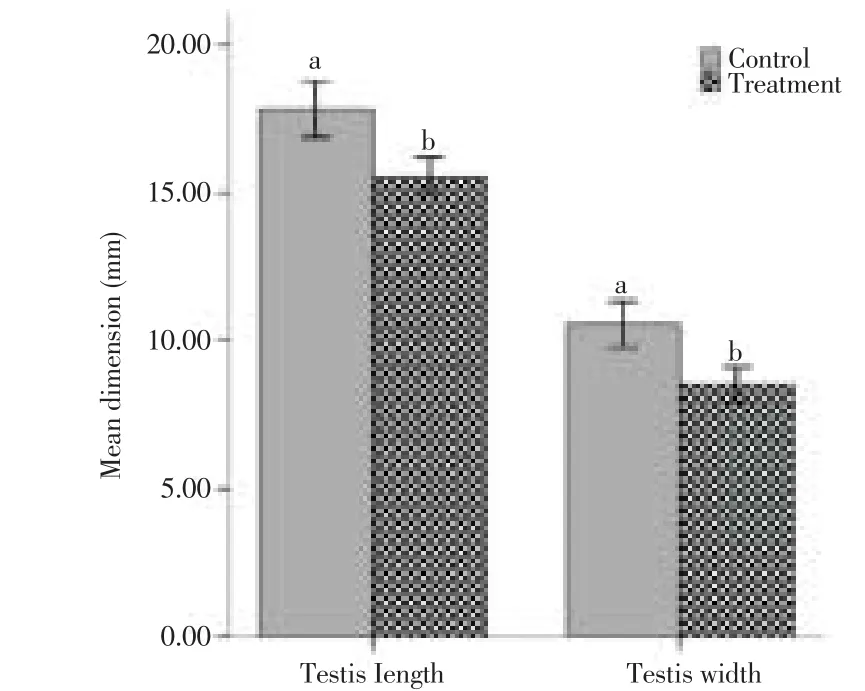
Figure 2. Compares dimensions of testes of control and experimental rats. Bars represent the mean±SD of 20 replicates.a,bP < 0.01.
3.1.3. Measurements of ST, GE and interstitial space
The diameter of the ST, GE height and the thickness of the IS of the control and experimental rats varied signifi cantly (P < 0.001). The mean diameters of ST of the control and experimental rats were (281.5±17.2) and (255.5±14.4) μm, respectively. The height of the GE was (93.7±9.6) and (72.6±7.7) μm, in the same respective as above. The thickness of the IS of the control was (110.5±11.3) μm and that of the experiment rats was (132.1±6.1) μm (Figure 3).
3.2. Histological aspects
Figure 4 shows the rats’ testicular parenchyma formed of ST and interstitial space. Figure 4A shows that the ST of control rats are separated from the interstitial space with an intact basement membrane. The cross sections of the ST appeared rounded or oval in shape containing spermatogenic and SC and resting on well-formed basement membrane formed of thin elongated simple epithelial cell layer and fi brous tissue. The diff erent types of the spermatogenic cells are easily identifi ed and so is the mitotic activity as revealed by the different stages of the spermatogenesis. The individual cells of the ST and interstitial tissue are clear and with normal features and the lumen contains obvious mass of sperms. Figure 4B illustrates many prominent abnormal features of the testis of experimental rats born to dams treated with HPC during pregnancy. The sections taken from testes of experiment group showed thin germinal layer compared to that of the control. The ST loses their usual confi guration with detached and/or folded basal lamina. The IS are wide with large blood vessels due to hypoplasia, shrinkage and destruction of ST. Figure 4C shows control rat ST at high magnifi cation (notice the normal germination and sperm masses). Figure 4D shows experimental rat ST with reduced layers of GE, sparse or no sperm masses and a wider ST lumen with obvious necrotic cellular debris.

Figure 4. Light photomicrographs of rats testes parenchyma.
