
Prenatal corticosterone altered glucocorticoid receptor and glucocorticoid metabolic enzyme gene expression in chicken ovary
2016-11-30 12:33AbdelkareemAhmedAmalSifaldinHassanMusaTahaMusaJaafarFedail
Abdelkareem A. Ahmed, Amal Z. Sifaldin, Hassan H. Musa, Taha H. Musa, Jaafar S. Fedail
1Department of Physiology and Biochemistry, Faculty of Veterinary Sciences, University of Nyala, Nyala, Sudan
2Department of Microbiology, Faculty of Medical Laboratory Sciences, University of Khartoum, Khartoum, Sudan
3Key Laboratory of Environmental Medicine, Ministry of Education, School of Public Health, Southeast University, Nanjing 210009, Jiangsu, China
4Department of Biology, Faculty of Education, University of Nyala, Sudan
Prenatal corticosterone altered glucocorticoid receptor and glucocorticoid metabolic enzyme gene expression in chicken ovary
Abdelkareem A. Ahmed1*, Amal Z. Sifaldin1, Hassan H. Musa2, Taha H. Musa3, Jaafar S. Fedail4
1Department of Physiology and Biochemistry, Faculty of Veterinary Sciences, University of Nyala, Nyala, Sudan
2Department of Microbiology, Faculty of Medical Laboratory Sciences, University of Khartoum, Khartoum, Sudan
3Key Laboratory of Environmental Medicine, Ministry of Education, School of Public Health, Southeast University, Nanjing 210009, Jiangsu, China
4Department of Biology, Faculty of Education, University of Nyala, Sudan
ARTICLE INFO
Article history:
Received
Received in revised form
Accepted
Available online
Corticosterone
Objective: The acute stress response is an adaptive physiological mechanism which allows an organism to respond and survive deleterious stimuli in the surrounding environment. In mammals, prenatal glucocorticoids exposure (GCs) reprograms of fspring phenotype and reproductive performance. In the present study, we investigated potential prenatal GC exposure on the glucocorticoid receptor (GR), mineralocorticoid receptor (MR) and GC metabolic genes mRNA expression in the ovary of chickens. Methods: We injected low (0.2 μg) and high (1.0 μg) doses of corticosterone (CORT) in ovo before incubation and measured the changes in GCs metabolic enzymes genes in ovarian follicles 1 (F1), F2 and F3 post hatching. Results: The high dose CORT treatment signif icantly (P < 0.0) decreased 11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1) mRNA expression in F1, F2, F3 and in the ovary compared to the control and low groups. However, the high dose CORT treatment signifi cantly (P < 0.0) increased 11β-hydroxysteroid dehydrogenase type 2 (11β-HSD2) mRNA expressions in F1, F2, F3 and in the ovary compared to control and low groups. Likewise, in ovo injection of high dose CORT signifi cantly (P < 0.0) decreased 20-hydroxysteroid dehydrogenase (20-HSD) mRNA expression in F2, F3 and ovary compared to the control and low groups. Moreover, CORT treatment reduced GR mRNA expression in F1, F2 and F3 but not ovary. CORT treatment decreased MR mRNA only in F2. Conclusions: Prenatal CORT exposure modif ied GR, MR and GC metabolic enzymes gene expression in ovarian follicles, thus it may reprogram reproductive function.
1. Introduction
Not only genetic factor controls the phenotype and reproductive function of an organisms but also environmental factors can play a critical role in shaping off spring morphology[1] and reprogram reproductive performance[2]. In avian species, maternal infl uences have attracted much attention after the discovery that avian eggs contain a variety of steroid hormones[3,4]. Maternal derived steroid hormones are considered as a tool to adjust offspring phenotype[5]. Corticosterone (CORT) is considered the predominant glucocorticoid (GC) in the plasma of avian species, and it has been reported to deposit in the eggs of domestic chickens[6]. Eggs CORT concentrations is modulated by several factors including physiological status of the hen[7], the environment[8] such as housing conditions[9] and artifi cial elevation of egg CORT[10].
