
紫芝担孢子交配型的鉴定*
2016-11-30 12:53:52柯斌榕卢政辉吴小平郭丽羡兰清秀
中国食用菌 2016年4期
柯斌榕,卢政辉,吴小平,郭丽羡,兰清秀
(1.福建省农业科学院食用菌研究所,特色食用菌繁育与栽培国家地方联合工程研究中心,福建福州3500014;2.福建农林大学菌物研究中心,福建福州350002)
紫芝担孢子交配型的鉴定*
柯斌榕1,卢政辉1,吴小平2**,郭丽羡2,兰清秀1
(1.福建省农业科学院食用菌研究所,特色食用菌繁育与栽培国家地方联合工程研究中心,福建福州3500014;2.福建农林大学菌物研究中心,福建福州350002)
以紫芝(Ganoderma sinense)的担孢子萌发获得的单核菌株为试验材料,采用三轮杂交法、OWE-SOJ技术及核迁移技术对其担孢子单核体交配型因子进行准确测定。结果表明,紫芝为四级性异宗结合菌类,其原生质体单核化菌株交配型分离比例符合1:1的预期,不同交配型担孢子的单核体符合1:1:1:1的自由分离规律,采用OWE-SOJ与核迁移试验相互印证,可以对紫芝担孢子交配型进行准确测定。
紫芝;交配型;四级性;核迁移;OWE-SOJ
紫芝(Ganoderma sinense J.D.Zhao,L.W.Hsu& X.Q.Zhang)又名黑芝、中国灵芝,是中国特有的一个灵芝种类,并由我国灵芝分类专家赵继鼎先生首次命名[1-2]。作为传统的药用灵芝品种,与赤芝和松杉灵芝一起收录入《中国药典》中并得到广泛使用[3]。紫芝味甘、性温,较赤芝而言苦味较轻,含有多种活性成分,如多糖、灵芝酸、生物碱等,具有安神、消炎、利尿等功效。
目前紫芝研究主要侧重于功效成分和药理研究,而在遗传育种研究方面则鲜有报道。栽培的紫芝菌种也主要以野生分离驯化为主,目前审定的唯一一个紫芝品种“武芝2号”也是通过野生驯化而得到的[4]。刘新锐等[5]通过原生质体单核化技术测定了紫芝的不亲和性因子,表明紫芝具有丰富的不亲和性因子,但未就担孢子单核体的交配因子进行分析。由于担孢子是减数分裂的产物,含有丰富的遗传变异因子。对紫芝担孢子单核体交配型进行研究,不仅有助于了解紫芝的生活史,也为新品种改良和杂交育种提供实验材料。本文通过对紫芝担孢子单核体进行紫芝交配型因子的研究,拟为将来利用担孢子单核体的遗传多样性进行杂交育种打下基础。
1 材料与方法
1.1供试菌株
供试菌株紫芝由福建农林大学菌物研究中心提供。
1.2培养基
PDA:土豆200 g,1 000 g水煮沸后过滤,2%琼脂条煮沸后加入葡萄糖20 g,搅拌均匀,然后制备试管斜面。