As in Figure 5A, the IS (A) of experimental rat testes appeared wider, contained large blood vessels and semi-empty cells except of few LC, connective tissue cells and interstitial vacuoles. Figure 5B shows degenerative spermatogenic cells with apoptotic changes in the form of inter- and intra-cellular cavitations. The SC are few in most sections with great vesicular cytoplasm cavities resulting in abnormal spacing (vacuoles) between the germ cells of the seminiferous epithelium.
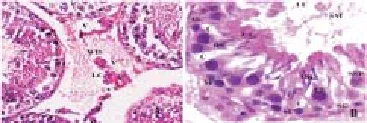
Figure 5. Micrograph A the interstitial space of experimental rat testis.
3.3. Testicular cells quantitation
Figure 6 shows that the counts of the diff erent testicular cells per cross section varied significantly (P < 0.01) with treatment. The mean counts of spermatogonia type A of the control rats and the experimental rats were (38.6±3.5 & 27.4±3.6) and type B were (36.5±3.1 & 21.8±4.5), respectively. The primary spermatocytes were (37.5±4.0 & 26.6±4.5), SC were (17.7±2.3 & 9.8±2.8) and LC per a single interstitial space were 5.8±1.5 & 2.1 ±1 .3 in the same order as above.
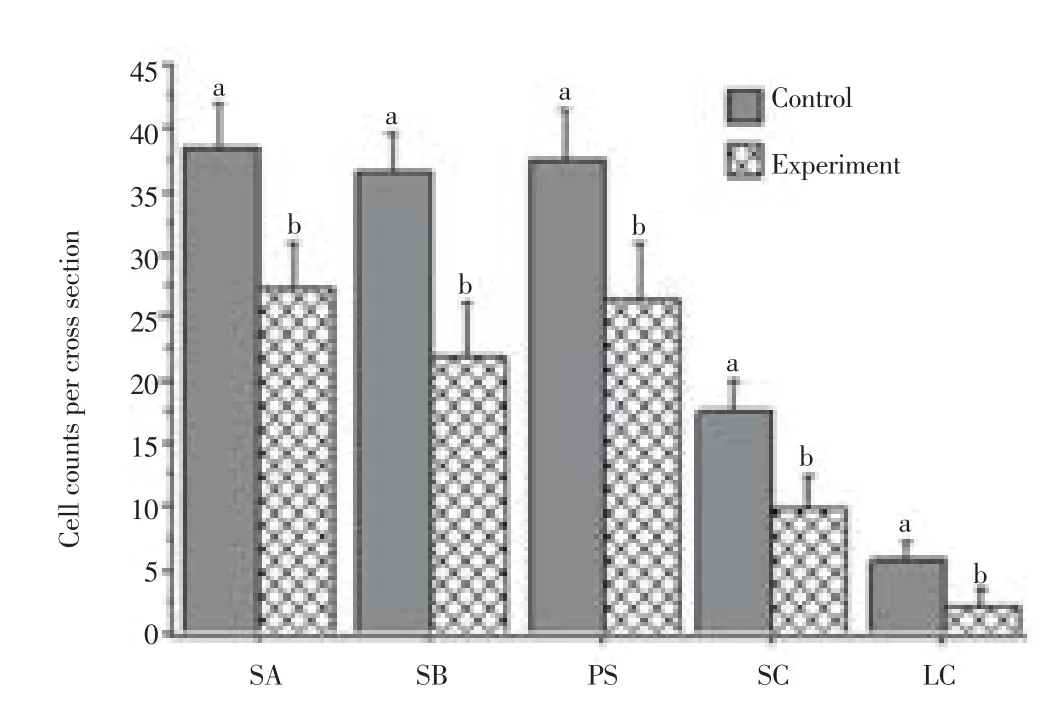
Figure 6. Comparison of siminiferous tubule cell counts.
4. Discussion
Morphometric, histologic examination and cell quantitation revealed that the testes of rats borne to dams treat with HPH had undergone diff erent gross and histological abnormal changes. The testis weight and dimensions are lesser than those of the controls. The ST appeared necrotic with folded basal laminae and they lost their usual configuration. The GE and SC suffered degenerative changes and lost their order. The diameter of the ST, thickness of the GE, luminal spermatozoa and the LC count were also aff ected.