In chickens, reproductive capacity was found to be re-programmed by prenatal CORT exposure[11]. In birds, maternal stress modulates reproductive hormone concentrations in the eggs[12] and therefore aff ects off spring phenotype[13] and behavior[14]. The majority of studies investigating the eff ect of elevated CORT in the eggs have focused on growth rate[8] and behavior[15]. However, little is known about the eff ects of embryonic CORT exposure on reproductive capacity in avian species. Moreover, it is still unknown the eff ect of prenatal CORT exposure on stress related gene expression in the ovary. The action of GCs on cells is mediated via glucocorticoid receptor (GR), and mineralocorticoid receptor (MR)[16]. The intracellular concentrations of active GC are under control of a number of metabolizing enzymes which is called pre-receptormodulation[17]. The 11β-hydroxysteroid dehydrogenase (11β-HSD1) activates, while 11β-hydroxysteroid dehydrogenase (11β-HSD2) deactivates GCs[18-22]. In avian species, 20-hydroxysteroid dehydrogenase (20-HSD) is an abundantly and ubiquitously expressed enzyme, which transforms GCs to inactive 20-dihydrocorticosterone[23]. The ovary is well equipped with local regulatory mechanisms of GC action[24]. The major regulatory system consists of changes in the expression of the two isoforms of 11β-HSDs that catalyze the inter-conversion of GCs in the ovary[25,26]. Yet, the effects of prenatal CORT exposure on GCs metabolic enzymes gene expression in chicken ovary remain less clear. Therefore, here we used a model of in ovo injection of CORT prior to incubation to examine our hypothesis that expression of genes involved in GC metabolism in the ovaries may be modifi ed by embryonic CORT exposure.
2. Materials and methods
2.1. Egg incubation and CORT injection
Two hundred and ten fertilized chicken eggs were selected from eggs laid by hens and divided into three groups (70 in each group). CORT (Sigma-Aldrich, USA) was dissolved in sesame oil and diluted in PBS to make doses of 0.2 μg and 1.0 μg in a volume of 100 μL solution. The high and low CORT dose was selected based on earlier publications[27,28]. Before incubation, the eggs were injected with PBS (control) and a 0.2 μg (low) or a 1.0 μg (high) dose of CORT under aseptic conditions. Eggs were injected by advancing a Hamilton syringe into a hole in the middle of the long axis until the yolk membrane was penetrated. The incubation condition was set based on our previous publication[29]. Chicks hatched inside the incubator and were left to dry completely (up to 12 h) before they were removed. Day-old chicks were individually wing banded, and placed into battery cages with 12 h fl uorescent lighting and 12 h dark. The temperature was adjusted to 32–35 °C during the fi rst week, and reduced approximately 3 °C per week until 21 °C. Both sexes were transferred to floor pens covered with sawdust litter. The stocking density was 20–25 kg/m2. The relative humidity was maintained at 40%–60%, and the lighting, ventilation, as well as the feeding procedures complied. On week 35, all chickens were killed by rapid decapitation. The ovaries were collected and weighed. The ovarian follicles were collected, washed with PBS then put in liquid nitrogen and later kept at-80 °C for further analysis. The experiment procedures were approved by the Animal Ethics Committee of Nanjing Agricultural University.
2.2. RNA extraction and mRNA quantification with Realtime PCR
Ovary and ovarian follicle were ground with pestle and mortar in liquid N2 and a portion of approximately 100 mg was used for the RNA extraction using the TRIzol total RNA kit (Invitrogen, Biotechnology Co, Ltd, Carlsbad, CA, USA) according to the manufacturer’s instructions, and reverse transcript to cDNA using 0.5 μg/μL (4 μL contains 4 μg) of RNA with the PrimeScript RT reagent kit according to the manufacturers instruction (RNase Free, D2215, Takara, Japan). To investigate the effect of the in ovo injection of CORT on the expression of hypothalamic genes, real-time PCR was performed in an Mx3000P (Stratagene, USA) according to published methods[30]. Mock RT and No Template Controls (NTC) were included to monitor the possible contamination of genomic and environmental DNA at the RT and PCR steps. A pooled sample made by mixing equal quantities of the RT products (cDNA) from all the samples was used for optimizing the PCR conditions and tailoring the standard curves for each target gene, and melting curves were performed to insure a single specifi c PCR product for each gene. The PCR products were sequenced to validate the identity of the amplicons. Primers specifi c for the 11β-HSD1, 11β-HSD2, 20-HSD, GR, and MR (Table 1) were synthesized by Geneary, Shanghai, China. Chicken β-actin was used as a reference gene for normalization purposes. The method of 2−ΔΔCt was used to analyze the real-time PCR data[31].
2.3. Statistical analysis
Descriptive statistics was performed to check the normality and homogeneity of variances before using parametric analyses. The relative quantitative data of gene expression were analyzed by oneway ANOVA using SPSS 16.0 for Windows, followed by a leastsignificant difference (LSD) test for individual comparisons. A P-value ≤ 0.05 was considered signifi cant.