OWE培养基:5%杂木屑浸汁200 mL,琼脂20 g,加蒸馏水补至1 000 mL。5%杂木屑浸汁制备,取50 g杂木屑于1 L水中煮60 min,过滤去杂质加蒸馏水补足1 L。
SOJ培养基:鲜榨橘汁200 mL,121℃高压灭菌30 min,使用时取20 mL,加入180 mL融化的水琼脂中倒平板备用。
1.3担孢子收集及单核体分离
选取新鲜刚开始弹射的紫芝子实体,采取活体套袋收集担孢子的方法进行收集。将当天收集的担孢子,常规稀释至100倍,在显微镜视野中有10个~20个孢子即可,并涂布到PDA培养基中,25℃条件下培养,随时观察担孢子萌发情况[6]。当有担孢子萌发,即可按谢宝贵[7]的方法挑取,并镜检获取孢子单核体。
1.4标准测交菌株制备
标准菌株采用原生质体单核化制备,方法参考吴小平等[8]制备赤芝原生质体的方法,对再生的菌株进行镜检,选取无锁状联合的单核体作为标准测交菌株[8]。随机选取一个单核菌株为T1,交配型定为AxBx;将能与其亲和配对的单核菌株定为T2,交配型定为AyBy,并将其作为标准测交菌株。
1.5交配型分析
采用三轮杂交法进行常规交配型分析[9],以标准菌株T1和T2作为测试菌株,将孢子单核体作为待测菌株进行配对,以锁状联合为标记进行亲和与否的判定,以此进行单核体的交配型分析,与标准测交菌株均不亲和的任选一个为测交菌株T3,将与T3亲和的菌株定为T4。
1.6交配型χ2测验
通过卡方测验对原生质体单核化菌株及4种担孢子萌发的单核体进行检测,验证不同交配型的单核体的比例是否符合预期原生质体单核化单核体1∶1及孢子单核体1∶1∶1∶1的比例[10]。
1.7单核菌株交配型准确测定
1.7.1OWE-SOJ反应
参考Darmono TW的方法将4种交配型T1、T2、T3、T4两两配对接种于OWE培养基上,两接种块间相距约0.5 cm。25℃对峙培养7 d~10 d。当接种块菌丝接触后,将两接种块沿配对区垂直整条切出宽4 mm,长1.5 cm的琼脂条,转接入SOJ培养基中。25℃培养10 d~15 d观察菌落形态。如果出现无锁状联合的绒毛状菌落,为A=B≠的反应;出现无锁状联合的栅栏型菌落,则为A≠B=的反应[11]。
1.7.2核迁移测定
