
2014年春季季风间期东印度洋赤道及其邻近海域硅藻群落
2016-11-30 05:35:35薛冰孙军丁昌玲王东晓
海洋学报 2016年2期
薛冰,孙军*,丁昌玲,王东晓
(1.天津科技大学海洋与环境学院,天津300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津300457;3.中国科学院南海海洋研究所热带海洋环境国家重点实验室,广东广州510301)
2014年春季季风间期东印度洋赤道及其邻近海域硅藻群落
薛冰1,2,孙军1,2*,丁昌玲1,2,王东晓3
(1.天津科技大学海洋与环境学院,天津300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津300457;3.中国科学院南海海洋研究所热带海洋环境国家重点实验室,广东广州510301)
2014年4月10日至5月13日在东印度洋赤道区及其邻近海域(10.08°N—6.00°S,80.00°~96.10°E)进行硅藻物种组成和群落结构的调查。分析了45个网采样品,共鉴定出浮游硅藻34属113种(包括变种、变形及未定名种),大部分物种为热带外洋性种以及暖海外洋性种,与该海区的热带及亚热带的环境特征一致。优势种为佛氏梯形藻(Climacodium frauenfeldianum)、地中海细柱藻(Leptocylindrus mediterraneus)、密聚角毛藻(Chaetoceros coarctatus)、美丽漂流藻(Planktoniella foromsa)、大西洋角毛藻那不勒斯变种(Chaetoceros atlanticus var.neapolitanus)、距端假管藻(Pseudosolenia calcar-avis,即距端根管藻Rhizosolenia calcaravis)、圆柱几内亚藻(Guinardia cylindrus)、达蒂角毛藻(Chaetoceros dadayi)、伏氏海线藻(Thalassionema frauenfeldii)、离心列海链藻(Thalassiosira excentrica)、瘤面角毛藻(Chaetoceros bacteriastroides)以及笔尖根管藻粗径变种(Rhizosolenia styliformis var.latissima)等。硅藻的平均细胞丰度为1.855×103个/m3,其平面分布不均匀,赤道断面细胞丰度较低,高值区出现在海区北部。聚类分析发现有7种生态类群,这些硅藻的种类和丰度平面分布与上层水体(200 m以浅)温度、盐度及营养盐的水柱平均值有较好的对应关系。
硅藻;东印度洋;种类组成;平面分布;聚类分析
1 引言
硅藻是海洋生态系统中重要的初级生产者,硅藻提供的净初级生产约占全球年净初级生产的20%~25%[1],是食物链及食物网的主要启动者[2],也是海洋动物的直接或间接饵料,对海洋中生物资源的开发及利用有着重要的指示作用[3],在生态系统的物质循环与能量流动中起着重要的作用[4]。
印度洋是世界第三大洋,受欧亚大陆上空冷暖气团的影响,形成了独特的季风气候[5]。东印度洋主要的洋流有西南或东北季风漂流、东印度沿岸流以及南赤道逆流或潜流[6],Jagadeesan等[7]在印度东南岸的曼纳湾及保克海峡的研究表明环流会对中型浮游动物的群落结构产生影响,由此推断环流也会对浮游植物群落结构产生影响。春季由于赤道西风的出现,印度洋处于季风交替期,由东北季风逐渐转为西南季风[8]。东印度洋的初级生产量相比于西印度洋的初级生产量较低[9]。受纬度洋流、赤道洋流以及温跃层较浅的影响,赤道以北海区的营养盐含量要高于赤道以南海区的营养盐含量。
硅藻种群结构的变化可以在云反照率[10]、海水光通量及热通量[11]上改变全球气候,而印度洋的气候变化在一定程度上可以影响我国气候的变化[12]。对于印度洋硅藻种群结构的研究国内几乎为空白,国外也只有很少数的研究,如Tarran等[13]通过使用显微镜及流式细胞仪的方法对阿拉伯海的浮游植物群落结构进行了调查,Schlüter等[14]使用HPLC的方法对印度洋南部的浮游植物群落及组成进行了调查。