3. Results
3.1. Ovary weight and oviduct weight
High dose of CORT treatment signifi cantly (P < 0.05) decreased ovary weight and oviduct weight compared to low and controlgroups. Low dose of CORT treatment did not aff ect ovary weight or oviduct weight compared to low and control groups (Table 2).
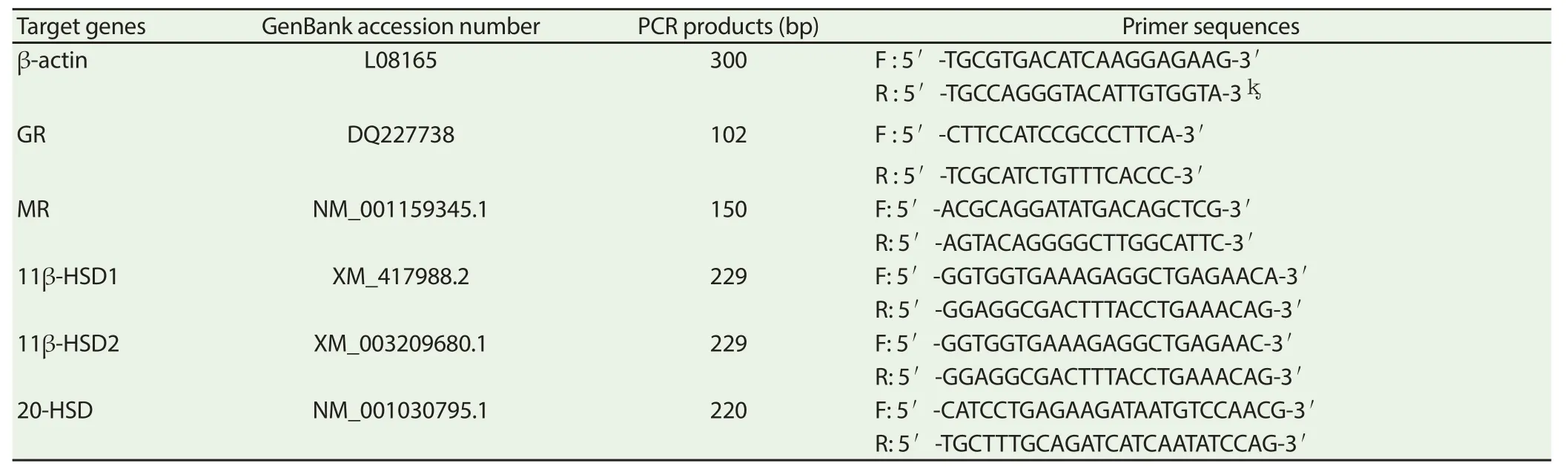
Table 1 Real-time PCR primers.
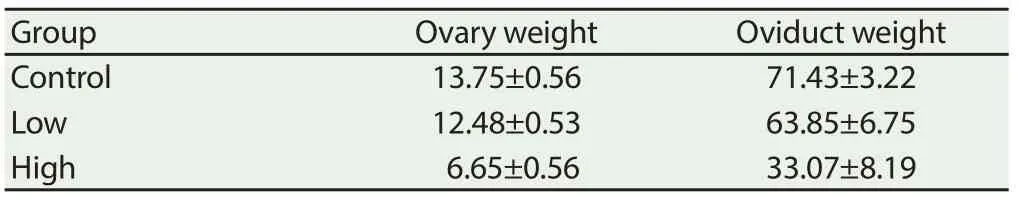
Table 2 Eff ect of in ovo injection of CORT on ovary weight and oviduct weight.
3.2. Ovarian follicles theca cells GC metabolic enzymes, GR and MR mRNA expression
CORT treatment signifi cantly decreased (P < 0.05) ovarian F1, F2, F3 and ovary 11β-HSD1 mRNA expression (Figure 1A). However, low dose of CORT did not alter 11β-HSD1 mRNA expression if F1. In contrast, high dose of CORT treatment significantly increased (P < 0.05) 11β-HSD2 mRNA F1, F2, F3 and ovary (Figure 1B), whereas, 20-HSD mRNA was downregulated in F2, F3 and ovary regarding CORT treatment (Figure 1C). Both low and high doses of CORT treatment significantly decreased (P < 0.05) GR mRNA expression in ovarian F2 and F3 but not F1 or ovary (Figure 1D). Likewise, low and high doses of CORT treatment decreased MR mRNA in F2 (Figure 1E).