These adverse eff ects are probably due to the eff ects of HPH and/ or its metabolites. This fi ndings support that of O’Donnell et al.[21] and Abney[22]. Also this fi nding agrees with that of Harini et al.[12] and Pushpalatha et al.[13]. The rats born to females treated with HPH in this study suff ered degeneration of SC and reduction in the number of LC. Proper male gonads development during prenatal life depends mainly on diff erentiation and development of LC and SC, which are necessary for the development of reproductive tract[23-25]. The fetal LC produce androgens during prenatal life[26] to develop the masculine characteristics of males[3] and SC; the first to be identifi ed in fetal testis; are the nurses of spermatogenesis and play an important role in development of the ST and LC[26,27]. Since in this study SC are seriously aff ected and LC number is reduced the diff erentiation and maturation of the GE is aff ected[28]. The main mission of SC is to orchestrate the epithelium germination through sequential phases of mitosis, meiosis, and differentiation. This mission is accomplished by providing hormonal, nutritional, and physical support
The SC of the experimental rats had undergone apoptotic changes such as cytoplasmic vacuoles and numerous lipid droplets. A similar phenomenon that aff ected spermatogenesis and sperm parameters was reported as a result of exogenous stimulant[29]. Furthermore empty vacuolar spaces were observed between SC (Places where spermatogonia and spermatocytes should germinate). The apoptotic changes observed in the GE agree with the fi ndings of Blanco et al.[30] who described apoptosis in hamster testis following treatment with anabolic androgenic steroids. Apoptosis of germ cells that occurs in the testicular epithelium serves as a mechanism to reduce the germ cell population to the level that the SC can support. Some drugs and toxic agents injure or disrupt the function of SC and reduce their supportive role, resulting in increment of the apoptotic elimination process[31].
The detachment of germ cells and miss-location of spermatids as well as spermatozoa to positions that are closely related to the basement membrane are probably due to disruption of SC-germ cell interactions a phenomenon reported by Richburg[31].
Toxic agents enhance production of reactive oxygen species (ROS) in cells & tissues these ROS exert oxidative stress (OS), which in turn increases the rates of cellular damage[32]. It seems that HP and/ or its metabolites; which can reach the fetal circuit[33]; have the capability to reach the fetal testes and induce similar oxidative stress. These oxidative stresses aff ect the testicular antioxidant system and lipid peroxidation leading to these cellular changes[34].
The decrement of spermatocytes and increment of the lumen of the ST in the present study are consistent with the study of Goyal et al.,[8]. The decrements in testicular weights and dimensions recorded in this study could be attributed to germinal and somatic cell loss, ST shrinkage and/or hypotrophy due to prenatal exposure to HP. The weight of the testis is largely dependent on the mass of diff erentiated spermatogenic cells and it has been used as a measure of spermatogenesis in rats[35]. A positive correlation was observed between weight of testis and number of germ cells[36]. Since the epididymis is the storehouse of sperms the decrement in its weight is presumably due to low sperm production. Similar fi ndings were reported by Pushpalatha et al.[13] and Harini et al.[12].
In conclusion, prenatal hydroxyprogesterone hexanoate exposure of male rats seriously aff ects their testicular morphometry, histology and cells quantities and presumably aff ects their future fecundity.
Declare of interest statement
We declare that we have no confl ict of interest.
[1] Carlsen E, Giwercman A, Keiding N, Skakkebaek NE. Evidence for decreasing quality of semen during past 50 years. Br Med J 1992; 304: 609-613.
[2] Pushpalatha T, Ramachandra Reddy P, Sreenivasula Reddy P. Eff ect of prenatal exposure to hydroxyprogesterone on steroidogenic enzymes in male rats. Naturwissenschaften 2002; 90: 40-43.
[3] Knez J. Endocrine-disrupting chemicals and male reproductive health. Reprod Biomed 2013; 26: 440-448.