本文对2014年春季东印度洋硅藻种群结构特征做了初步分析,以期对东印度洋相关研究工作提供基础的研究资料,为以后更加深入细致的研究打下基础。
2 材料及方法
2.1调查站位
调查区域为印度洋东部(10.08°N—6.00°S,80.00°~96.10°E)的海域。2014年4月10日至5月13日乘“实验1号”船对该海域进行了生物、化学、地质等综合调查,共45个站位,其中包含赤道断面、与赤道平行及垂直断面以及与印度尼西亚平行断面,设置拖网站位如图1。
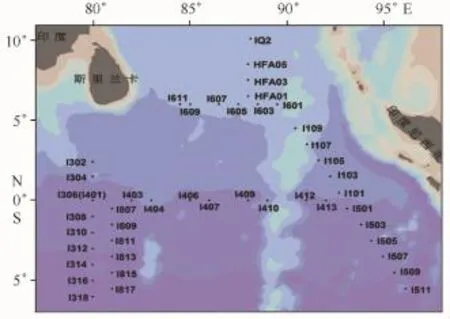
图1 东印度洋调查站位Fig.1 Sampling stations in eastern Indian Ocean
2.2采样及分析方法
按照《海洋调查规范》[15],采样工具为大洋型小型浮游生物网(网口直径为80 cm,网口面积为0.5 m2,网身长270 cm,网目为76μm),采样方式为在每个调查站位从底层至表层垂直拖网1次(水深超过200 m时,自200 m水深垂直拖网)。样品放入1 000 mL的PE瓶中,用甲醛溶液固定后置于阴凉处保存(甲醛体积终浓度为2%)。实验室内分析方法为:将浮游植物样品轻轻混匀,取2.97 m L浮游植物亚样品于Utermöhl型计数框中,在Motic AE2000型倒置显微镜200倍或400倍下进行浮游植物的物种鉴定和细胞计数。浮游植物的物种鉴定参照Tomas[16]。浮游植物的中文名及拉丁文名参照孙军和刘东艳[17]。本文所用的温度、盐度数据来自于Seabird CTD探头数据,营养盐数据参考World Ocean Atlas(WOA)13春季营养盐平均浓度值(http://www.nodc.noaa.gov/OC5/woa13/)。
2.3数据处理方法
浮游植物的优势种根据优势度的值(Y)来确定[18]:

式中,ni为第i种的细胞个数;N为所采集的样品中所有种的总细胞个数;fi为第i种在各样品中出现的频率。
3 结果
3.1海区环境特征
调查海区200 m以浅平均温度、盐度分布如图2a,b。调查期间,温度变化较大(20.73~24.88℃),平均值为23.22℃,盐度值介于33.9~35.1,平均值为34.7。在垂直分布上,温度一般随深度增加而减小,而盐度则呈相反趋势。由温度、盐度的水平分布图可以看出,调查海区温度、盐度分布不均匀,温度由北向南呈先降低再增加继而又降低的趋势,盐度为先增加再减小的趋势。海区北部与赤道垂直的断面为高温低盐的海水,而赤道断面为高温高盐的海水。
3.2硅藻的物种组成及平面分布
对本次调查获得的45个浮游植物网样初步分析,共发现硅藻34属113种(包括变种、变形及未定名种),其中种类最多的属为角毛藻属38种,占硅藻总种数的33.63%;其次为根管藻属13种,占硅藻总种数的11.50%;再次为圆筛藻属9种,占硅藻总种数的7.96%。本次调查发现硅藻的生态类型多为热带外洋性、暖海外洋性,说明该海区具有热带及亚热带气候特征。热带外洋性类型主要代表物种有丛毛辐杆藻(Bacteriastrum comosum)、密聚角毛藻(Chaetoc-eros coarctatus)、达蒂角毛藻(Chaetoceros dadayi)、短刺角毛藻(Chaetoceros messanense)、美丽漂流藻(Planktoniella foromsa)等;暖海外洋性类型主要代表物种有洛氏角毛藻(Chaetoceros lorenzianus)、热带顾氏藻(Gossleriella tropica)、霍氏半管藻(Hemiaulus hauckii)、地中海细柱藻(Leptocylindrus mediterraneus)、伯氏根管藻(Rhizosolenia bergonii)等;广温广布性类型主要代表物种有尖刺伪菱形藻(Pseudonitzschia pungens)、翼鼻状藻(Proboscia alata)、纤细型翼鼻状藻(Proboscia alata f.gracillima)、笔尖根管藻(Rhizosolenia styliformis)、菱形海线藻(Thalassionema nitzschiöides)等。