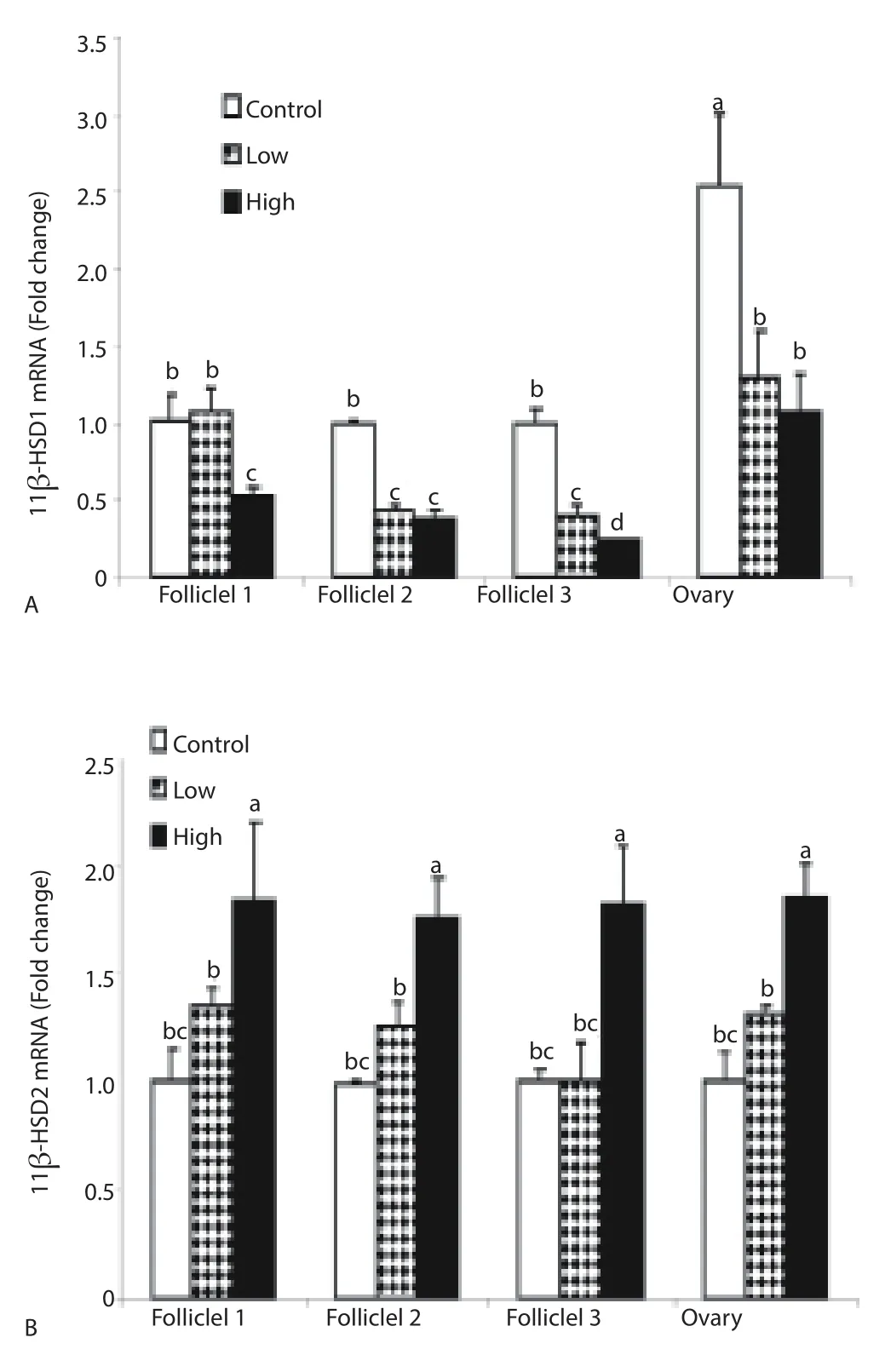
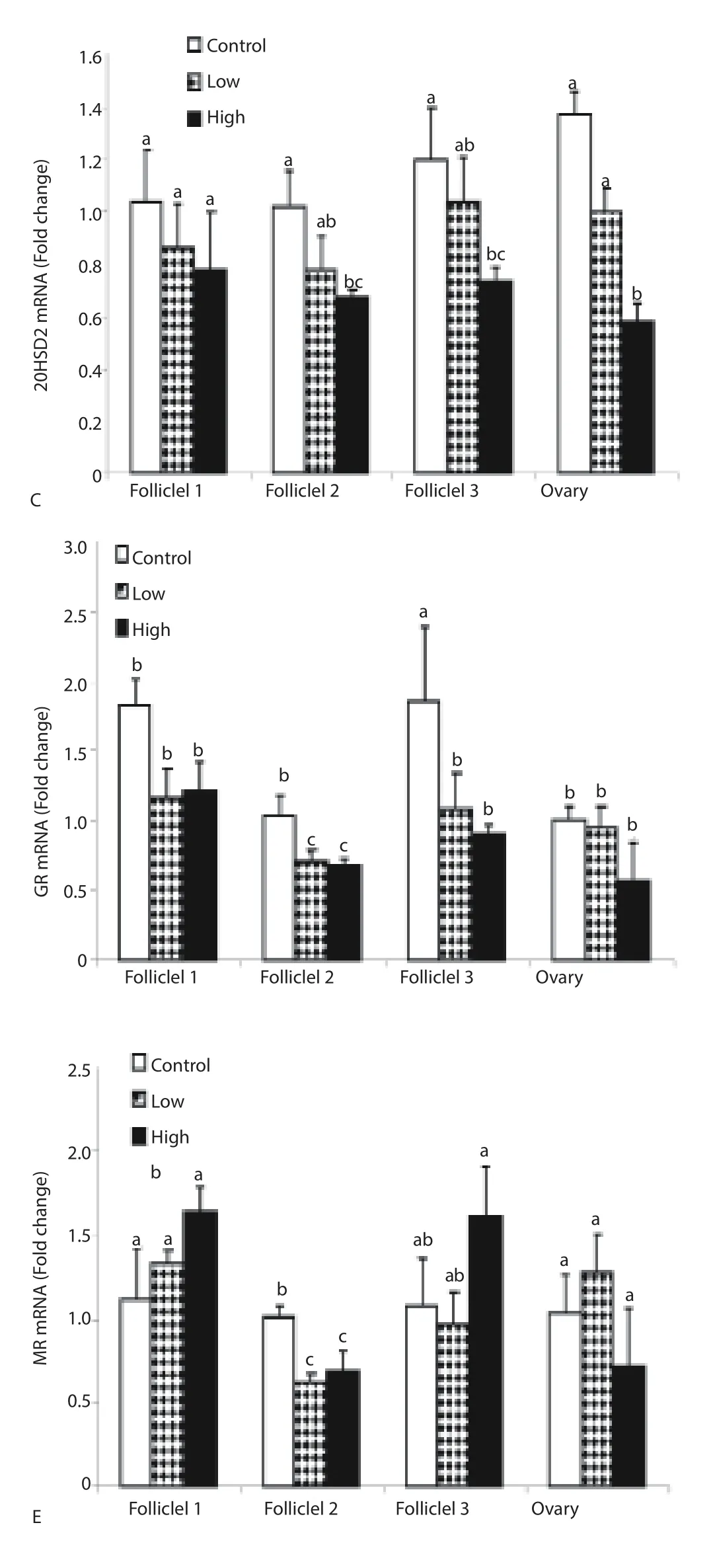
Figure 1. Effect of in ovo injection of CORT on ovarian follicles (F1, F2 and F3) and ovary 11β-HSD1 mRNA (A), 11β-HSD2 mRNA (B), 20-HSD mRNA (C), GR mRNA (D) and MR mRNA (E) expression.
4. Discussion
In mammals, early life experiences re-program the hypothalamic-pituitary-gonadal (HPG) axis, which is associated with increased reproductive dysfunction later in life (Heim and Nemeroff , 2001). We report here, for the first time, the effect of prenatal CORT exposure on GC metabolic enzymes gene expression in the ovarian follicles in chickens. In ovo administration of CORT decreased ovary weight and oviduct weight. These fi ndings are in agreement with previous studies that prenatal stress has been reported to decrease ovary weight in guinea pig off spring[32].
Previous reports indicated that maternal adversity during the prenatal period altered 11β-HSDs expression in humans[33,34]. Here we reported for the first time the effect of CORT in ovo on 11β-HSDs in the ovary. CORT treatment decreased ovarian follicles 11β-HSD1 and 20-HSD mRNA expression whereas increased 11β-HSD2 mRNA expression in ovarian follicles. Earlier studies indicated that upregulation of 11β-HSD1 may follow prenatal stress hormone exposure which is considered a novel mechanism for fetal origins of adult metabolic syndrome[35], whereas 11B-HSD2 appears/is involved in fetal programming of behavior in mice[36]. In human ovaries, the simultaneous upregulation of 11β-HSD1 and downregulation of 11β-HSD2[37] induced luteinized granulose cells to be rich with 11β-reductase activity[38], which most probably explains why both total and free cortisol levels in human follicular fl uid become high after the ovulation inducing LH surge[39]. The enzymes 11β-HSDs, are the main GC activation/inactivation enzymes in the placenta of mammals, has also been reported in the oviduct and ovary of chickens[40] and in the ovary of zebra fi nches[41]. The alteration of these enzymes mRNA might explain the mechanism of low transferring rate of CORT from the maternal plasma to her eggs and thus suggests that mammals and birds have similar strategies to protect their embryos from overexposure to maternal CORT.