[4] Pushpalatha T, Ramachandra Reddy P, Sreenivasula Reddy P. Gestational exposure to hydroxyprogesterone caproate suppresses reproductive potential in male rats. Naturwissenschaften 2005, 92: 385-388.
[5] G´eraldine D, Levacher C, Rene´ H. Estrogen effects on fetal and neonatal testicular development. Reproduction 2006; 132: 527-538.
[6] Storgaard L, Bonde JP, Olsen J. Male reproductive disorders in humans and prenatal indicators of estrogen exposure. A review of published epidemiological studies. Reprod Toxicol 2006; 21: 4-15.
[7] Fielden MR, Halgren RG, Fong CJ, Staub C, Johnson L, Chou K, et al. Gestational and lactational exposure of male mice to diethylstilbestrol causes long-term eff ects on the testis sperm fertilizing ability in vitro and testicular gene expression. Endocrinology 2002; 143: 3044-3059.
[8] Goyal HO, Robateau A, Braden TD, Williams CS, Srivastava KK, Ali KK. Neonatal estrogen exposure of male rats alters reproductive functions at adulthood. Biol Reprod 2003; 68: 2081-2091.
[9] Palmlund A, Pfel R, Buitendijk S, Cabau A, Forsberg JG. Effect of diethylstilbestrol (DES) medication during pregnancy. J Psychosom Obstet Gynaecol 1993; 14: 71-89.
[10] Vidaeff AC, Belfort MA. Critical appraisal of the efficacy, safety, and patient acceptability of hydroxyprogesterone caproate injection to reduce the risk of preterm birth. Patient Preference and Adherence 2013; 7: 683-691.
[11] Marianowski P, Radwańska E. Intramuscular vs vaginal progesterone for luteal support in cycles of in vitro fertilization. Ginekol Pol 2000; 71: 1064-1070.
[12] Harini C, Sainath SB, Sreenivasula Reddy P. Recovery of suppressed male reproduction in mice exposed to progesterone during embryonic development by testosterone. Reproduction 2009; 137: 439-448.
[13] Pushpalatha T, Ramachandra Reddy P, Trivikram G, Sreenivasula Reddy P. Reduced spermatogenesis in rats exposed trans-placently to hydroxyprogesterone. Cytologia 2003; 68: 369-373.
[14] Diamanti-Kandarakis E, Bourguignon JP, Giudice LC, Hauser R, Prins GS, Soto AM, et al. Endocrine-disrupting chemicals. Endocr Rev 2009; 30: 293-342.
[15] Pushpalatha T, Ramachandra Reddy P, Sreenivasula Reddy P. Impairment of male reproduction in adult rats exposed to hydroxyl-progesterone caproate in utero. Naturwissenschaften 2004; 91: 242-244.
[16] Bancroft JD. Fixation and fixatives. In: Theory and practice of histological techniques. 5th Ed. New York, London, San Francisco, Tokyo: Churchill Livingstone; 2003: 76.
[17] Thienpot D, Rochelle F, Vanpariis OFJ. Diagnosing Helmin-thiasis by Corprological Examination. 2nd Ed. Beerse: Janssen Foundation; 1986: 26-27.
[18] Batra N, Nehru B, Bansal MP. Influence of lead and zinc on rat male reproduction at biochemical and histopathological level. J Appl Toxicol 2001; 21: 507-512.
[19] Heller G, Leach GDR. Quantitation of Leydig cells and measurement of Leydig cell size following administration of human chorionic gonadotrophin to normal men. J Reprod Fertil 1971; 25: 185-192.
[20] Dykes J. Histometric assessment of human testicular biopsies. J Pathol Bacteriol 2001; 197: 429.
[21] O’Donnell L, Robertson KM, Jones M E, Simpson ER. Estrogen and spermatogenesis. Endocr Rev 2001; 22: 289-318.
[22] Abney TO. The potential roles of estrogens in regulating Leydig cell development and function: a review. Steroids 1999; 64: 610-617.
[23] Lejeune H, Habert R, Saez JM. Origin, proliferation and diff erentiation of Leydig cells. J Mol Endocrinol 1998; 20: 1-25.