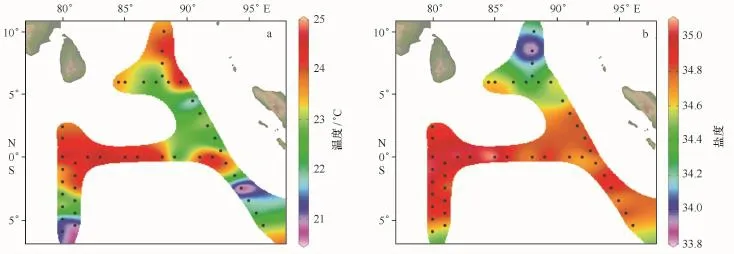
图2 调查海域200 m以浅平均温度和盐度的平面分布Fig.2 Horizontal distribution of average temperature and salinity above 200 m water depth in the survey area
硅藻的细胞丰度介于0.099×103~7.146×103个/m3,平均细胞丰度为1.855×103个/m3。硅藻平面分布如图3,分布趋势呈明显地斑块状分布,细胞丰度最大值出现于I605站(6.00°N,87.50°E),该站共出现44个种,主要物种有短刺角毛藻、地中海细柱藻、柔弱伪菱形藻、洛氏角毛藻、佛氏梯形藻、粗股角毛藻、丛毛辐杆藻等,热带外洋性及暖海外洋性物种居多;其次为与其相距不远的I609站(6.893×103个/m3)(6.00°N,85.02°E),该站共出现49个种,主要物种为佛氏梯形藻、柔弱伪菱形藻、丛毛辐杆藻、密聚角毛藻、洛氏角毛藻、霍氏半管藻等,大部分为热带外洋性及暖海外洋性物种。分布密集区主要位于这两个站出现的断面,即调查海区北部与赤道平行断面(平均细胞丰度为5.026×103个/m3),该断面温度值与盐度值居中,3种营养盐浓度都很高(图6);其次为西部与赤道垂直的两个断面(平均细胞丰度为1.994×103个/m3),赤道断面的细胞丰度较低(平均细胞丰度为0.475× 103个/m3),这可能受赤道断面的营养盐浓度较低的影响。硅藻分布具有更小尺度的水平分布特征。
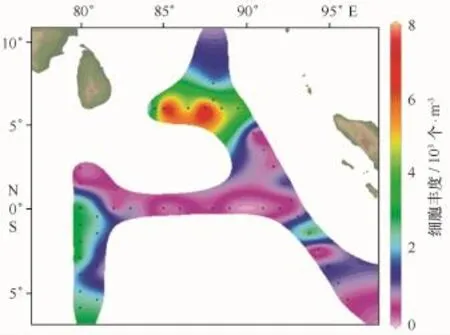
图3 东印度洋硅藻平面分布Fig.3 Horizontal distribution of diatoms in the eastern Indian Ocean
3.3优势种组成及平面分布
3.3.1聚类分析
对出现频率fi大于0.5的物种进行聚类分析,再结合优势度Y值确定出优势种,即从聚类分析结果中相类似的物种中根据优势度Y值的大小选出有代表性的优势种。聚类分析结果如图4,fi大于0.5的物种的丰度比例及优势度见表1。由图4可以看出,可以将30个物种分为7个类群,根据优势度Y的大小,分别从这7个类群中挑选出一定数量的物种作为优势种,按从左至右的顺序将这7个类群依次命名为Ⅰ~Ⅶ,Ⅰ区包含的物种有笔尖形根管藻粗径变种、北方角毛藻以及范氏星纹藻,对于北方角毛藻的生态类型,金德祥等[19]认为是广温性外洋种,而小久保清治[20]认为是寒带及北温带种;Ⅱ区包含的物种只有一个即瘤面角毛藻,为热带外洋性种;Ⅲ区包含的物种有佛氏梯形藻、地中海细柱藻、短刺角毛藻等;Ⅳ区包含的物种有翼鼻状藻、伏氏海线藻、离心列海链藻等;Ⅴ区包含的物种只有达蒂角毛藻,为热带外洋性种;Ⅵ区包含的物种有距端假管藻、圆柱几内亚藻、霍氏半管藻等,霍氏半管藻为暖海外洋性种,夏季在太平洋的中部及东部可以大量生长[21];Ⅶ区包含的物种较多,有密聚角毛藻、美丽漂流藻、大西洋角毛藻那不勒斯变种、丛毛辐杆藻、伯氏根管藻等。分别从这7个区中选出的优势种共12个作图(图5),分别为佛氏梯形藻、地中海细柱藻、密聚角毛藻、美丽漂流藻、大西洋角毛藻那不勒斯变种、距端假管藻、圆柱几内亚藻、达蒂角毛藻、伏氏海线藻、离心列海链藻、瘤面角毛藻以及笔尖根管藻粗径变种。
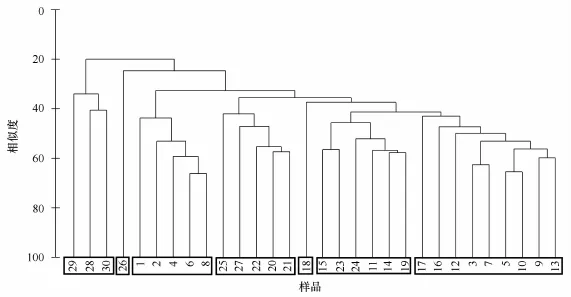