In addition, CORT treatment decreased GR receptor mRNA and MR receptor mRNA levels in theca layer of ovarian follicles. These fi ndings are in agreement with previous fi ndings that exposure to dexamethasone decreased GR mRNA levels in hamster ovarian cell line[42]. Because the current study is the fi rst to reveal the eff ects of prenatal CORT exposure on changes in GR and MR mRNA expression in the ovary of domestic fowl, we will consider the potential implications of these results. In mammals and humans, disruption of GR/MR stability is associated with several health disorders, such as post-traumatic stress disorder[43], depression[44], and anxiety[45]. In general, disturbances in the ratio of GR/MR after chronic stress increases the overall ‘wear and tear’ in the organism, frequently referred to as allostatic load[46]. Such disruption decreases resilience and reduces the ability of an organism to respond appropriately to acute stress[47]. Further studies are required to elucidate whether these transcriptional changes are permanent or transient.
In conclusion, we demonstrate that embryonic exposure to CORT decreased oviduct and ovary weight associated with alterations of GR, MR and glucocorticoid metabolic enzymes genes expression in chicken ovary.
Declare of interest statement
We declare that we have no confl ict of interest.
Acknowledgments
The authors are highly grateful to Professor Zhao Ruqian at the Nanjing Agricultural University, for her support to conduct this research.
[1] Nijhout HF. Development and evolution of adaptive polyphenisms. Evol Dev 2003; 5: 9-18.
[2] Jr AMD, Clobert J, Møller AP. Hormones, developmental plasticity and adaptation. Trends Ecol Evol 2002; 17: 190-196.
[3] Groothuis TG, Muller W, von Engelhardt N, Carere, Eising C. Maternal hormones as a tool to adjust offspring phenotype in avian species. Neurosci Biobehav Rev 2005; 29: 329-352.
[4] Schwabl H. Yolk is a source of maternal testosterone for developing birds. Proc Natl Acad Sci U S A 1993; 90: 11446-11450.
[5] Meylan S, Miles DB, Clobert J. Hormonally mediated maternal eff ects, individual strategy and global change. Philos Trans R Soc Lond B Biol Sci 2012; 367: 1647-1664.
[6] Rettenbacher S, Mostl E, Groothuis TG. Gestagens and glucocorticoids in chicken eggs. Gen Comp Endocrinol 2009; 164: 125-129.
[7] Saino N, Romano M, Ferrari RP, Martinelli R, Moller AP. Stressed mothers lay eggs with high corticosterone levels which produce lowquality off spring. J Exp Zool A Comp Exp Biol 2005; 303: 998-1006.
[8] Hayward LS, Wingfield JC. Maternal corticosterone is transferred to avian yolk and may alter off spring growth and adult phenotype. Gen Comp Endocrinol 2004; 135: 365-371.
[9] Lay DC Jr, Fulton RM, Hester PY, Karcher DM, Kjaer JB, Mench JA, et al. Hen welfare in diff erent housing systems. Poult Sci 2011; 90: 278-294.
[10] Ahmed AA, Ma W, Ni Y, Wang S, Zhao R. Corticosterone in ovo modifi es aggressive behaviors and reproductive performances through alterations of the hypothalamic-pituitary-gonadal axis in the chicken. Anim Reprod Sci 2014; 146: 193-201.
[11] Dunn E, Kapoor A, Leen J, Matthews SG. Prenatal synthetic glucocorticoid exposure alters hypothalamic-pituitary-adrenal regulation and pregnancy outcomes in mature female guinea pigs. J Physiol 2010; 588: 887-899.
[12] Henriksen R, Groothuis TG, Rettenbacher S. Elevated plasma corticosterone decreases yolk testosterone and progesterone in chickens: linking maternal stress and hormone-mediated maternal eff ects. PLoS One 2011; 6: e23824.
[13] Gil D. Golden eggs: Maternal manipulation of off spring phenotype by egg androgen in birds. Ardeola 2003; 50: 281-294.
[14] Riedstra B, Pfannkuche K, Groothuis TGG. Increased exposure to yolk testosterone has feminizing eff ects in chicken, Gallus gallus domesticus. Animal Behaviour 2013; 85: 701-708.