四级性的异宗结合交配系统,是由A和B两个交配型因子控制。当2个单核菌株A因子相同B因子不同时,会出现核迁移现象,但不形成锁状联合结构。具体操作步骤参考陈莉[12]的方法,取单核菌株T1分别与T3及T4菌株进行第一次配对。待菌丝交接后,取T1外侧的菌块与T4配对,取T3外侧的菌块与T2配对。观察第二次配对是否可亲和,如果可亲和,则T1与T3间存在核迁移,其B因子不相同。反之则T1与T3间无核迁移,由此可以推测T3和T4的交配型。
2 结果与分析
2.1紫芝子实体及担孢子形态
紫芝子实体及担孢子形态见图1。
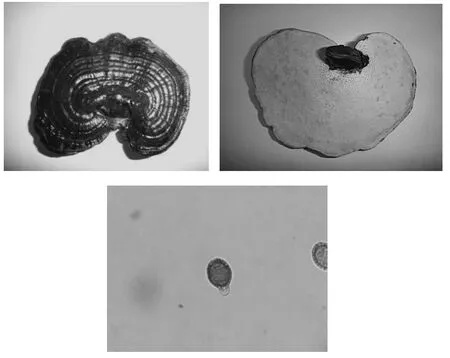
图1 紫芝子实体及担孢子形态Fig.1Fruit body and basidiospores morphologies of Ganoderma sinense
由图1可见,紫芝子实体呈半圆形、肾形或如意形,人工段木栽培条件下也常呈现近圆形;子实体皮壳成紫黑色,具有漆样光泽;菌盖表面有同心环纹。菌管层表面呈浅褐色,成熟后则呈褐色。菌柄侧生,同菌盖类似具有漆样光泽。紫芝担孢子呈卵形,相较赤芝担孢子要大。顶端呈脐突形,或稍平截,双层壁,外壁透明,内壁平滑,内壁淡褐色有明显小刺,成熟担孢子大小为(10~12)μm×(6.5~8.0)μm。
2.2标准测交菌株及担孢子单核体分离
单核体菌株分离见图2。
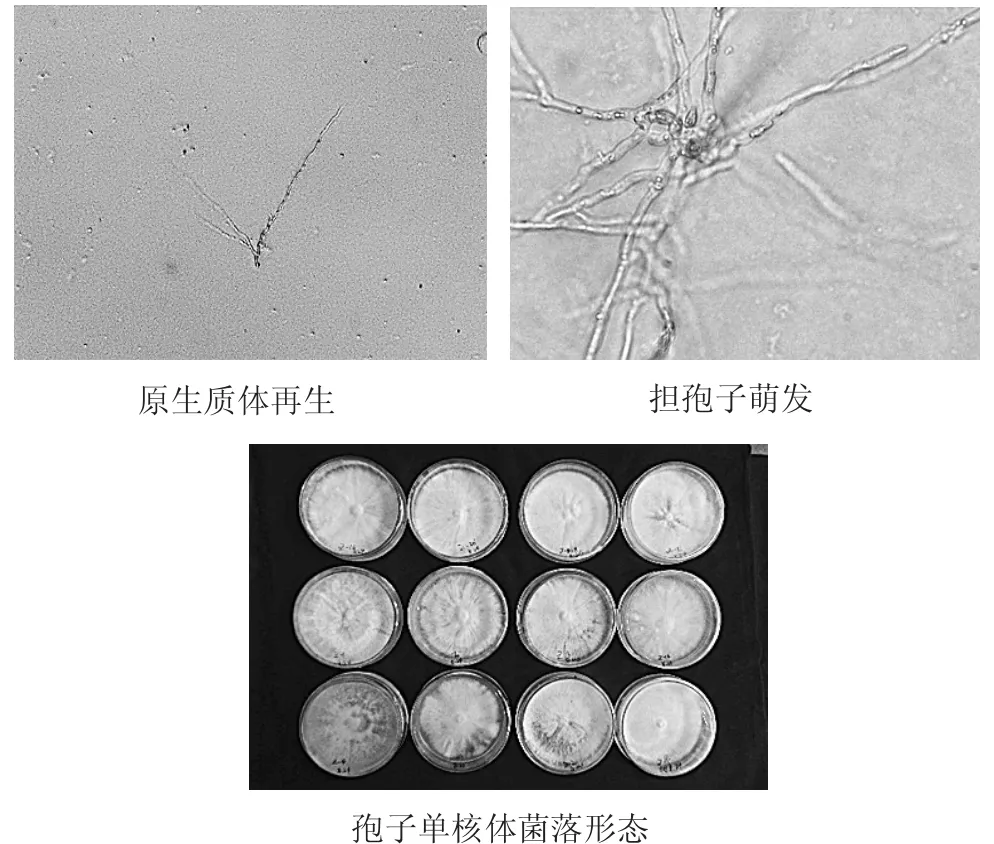
图2 单核体菌株分离Fig.2Obtained monokaryogenesis
通过原生质体再生,共挑取了50个再生菌株,经镜检筛选获得了29个单核菌株,单核化率为58.00%。试验过程发现紫芝的原生质体单核化率与制作原生质体所采用的菌丝培养时间关系密切,幼嫩的菌丝体更易再生且获得单核菌株。利用担孢子萌发的方式,共获得紫芝孢子单核体31个。因为担孢子是减数分裂的产物,存在广泛的变异,具体表现在不同单核菌株间生长速度和菌落形态存在明显差异。
2.3交配型结果分析
通过亲和配对,将原生质体单核化获得的单核菌株分为T1和T2两类,并以其作为标准菌株进行孢子单核体的亲和配对。通过常规三轮杂交配对,结果见表1。
由表1可见,通过χ2验证表明,原生质体单核化2种交配型的分离比例符合预期,孢子单核体的分离比例也符合1∶1∶1∶1的自由分离定律。
2.4交配型准确测定结果
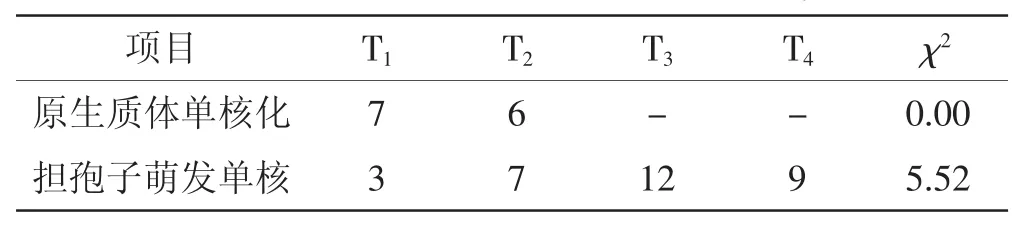
表1 紫芝原生质体单核化及单核菌株的交配型分析Tab.1Analysis on mating type of monokaryons