图4 聚类分析结果Fig.4 The result of cluster analysis

表1 调查区fi大于0.5的物种Tab.1 Species of fiexceeds 0.5 in survey area

续表1
3.3.2优势种平面分布
(1)佛氏梯形藻的平面分布如图5a。佛氏梯形藻的优势度明显高于其他物种,优势地位显著,为热带及亚热带外洋性种,印度洋、马来西亚群岛、地中海、印度尼西亚的爪哇都有出现且丰度较高[19]。其平均细胞丰度为0.204×103个/m3,在各站中的出现频率为0.956,仅在I406及HFA03站位没有出现。最大值出现在I312站位,最大细胞丰度为0.718×103个/m3,占该站总细胞丰度的25.3%。
(2)地中海细柱藻的平面分布如图5b。地中海细柱藻的生态类型属于暖海外洋性种,经常与一种原生生物共生,广泛分布于太平洋41°N—34°S之间[22]。地中海细柱藻平均细胞丰度为0.153×103个/m3,出现频率为0.766,最大值出现在I310站位,最大细胞丰度为0.718×103个/m3,占该站位总细胞丰度的34.9%。
(3)密聚角毛藻的平面分布如图5c。密聚角毛藻为热带至亚热带外洋性种,印度洋、菲律宾沿岸以及印度尼西亚的爪哇都有分布。其平均细胞丰度为0.077×103个/m3,出现频率为0.889,最大值出现在I312站位(0.384×103个/m3)。
(4)其余优势种的平面分布如图5d-l。美丽漂流藻(图5d)平均细胞丰度为0.067×103个/m3,大西洋角毛藻那不勒斯变种(图5e)平均细胞丰度为0.068×103个/m3,距端假管藻(图5f)平均细胞丰度为0.038×103个/m3,圆柱几内亚藻(图5g)平均细胞丰度为0.033×103个/m3,达蒂角毛藻(图5h)平均细胞丰度为0.037×103个/m3,伏氏海线藻(图5i)平均细胞丰度为0.037×103个/m3,离心列海链藻(图5j)平均细胞丰度为0.024×103个/m3,瘤面角毛藻(图5k)平均细胞丰度为0.019×103个/m3,笔尖根管藻粗径变种(图5l)平均细胞丰度为0.007×103个/m3。美丽漂流藻为热带外洋性种;大西洋角毛藻那不勒斯变种为暖海外洋性种,大西洋分布到50°N,美国加利福尼亚及地中海有记录;距端假管藻为暖水外洋性种,分布较广,印度洋、北大西洋以及黑海都有记录;圆柱几内亚藻为热带外洋性种,在大西洋、太平洋、印度尼西亚的爪哇都有记录;离心列海链藻为大洋性物种,但常见于近岸;笔尖根管藻粗径变种为暖水外洋性种,印度尼西亚的爪哇以及地中海都有记录。
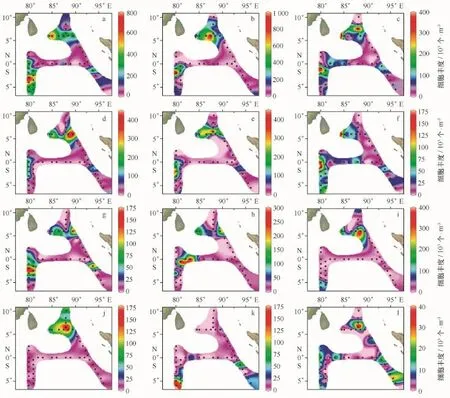
图5 优势种平面分布Fig.5 Horizontal distribution of the dominant species
4 讨论
本航次大面站营养盐实测数据缺乏,因此本文参考了World Ocean Atlas(WOA)13春季营养盐平均浓度值进行讨论。Sardessai等[24]的研究表明:东印度洋硝酸盐与磷酸盐的浓度与其他季节相比较低,与硝酸盐和磷酸盐相比,硅酸盐浓度较高。根据WOA 13数据春季调查海区200 m以浅平均营养盐分布如图6a-c,3种营养盐分布都不均匀。这3种营养盐的浓度在调查海区北部都较高,受赤道高温高盐水体的影响,赤道附近营养盐浓度较低。硝酸盐浓度从北至南呈降低趋势;磷酸盐浓度除个别站位外,其余浓度较高;而硅酸盐浓度除调查海区北部以外,浓度较低。硝酸盐平均浓度为13.41μmol/L,硝酸盐缺乏;磷酸盐的平均浓度为1.02μmol/L;硅酸盐平均浓度为11.12μmol/L,泡沫或大气输入[24]以及泡沫的破碎[25]会导致硅酸盐浓度升高。
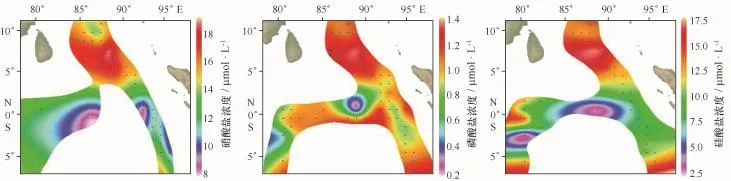