[15] Henriksen R, Rettenbacher S, Groothuis TGG.. Prenatal stress in birds: Pathways, eff ects, function and perspectives. Neurosci Biobehav R 2011; 35: 1484-1501.
[16] Tsugita M, Iwasaki Y, Nishiyama M, Taguchi T, Shinahara M, Taniguchi Y, et al. Glucocorticoid receptor plays an indispensable role in mineralocorticoid receptor-dependent transcription in GR-defi cient BE(2)C and T84 cells in vitro. Mol Cell Endocrinol 2009; 302: 18-25.
[17] Edwards CR, Benediktsson R, Lindsay RS, Seckl JR. 11 beta-Hydroxysteroid dehydrogenases: key enzymes in determining tissuespecifi c glucocorticoid eff ects. Steroids 1996; 61: 263-269.
[18] Diederich S, Hanke B, Burkhardt P, Muller M, Schoneshofer M, Bahr V, et al. Metabolism of synthetic corticosteroids by 11 beta-hydroxysteroiddehydrogenases in man. Steroids 1998; 63: 271-277.
[19] Harris HJ, Kotelevtsev Y, Mullins JJ, Seckl JR, Holmes MC. Intracellular regeneration of glucocorticoids by 11beta-hydroxysteroid dehydrogenase (11beta-HSD)-1 plays a key role in regulation of the hypothalamicpituitary-adrenal axis: analysis of 11beta-HSD-1-deficient mice. Endocrinology 2001; 142: 114-120.
[20] Holmes MC, Seckl JR. The role of 11beta-hydroxysteroid dehydrogenases in the brain. Mol Cell Endocrinol 2006; 248: 9-14.
[21] Holmes M., Yau JL, Kotelevtsev Y, Mullins JJ, Seckl JR. 11 Betahydroxysteroid dehydrogenases in the brain: two enzymes two roles. Ann N Y Acad Sci 2003; 1007: 357-366.
[22] Stewart PM, Krozowski ZS. 11 beta-Hydroxysteroid dehydrogenase. Vitam Horm 1999; 57: 249-324.
[23] Kucka M, Vagnerova K, Klusonova P, Miksik I, Pacha J. Corticosterone metabolism in chicken tissues: evidence for tissue-specifi c distribution of steroid dehydrogenases. Gen Comp Endocrinol 2006; 147: 377-383.
[24] Whirledge S, Cidlowski JA. Glucocorticoids, stress, and fertility. Minerva Endocrinol 2010; 35: 109-125.
[25] Webb RJ, Sunak N, Wren L, Michael AE. Inactivation of glucocorticoids by 11beta-hydroxysteroid dehydrogenase enzymes increases during the meiotic maturation of porcine oocytes. Reproduction 2008; 136: 725-732. [26] Seckl JR, Walker BR. Minireview: 11β-hydroxysteroid dehydrogenase type 1– a tissue-specific amplifier of glucocorticoid action. Endocrinology 2001; 142: 1371-1376.
[27] Heiblum R, Arnon E, Chazan G, Robinzon B, Gvaryahu G, Snapir N. Glucocorticoid administration during incubation: embryo mortality and posthatch growth in chickens. Poult Sci 2001; 80: 1357-1363.
[28] Haussmann MF, Longenecker AS, Marchetto NM, Juliano SA, Bowden RM. Embryonic exposure to corticosterone modifi es the juvenile stress response, oxidative stress and telomere length. Proc Biol Sci 2012; 279: 1447-1456.
[29] Ahmed AA, Ma W, Ni Y, Zhou Q, Zhao R. Embryonic exposure to corticosterone modifi es aggressive behavior through alterations of the hypothalamic pituitary adrenal axis and the serotonergic system in the chicken. Hormones and Behavior 2014; 65: 97-105.
[30] Ahmed AA, Ma W, Ni Y, Zhou Q, Zhao R. Embryonic exposure to corticosterone modifi es aggressive behavior through alterations of the hypothalamic pituitary adrenal axis and the serotonergic system in the chicken. Horm Behav 2014; 65: 97-105.
[31] Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001; 25: 402-408.
[32] Kapoor A, Matthews SG. Prenatal stress modifies behavior and hypothalamic-pituitary-adrenal function in female guinea pig off spring: effects of timing of prenatal stress and stage of reproductive cycle. Endocrinology 2008; 149: 6406-6415.
[33] Jensen Pena C, Monk C, Champagne FA. Epigenetic eff ects of prenatal stress on 11beta-hydroxysteroid dehydrogenase-2 in the placenta and fetal brain. PLoS One 2012; 7: e39791.