通过OWE-SOJ技术进行不同孢子单核体间的配对,见图3。从图3可以观察到3种不同的结果,无锁状联合的栅栏菌落、有锁状联合的绒毛状菌落和无锁状联合的绒毛状菌落。通过不同交配型因子的单核体间配对反应类型的不同,可以推测出T3和T4的交配型应为AXBY和AYBX。

图3 不同孢子单核体配对的OWE-SOJ反应类型Fig.3Interaction types of confrontation culture of different monospores using OWE-SOJ technique
利用核迁移技术进一步验证OWE-SOJ反应的结果,再次验证T3和T4的交配型,结果见图4。
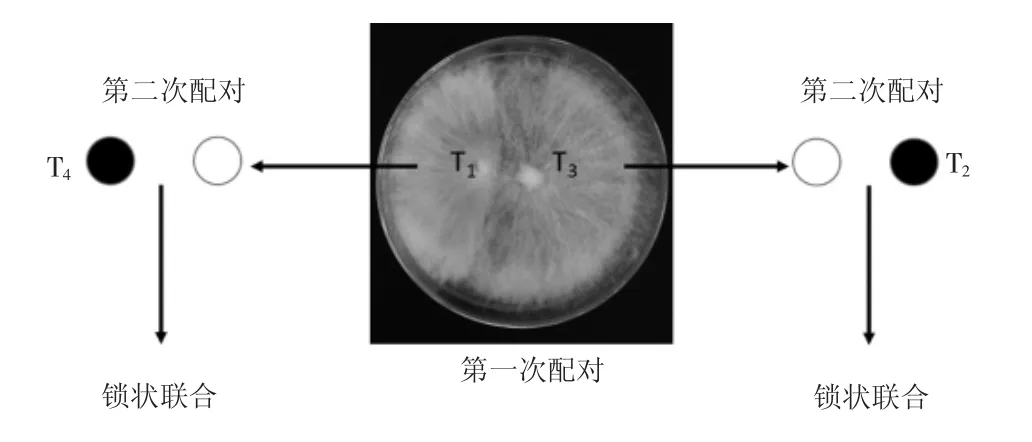
图4 紫芝单核菌丝的核迁移Fig.4Nuclear migration of Ganoderma sinense
由图4可见,将T1与T3进行第一次配对后,再与不亲和的T4和T2进行配对,通过镜检第二次配对发现锁状联合,所以可以推断T1和T3、T2和T4之间存在核迁移现象,T1和T4、T2和T3之间不存在核迁移。因此可以推定紫芝4种孢子单核体的准确交配型为:T1(AXBX)、T2(AyBy)、T3(AXBY)和T4(AYBX)。
3 讨论
紫芝为四级性异宗结合菌类,其交配型受两对独立的交配型因子控制。其中交配型因子A控制菌丝融合和锁状联合的形成,B因子则控制细胞核迁移。担孢子萌发获得的单核体根据交配型不同可以分为四类,通过与标准测交菌株配对可以获得亲本型和非亲本型的单核体。本研究获得的孢子单核体符合预期的自由分离比例,但非亲本型的单核体多于亲本型,这可能与担孢子的萌发率或挑取的单核体数量不足有关。陈裕新等[13]发现可以使用OWESOJ技术研究赤芝交配型因子。本研究采用OWESOJ技术与和迁移技术相结合,既可以克服OWESOJ反应中反应形态不够明显,也可以解决核迁移试验中锁状联合不易观察的问题[13]。二者相互验证可以准确测定T3和T4的交配型,为下一步开展品种选育与改良试验奠定基础。
本研究并没有发现重组型的交配型因子出现,这可能与交配型因子中各亚基的距离较近,重组率较低有关。随着全基因组测序工作的开展,赤芝的交配型因子结构已经绘制,这将有助于指导交配型因子的深入研究[14]。同时研究过程中发现,不同的孢子单核体在菌落形态、生长速度等方面具有多态性。孢子单核体遗传多样性的深入研究,将有助于挖掘品种特性,并应用于紫芝品种改良和遗传研究。
[1]林志彬.灵芝的现代研究(第三版)[M].北京:北京大学医学出版社,2007:3-8.