图6 2013年春季调查海域200 m以浅平均营养盐浓度平面分布Fig.6 Horizontal distribution of average nutrients concentration above 200 m water depth in the survey area in spring 2013
温度、盐度以及营养盐都是限制硅藻生长的因素,而营养盐浓度变化甚至会导致硅藻种群结构的变化[26]。本次调查期间赤道温度盐度较高,而WOA 13春季营养盐平均浓度数据也显示赤道的营养盐浓度较低,因此造就了赤道断面硅藻丰度的低值,而硅藻细胞丰度出现明显高值的两个站位,其营养盐的浓度值也较高。根据该海区3种营养盐的分布趋势以及硅藻的分布趋势可以看出,硅酸盐的浓度对硅藻的影响更为突出,这可能是因为相较于其他营养盐,硅酸盐对硅藻的影响更大。根据营养盐的分布趋势与12个优势种的分布趋势来看,除瘤面角毛藻外,其他优势种的分布与营养盐的分布相关性较大。
对于本次调查区域的浮游植物群落研究几乎没有,但是对相近海域的浮游植物群落的研究有一些,如Devassy和Goes[27]于1979年11月至1980年10月对印度西岸的浮游植物群落结构进行了调查,共鉴定出82个物种,其中包含62个硅藻;Choudhury和Pal[28]于2005年4月至2007年3月对孟加拉湾的印度东部沿岸进行了两年调查,共鉴定出浮游植物43种,细胞丰度也介于350~3 000个/mL;Li等[29]于2010年春季季风间期对孟加拉湾10°N断面进行了研究,硅藻最大丰度值达到4.0×104cells/L。此次研究对于填补整个印度洋的空白数据是十分有必要的。
海洋硅藻含有丰富的脂肪、多糖、蛋白质等物质,是海洋鱼类的重要饵料生物,它可以通过食物链直接或间接被海洋虾类、鱼类等经济动物摄食。本次调查的优势种角毛藻较多,而角毛藻是虾类[30]等的重要饵料,因此在一定程度上,本文所研究浮游硅藻结构可为印度洋远洋渔业资源生物分布提供参考,例如金枪鱼等。
5 结论
本次调查共鉴定出硅藻34属113种(包括6个变种,3个变型及5个未定种),其中角毛藻属物种数最多;优势种为佛氏梯形藻、地中海细柱藻、密聚角毛藻美丽漂流藻、大西洋角毛藻那不勒斯变种、距端假管藻、圆柱几内亚藻等,生态类型为热带外洋性或暖海外洋性物种;调查海区硅藻平均细胞丰度为1.855 ×103个/m3,北部海区为分布密集区,赤道断面细胞丰度整体较低。
东印度洋赤道区浮游植物物种组成和群落结构的研究成果还颇少,本文只是初步的研究成果,今后还需加强印度洋浮游植物群落的长周期调查以及其与全球变化和渔业之间关系的深入研究。
致谢:本次拖网样品的采集得到翟惟东、吴礼彬、苏宏飞、宋勇军的帮助以及“实验1号”全体船员的辛勤工作,特此致谢。
[1]孙军,刘东艳,钱树本.琉球群岛及其邻近海域的浮游硅藻——1997年夏季的种类组成和丰度分布[G]//中国海洋学文集第12集.北京:海洋出版社,2000:158-169.Sun Jun,Liu Dongyan,Qian Shuben.Planktonic diatoms in Ryūkyū-guntōand its adjacent waters-species composition and their abundance distribution in the summer of 1997[G]//China Oceanography CorpusⅡ.Beijing:China Ocean Press,2000,12:158-169.
[2]钱树本,刘东艳,孙军.海藻学[M].青岛:中国海洋大学出版社,2005:1-5.Qian Shuben,Liu Dongyan,Sun Jun.Hai Zao Xue[M].Qingdao:Ocean University of China Publishing House,2005:1-5.