[34] Lucassen PJ, Bosch OJ, Jousma E, Krömer SA, Andrew R, Seckl JR, et al. Prenatal stress reduces postnatal neurogenesis in rats selectively bred for high, but not low, anxiety: possible key role of placental 11betahydroxysteroid dehydrogenase type 2. Eur J Neurosci 2009; 29: 97-103.
[35] Nyirenda MJ, Carter R, Tang JI, de Vries A, Schlumbohm C, Hillier SG, et al. Prenatal programming of metabolic syndrome in the common marmoset is associated with increased expression of 11betahydroxysteroid dehydrogenase type 1. Diabetes 2009; 58: 2873-2879.
[36] Holmes MC, Abrahamsen CT, French KL, Paterson JM, Mullins JJ, Seckl JR. The mother or the fetus? 11beta-hydroxysteroid dehydrogenase type 2 null mice provide evidence for direct fetal programming of behavior by endogenous glucocorticoids. J Neurosci 2006; 26: 3840-3844.
[37] Tetsuka M, Thomas FJ, Thomas MJ, Anderson RA, Mason JI, Hillier SG. Diff erential expression of messenger ribonucleic acids encoding 11betahydroxysteroid dehydrogenase types 1 and 2 in human granulosa cells. J Clin Endocrinol Metab 1997; 82: 2006-2009.
[38] Thomas FJ, Thomas MJ, Tetsuka M, Mason JI, Hillier SG. Corticosteroid metabolism in human granulosa-lutein cells. Clin Endocrinol (Oxf) 1998; 48: 509-513.
[39] Harlow CR, Jenkins JM, Winston RM. Increased follicular fluid total and free cortisol levels during the luteinizing hormone surge. Fertil Steril 1997; 68: 48-53.
[40] Klusonova P, Kucka M, Miksik I, Bryndova J, Pacha J. Chicken 11betahydroxysteroid dehydrogenase type 2: partial cloning and tissue distribution. Steroids 2008; 73: 348-355.
[41] Katz A, Oyama RK, Feng N, Chen X, Schlinger BA. 11betahydroxysteroid dehydrogenase type 2 in zebra fi nch brain and peripheral tissues. Gen Comp Endocrinol 2010; 166: 600-605.
[42] Bellingham DL, Sar M, Cidlowski JA. Ligand-dependent downregulation of stably transfected human glucocorticoid receptors is associated with the loss of functional glucocorticoid responsiveness. Mol Endocrinol 1992; 6: 2090-2102.
[43] de Kloet ER, Joels M, Holsboer F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci 2005; 6: 463-475.
[44] de Kloet ER, Derijk RH, Meijer OC. Therapy Insight: is there an imbalanced response of mineralocorticoid and glucocorticoid receptors in depression? Nat Clin Pract Endocrinol Metab 2007; 3: 168-179.
[45] Herrero AI, Sandi C, Venero C. Individual differences in anxiety trait are related to spatial learning abilities and hippocampal expression of mineralocorticoid receptors. Neurobiol Learn Mem 2006; 86: 150-159.
[46] McEwen BS. Protective and damaging eff ects of stress mediators: central role of the brain. Dialogues Clin Neurosci 2006; 8: 367-381.
[47] Joëls M, Karst H, DeRijk R, de Kloet ER. The coming out of the brain mineralocorticoid receptor. Trends Neurosci 2008; 31: 1-7.
ent heading
10.1016/j.apjr.2016.04.007
*Corresponding author: Dr. Abdelkareem Abdallah Ahmed, Department of Physiology and Biochemistry, Faculty of Veterinary Science, University of Nyala, 155 Nyala, Sudan.
E-mail: kareemo151@gmail.com, kareemo151@vet.nyalau.edu.sd
Fax: 00249711833123
Egg
Glucocorticoid metabolic enzymes Ovary
 Asian Pacific Journal of Reproduction2016年3期
Asian Pacific Journal of Reproduction2016年3期
- Asian Pacific Journal of Reproduction的其它文章
- A rare cause of infertility: A late complication of female genital mutilation
- Peroxisome proliferator activated receptor gamma polymorphism Pro12 Ala in polycystic ovary syndrome (PCOS) of South Indian Population
- A new looming of Zika virus
- Effect of polycystic ovaries on in vitro fertilisation and intra-cytoplasmic sperm injection treatment outcome
- Alteration in oestus cycle and implantation in Mus musculus administered aqueous wood ash extract of Azadirachta indica (neem)
- Ameliorative potentials of quercetin against cotinine-induced toxic effects on human spermatozoa