[24] Magre S, Jost A. Sertoli cells and testicular diff erentiation in the rat fetus. J Electron Microsc Tech 1991; 19: 172-188.
[25] Yu M, Wang J, Liu W, Qin J, Zhou Q, Wang Y. Effects of tamoxifen on the sex determination gene and the activation of sex reversal in the developing gonad of mice. Toxicology 2014; 321: 89-95.
[26] Svechnikov K, Stukenborg JB, Savchuck I, Soder O. Similar causes of various reproductive disorders in early life. Asian J Androl 2014; 16: 50-59.
[27] Sharpe RM, McKinnell C, Kivlin C, Fisher JS. Proliferation and functional maturation of Sertoli cells and their relevance to disorders of testis function in adulthood. Reproduction 2003; 125: 769-784.
[28] Manivannan B, Mittal R, Goyal S, Ansary AS, Lohiya NK. Sperm characteristics and ultrastructure of testes of rat after long-term treatment with the methanol subfraction of carica papaya seeds. Asian J Androl 2009; 11: 583-599.
[29] Gopalkrishnan K, Gill-Sharma MK, Balasinor N, Padwal V, D’Souza S, Parte SP, et al. Tamoxifen-induced light and electron microscopic changes in the rat testicular morphology and serum hormonal profi le of reproductive hormones. Contraception 1998; 57: 261-269.
[30] Blanco A, Flores-Acuna F, Roldan-villalobos R, Monterde JG. Testicular damage from anabolic treatments with the beta (2)-adrenergic agonist clenbuterol in pigs: a light and electron microscope study. Vet J 2002; 163: 292-298.
[31] Richburg JH. The relevance of spontaneous- and chemically-induced alterations in testicular germ cell apoptosis to toxicology. Toxicol Lett 2000; 112-113: 79-86.
[32] Davies KJ, Quintanilha AT, Brooks GA, Packer L. Free radicals and tissue damage produced by exercise. Biochem Biophys Res Commun 1982; 107: 1198-1205.
[33] Hemauer SJ, Yan R, Patrikeeva SL, Mattison DR, Hankins GDV, Ahmed MS, et al. Transplacental transfer and metabolism of 17-alphahydroxyprogesterone caproate. Am J Obstet Gyneco 2008; 199: 169-170. [34] Husain K, Somani SM. Interaction of exercise training and chronic ethanol ingestion on testicular antioxidant system in rat. J Appl Toxicol 1998; 18: 421-429.
[35] Kavlock RJ, Daston GP, De Rosa C, Fenner CP, GrayKattari LE, Lucier SG, et al. Research needs for the risk assessment of health and environmental eff ect of endocrine disruptors: A report of the USEPA sponsored workshop. Env Hlth Perspect 1996; 104: 714-740.
[36] Schlappack OK, Delic JI, Harwood JR, Stanley JA. Protection from radiation-induced damage to spermatogenesis in the androgen pre-treated rats. Radiother Oncol 1988; 12: 219-224.
ent heading
10.1016/j.apjr.2016.04.015
*Corresponding author: Adil Salim Elsheikh, Department of Applied Medical sciences, Community College Najran, KSA.
E-mail: adilelgarrai@yahoo.com
Hydroxyprogesterone hexanoate Rat
Testis histology
 Asian Pacific Journal of Reproduction2016年3期
Asian Pacific Journal of Reproduction2016年3期
- Asian Pacific Journal of Reproduction的其它文章
- A rare cause of infertility: A late complication of female genital mutilation
- Peroxisome proliferator activated receptor gamma polymorphism Pro12 Ala in polycystic ovary syndrome (PCOS) of South Indian Population
- A new looming of Zika virus
- Effect of polycystic ovaries on in vitro fertilisation and intra-cytoplasmic sperm injection treatment outcome
- Alteration in oestus cycle and implantation in Mus musculus administered aqueous wood ash extract of Azadirachta indica (neem)
- Ameliorative potentials of quercetin against cotinine-induced toxic effects on human spermatozoa