[3]朱根海,宁修仁,蔡昱明,等.南海浮游植物种类组成和丰度分布的研究[J].海洋学报,2003,25(2):8-23.Zhu Genhai,Ning Xiuren,Cai Yuming,et al.Studies on species composition and abundance distribution of phytoplankton in the South China Sea[J].Haiyang Xuebao,2003,25(2):8-23.
[4]Round F E.The Ecology of Algae[M].New York:Cambridge University Press,1981:461-483.
[5]徐建军,王东晓.印度洋-太平洋海温的年际、年代际异常及其对亚洲季风的影响[J].海洋学报,2000,22(3):34-43.Xu Jianjun,Wang Dongxiao.Diagnosis of interannual and interdecadal variation in SST over Indian-Pacific Ocean and numerical simulation of their effect on Asian summer monsoon[J].Haiyang Xuebao,2000,22(3):34-43.
[6]Wiggert J D,Hood R R,Naqvi S W A,et al.Indian Ocean Biogeochemical Processes and Ecological Variability[M].American Geophysical Union,Washington,DC,2009:409-428.
[7]Jagadeesan L,Jyothibabu R,Anjusha A,et al.Ocean currents structuring the mesozooplankton in the Gulf of Mannar and the Palk Bay,southeast coast of India[J].Prog Oceanogr,2013,110:27-48.
[8]宣莉莉,邱云,许金电,等.热带东印度洋表层环流季节变化特征研究[J].热带海洋学报,2014,33(1):26-35.Xuan Lili,Qiu Yun,Xu Jindian,et al.Seasonal variation of surface-layer circulation in the eastern tropical Indian Ocean[J].Journal of Tropical Oceanography,2014,33(1):26-35.
[9]Madhupratap M,Gauns M,Ramaiah N,et al.Biogeochemistry of the Bay of Bengal:physical,chemical and primary productivity characteristics of the central and western Bay of Bengal during summer monsoon 2001[J].Deep-Sea Research PartⅡ:Topical Studiesin Oceanography,2003,50(5):881-896.
[10]Charlson R J,Lovelock J E,Andreae M O,et al.Oceanic phytoplankton,atmospheric sulphur,cloud albedo and climate[J].Nature,1987,326(6114):655-661.
[11]Sathyendranath S,Gouveia A D,Shetya S R,et al.Biological control of surface temperature in the Arabian Sea[J].Nature,1991,349(6304):54 -56.