[2]戴玉成,杨祝良.中国药用真菌名录及部分名称的修订[J].菌物学报,2008,27(6):801-824.
[3]中华人民共和国卫生部药典委员会.中华人民共和国药典[M].北京:中国医药科技出版社,2010:174.
[4]钟礼义.紫芝新品种“武芝2号”[J].农村百事通,2013(16):39.
[5]刘新锐,王圣铕,谢宝贵,等.紫芝不亲和性因子分析[J].菌物学报,2014,33(2):464-468.
[6]柯斌榕,吴小平.赤芝与无柄灵芝担孢子的萌发特性比较[J].食药用菌,2015,23(5):282-287.
[7]谢宝贵.倒置显微镜单孢分离法[J].中国食用菌,2008,24(6):56.
[8]吴小平,刘方,谢玉荣,等.灵芝原生质体单核化杂交育种[J].中国农学通报,2011,25(23):64-69.
[9]季哲,李玉祥,薛淑玉.黄伞的交配型性状研究[J].菌物学报,2004,23(1):38-42.
[10]董洪新,蔡德华,李玉.猪肚菇担孢子交配型的分析[J].微生物学报,2010,37(11):1617-1620.
[11]Darmono TW.Morphological characteristics of incompatibility reactions and evidence for nuclear migration in Armillaria mellea[J].Mycologia,1992,84(3):367-375.
[12]陈莉,李安政,林范学,等.糙皮侧耳担孢子交配型的鉴定[J].微生物学通报,2007,34(6):1086-1089.
[13]陈裕新,夏志兰,刘鹏,等.灵芝群体交配基因型分析[J].中国农学通,2012,28(10):213-218.
[14]吴弘毅.灵芝交配型与出菇相关基因之选殖与分析[D].台北市:国立台湾大学,2005:80-85.
Accurate Determination of Mating Type of Ganoderma sinense Basidispores
KE Bin-rong1,LU Zheng-hui1,WU Xiao-ping2,GUO Li-xian2,LAN Qing-xiu1
(1.Edible Fungi Institute of Fujian Academy of Agricultural Sciences,National and Local Joint Engineering Research Center for Breeding&Cultivation of Featured Edible Fungi,Fujian Academy of Agricultural Sciences,Fuzhou 350014,China; 2.Mycological Research Center of Fujian Agriculture and Forestry University,Fuzhou 350002,China)
Four mating types of basidispores from Ganderma sinense were determined using three-cycle mating system,OWESOJ technique and nuclear migration test.Test results showed that G.sinense was a kind of basidiomycete with tetrapolarity heterothallism.The different mating types of protoplasted monokaryon and basidispores were present in a 1:1 and 1:1:1:1 ratio. The nuclear migration test is consist with that of OWE-SOJ,and they could establish corroboration for each other.Therefore it was feasible to apply this method to determine the mating type of basidispores accurately.
Ganoderma sinense;mating type;tetrapolarity;nuclear migration;OWE-SOJ
S646.9
A
1003-8310(2016)04-0024-04
10.13629/j.cnki.53-1054.2016.04.006
福建省种业创新与产业化工程专项项目(2014S1477-21);福建省食用菌产业重大农技推广服务试点项目(KNJ153013)。
柯斌榕(1986-),男,硕士,研究实习员,主要从事食(药)用菌栽培及育种。E-mail:kebinrong@qq.com
**通信作者:吴小平(1965-),男,博士,教授,主要从事食用菌教学与科研工作。E-mail:fjwxp@126.com
2016-05-28
猜你喜欢
广东园林(2021年3期)2021-07-12 00:57:58
福建农业学报(2020年7期)2020-10-31 06:53:44
天然产物研究与开发(2018年10期)2018-11-06 07:43:32
食用菌(2017年5期)2017-10-19 03:02:28
天然产物研究与开发(2016年6期)2016-06-05 10:29:26
首都医科大学学报(2015年4期)2015-12-16 13:00:08
大连工业大学学报(2015年4期)2015-12-11 04:06:50
特产研究(2015年1期)2015-04-12 06:36:11
食品工业科技(2014年23期)2014-03-11 18:19:06
无机化学学报(2014年12期)2014-02-28 17:33:53