[12]杨秋明.夏季江淮地区雨量与印度洋海温联系的年代际变化[J].热带海洋学报,2005,24(5):31-42.Yang Qiuming.Interdecadal variations of relationships between summer rainfall in Changjiang and Huaihe Valleys and SST in Indian Ocean[J].Journal of Tropical Oceanography,2005,24(5):31-42.
[13]Tarran G A,Burkill P H,Edwards E S,et al.Phytoplankton community structure in the Arabian Sea during and after the SW monsoon,1994[J].Deep-Sea Research PartⅡ:Topical Studies in Oceanography,1999,46(3/4):655-676.
[14]Schlüter L,Henriksen P,Nielsen T G,et al.Phytoplankton composition and biomass across the southern Indian Ocean[J].Deep-Sea Research PartⅠ:Oceanographic Research Papers,2011,58(5):546-556.
[15]国家技术监督局.GB/T 12763.6-1991,海洋调查规范:海洋生物调查[S].北京:中国标准出版社,1992:17-20.State Bureau of Technical Supervision.GB/T 12763.6-1991,The specification for oceanographic survey:marine biological survey[S].Beijing:China Standards Press,1992:17-20.
[16]Tomas C R.Identifying Marine Phytoplankton[M].San Diego:Academic Press,1997:1-858.
[17]孙军,刘东艳.中国海区常见浮游植物种名更改初步意见[J].海洋与湖沼,2002,33(3):271-286.Sun Jun,Liu Dongyan.The preliminary notion on nomenclature of common phytoplankton in China seas waters[J].Oceanologia et Limnologica Sinica,2002,33(3):271-286.
[18]孙儒泳.动物生态学原理[M].2版.北京:北京师范大学出版社,1992:356-357.Sun Ruyong.Dongwu Shengtaixue Yuanli[M].2nd.Beijing:Beijing Normal University Press,1992:356-357.
[19]金德祥,陈金环,黄凯歌.中国海洋浮游硅藻类[M].上海:上海科学技术出版社,1965:1-230.Jin Dexiang,Chen Jinhuan,Huang Kaige.Zhongguo Haiyang Fuyouguizaolei[M].Shanghai:Shanghai Science Press,1965:1-230.
[20]小久保清治.浮游矽藻类[M].华汝成,译.上海:上海科学技术出版社,1960.Seiji Kokubo.Fuyou Xizaolei[M].Hua Rucheng,trans.Shanghai:Shanghai Scientific and Technical Publishers,1960.
[21]Villareal T A,Adornato L,Wilson C,et al.Summer blooms of diatom-diazotroph assemblages and surface chlorophyll in the North Pacific gyre:a disconnect[J].Journal of Geophysical Research,2011,116(C3).
[22]Gómez F.The Consortium of the protozoan Solenicolasetigera and the diatom Leptocylindrusmediterraneus in the Pacific Ocean[J].Acta Protozoologica,2007,46(1):15-24.
[23]彭欣,宁修仁,孙军,等.南海北部浮游植物生长对营养盐的响应[J].生态学报,2006,26(12):3959-3968.Peng Xin,Ning Xiuren,Sun Jun,et al.Responses of phytoplankton growth on nutrient enrichments in the northern South China Sea[J].Acta Ecologica Sinica,2006,26(12):3959-3968.
[24]Sardessai S,Shetye S,Maya M V,et al.Nutrient characteristics of the water masses and their seasonal variability in the eastern equatorial Indian Ocean[J].Marine Environmental Research,2010,70(3/4):272-282.
[25]Harvey H W.The Chemistry and Fertility of Sea Waters[M].Cambridge:Cambridge University Press,1955:240.
[26]Baylor E R,Sutcliffe W H,Hirschfeld D S.Adsorption of phosphate onto bubbles[J].Deep-Sea Research,1962,9:120-124.
[27]Devassy V P,Goes J I.Phytoplankton community structure and succession in a tropical estuarine complex(central west coast of India)[J].Estuarine,Coastal and Shelf Science,1988,27(6):671-685.
[28]Choudhury A K,Pal R.Phytoplankton and nutrient dynamics of shallow coastal stations at Bay of Bengal,Eastern Indian coast[J].Aquatic Ecology,2010,44(1):55-71.
[29]Li Gang,Ke Zhixin,Lin Qiang,et al.Longitudinal patterns of spring-intermonsoon phytoplankton biomass,species compositions and size structure in the Bay of Bengal[J].Acta Oceanologica Sinica,2012,31(2):121-128.
[30]曹淑莉,向葆卿.8种海洋饵料微藻蛋白质含量及氨基酸组成比例的比较研究[J].海洋学报,1993,15(4):98-103.Cao Shuli,Xiang Baoqing.Comparative study of 8 marine bait microalgae protein content and ammonia acid composition proportion[J].Haiyang Xuebao,1993,15(4):98-103.
Diatom communities in equatorial region and its adjacent areas of eastern Indian Ocean during spring intermonsoon 2014
Xue Bing1,2,Sun Jun1,2,Ding Changling1,2,Wang Dongxiao3
(1.College of Marine and Environmental Sciences,Tianjin University of Science and Technology,Tianjin 300457,China;2.Tianjin Key Laboratory of Marine Resources and Chemistry,Tianjin University of Science and Technology,Tianjin 300457,China;3.State Key Laboratory of Tropical Oceanography,South China Sea Institute of Oceanology,Chinese Academy of Sciences,Guangzhou 510301,China)
Phytoplankton net samples were collected from 45 grid stations(10.08°N—6.00°S,80.00°—96.10°E)from April 10 to May 13,2014 in eastern Indian Ocean,diatoms including 34 genera and 113 species(including variety,form and unidentified taxa)were found in the survey area.Over half species were tropical oceanic species and warm oceanic species,which was in line with the tropical and subtropical environmental characteristics in the survey area.The dominant species included Climacodium frauenfeldianum,Leptocylindrus mediterraneus,Chaetoceros coarctatus,Planktoniella foromsa,Chaetoceros atlanticus var.neapolitanus,Pseudosolenia calcar-avis,Guinardia cylindrus,Chaetoceros dadayi,Thalassionema frauenfeldii,Thalassiosira excentrica,Chaetocerosbacteriastroides,Rhizosolenia styliformis var.latissima.The average cell abundance of diatoms was 1.855×103cells/m3.The horizontal distribution of diatoms was asymmetrical,the cell abundance was low in the equatorial section,and the northern area was the high abundance zone.Seven groups were divided through the cluster analysis,the species composition and horizontal distribution of diatoms had a good correlation with the integrated average temperature,salinity and nutrients above 200 m.
diatoms;eastern Indian Ocean;species composition;horizontal distribution;cluster analysis
薛冰,孙军,丁昌玲,等.2014年春季季风间期东印度洋赤道及其邻近海域硅藻群落[J].海洋学报,2016,38(2):112—120,
10.3969/j.issn.0253-4193.2016.02.011
Xue Bing,Sun Jun,Ding Changling,et al.Diatom communities in equatorial region and its adjacent areas of eastern Indian Ocean during spring intermonsoon 2014[J].Haiyang Xuebao,2016,38(2):112—120,
10.3969/j.issn.0253-4193.2016.02.011
Q948.8
A
0253-4193(2016)02-0112-09
2015-04-20;
2015-09-27。
国家自然科学基金(41276124,41176136,41406155);教育部新世纪优秀人才计划(NCET-12-1065);天津市高校创新团队计划(TD12-5003)。
薛冰(1991—),女,天津市人,主要从事海洋浮游植物群落的研究。E-mail:xuebingjy@163.com
孙军,男,教授,主要从事海洋生态学的研究。E-mail:phytoplankton@163.com
猜你喜欢
大自然探索(2023年7期)2023-11-14 13:07:36
课堂内外(小学版)(2023年9期)2023-10-11 14:39:26
海洋信息技术与应用(2021年3期)2022-01-17 02:37:26
黑龙江水利科技(2020年8期)2020-01-12 06:26:00
建筑热能通风空调(2018年5期)2018-07-09 03:16:44
上海建材(2018年1期)2018-04-18 12:15:16
水利技术监督(2017年6期)2017-12-19 13:28:17
海洋渔业(2016年6期)2016-04-16 03:00:23
中国工程咨询(2016年9期)2016-02-13 03:12:40
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
