
云南野生莲瓣兰形态分异与海拔的关系
2016-11-28 09:03:49毕玉芬王鸿泽
生态学报 2016年19期
索 默, 毕玉芬,*, 王鸿泽, 唐 敏, 叶 广
1 云南农业大学 动物科学技术学院, 昆明 650201 2 云南农业大学 园林园艺学院, 昆明 650201
云南野生莲瓣兰形态分异与海拔的关系
索 默1, 毕玉芬1,*, 王鸿泽1, 唐 敏2, 叶 广2
1 云南农业大学 动物科学技术学院, 昆明 650201 2 云南农业大学 园林园艺学院, 昆明 650201
莲瓣兰是中国西南地区特有的兰科濒危物种。2013年和2014年8月至10月, 在云南省西北地区选取了28个野生莲瓣兰居群进行野外调查。结果表明:野生莲瓣兰在滇西北海拔为1380—2557 m的区域均有分布;莲瓣兰叶片数、花葶长、花瓣长、花瓣宽、萼片长均与海拔呈显著负相关(P<0.01)。与年平均气温呈正相关的形态指标有:叶长(P<0.05), 株高(P<0.05), 花葶长(P<0.01)。与空气湿度呈显著正相关的形态指标有:叶宽(P<0.05), 株高(P<0.05), 花葶长(P<0.05), 叶长(P<0.01)。随着野生莲瓣兰分布地海拔的升高, 株型矮化, 形态变小;年均温越高、空气湿度越大的分布地, 莲瓣兰叶片越长、越宽, 植株越高, 花葶越长;分布地辐射强度越大, 莲瓣兰叶片越短, 植株越矮, 根系越发达。野生莲瓣兰居群间的叶长、叶宽、株高、根长及花葶长度的变化较大, 变异系数超过了15%, 表明其存在着较大的形态变异, 生态适应幅度变宽, 且植株矮化严重, 需加强就地和迁地保护。研究结果阐明了滇西北海拔、温度等生态因子对野生莲瓣兰形态特征的影响, 为进一步保护野生莲瓣兰资源提供了理论依据。
野生莲瓣兰;海拔;年平均温度;空气湿度;辐射强度
野生莲瓣兰(Cymbidiumtortisepalum)主要分布于云南大理、怒江、保山、迪庆和丽江等州市[1]。该区域属青藏高原南延部分, 为横断山脉纵谷区, 地形波状起伏, 平均海拔2000 m左右, 高山深谷相间, 相对高差较大, 地势险峻, 气候变化显著, 是野生莲瓣兰的主要分布地。近年来随着莲瓣兰观赏价值的日益提升, 对野生资源的采挖及商业交易日趋频繁, 导致其野生居群数量急剧减少, 生存环境遭到严重破坏。莲瓣兰已被列入《野生动植物濒危物种国际贸易公约》[2], 急需得到保护。目前对于莲瓣兰的研究主要集中在资源分子鉴定和分类学[3-6]、组织培养[7-8]、形态变异[9-10]、鉴赏栽培[11-12]方面, 有关资源保育研究仍处于初级阶段[13-14]。
有关莲瓣兰野生居群的研究较少。叶广等[15]对云南迪庆州野生莲瓣兰居群形态特征与分布地海拔、坡向的关系进行了研究, 指出野生莲瓣兰的株高、叶长、叶宽与分布地海拔高度呈显著负相关;西北坡向、西坡向和北部坡向是野生莲瓣兰的主要分布区。但研究涉及的生态因子较少, 且范围仅限于迪庆州。本研究通过对滇西北(怒江、保山、大理、迪庆、丽江)28个野生莲瓣兰居群分布地的生态因子和莲瓣兰形态指标的测定和分析, 解析生态因子对莲瓣兰形态特征的影响, 了解维持该物种繁衍的生态环境, 为云南野生莲瓣兰资源的保育提供理论依据。
1 材料和方法
1.1 资源采集地的确定
采集地集中在云南西北部, 以保山市施甸县、大理州云龙县、丽江市玉龙县、怒江州兰坪县、迪庆州维西县为主要采集地。样地之间的直线距离均大于2 km。
1.2 生态因子和野生居群特征测定方法
2013年和2014年进行实地考察。8月至10月进行野外生境考察和居群调查。共考察到28个莲瓣兰野生居群分布地点(表1)。居群样方面积为100 m2。样地对角线上随机取5个测定点测定生态因子数值并取平均值。利用GPS对海拔及经纬度进行记录;利用空气温湿度仪(DT-321S)和土壤温湿度测定记录仪(HA1055)分别测定居群空气和土壤温湿度;利用太阳辐射测试仪(FM-TF1)测定太阳辐射强度;利用风速仪测定风速。在居群内随机选取10株莲瓣兰, 用游标卡尺和直尺测量每株莲瓣兰的株高(自然高度)、叶长、叶宽、根长、根直径, 记录叶片数及侧根数, 每个居群单独计算平均值;在开花期测定每株莲瓣兰所有花朵的花瓣长、花瓣宽、萼片长、萼片宽和花葶长(自基生莲座至花底部的长度), 每个居群单独计算平均值;若居群不足10株则全部测量。用SPSS 19及Microsoft Excel 2007进行分析。
1.3 野生莲瓣兰形态变异系数的计算方法
变异系数(C.V)的计算公式:

式中,SD为标准偏差,MN为平均值。
2 结果与分析
2.1 分布地海拔高度对野生莲瓣兰形态特征的影响
对28个野生莲瓣兰居群分布地生态因子和形态指标进行测定, 结果见表1和表2。由表1和表2可知, 云南野生莲瓣兰居群均分布于海拔较高的山区, 且分布地海拔梯度较大。野生居群分布的最低海拔在大理州云龙县旧州镇, 海拔为1380 m;最高海拔分布在丽江市玉龙县黄山镇, 海拔为2557 m。两样地的海拔高度相差1270 m;两样地莲瓣兰居群的叶长、叶宽、株高分别相差20.99、0.22、15.7 cm, 且差异显著(P<0.05)。由图1和图2可知, 其叶长和株高与海拔呈显著负相关(P<0.01), 叶宽与海拔也呈负相关(P<0.05)。表明野生莲瓣兰对海拔的适应幅度比较广, 1380—2557 m是云南滇西北野生莲瓣兰分布的海拔范围。上述结果表明, 海拔是影响莲瓣兰株高、叶长和叶宽的重要因子, 其中叶片长度对海拔变化最为敏感。
从图1—图3可见, 野生莲瓣兰叶片数、花葶长、花瓣长、花瓣宽、萼片长均与海拔呈负相关(P<0.01)。根长与海拔呈正相关, 但差异不显著。表明云南28个野生莲瓣兰居群随着分布地海拔的升高, 叶片逐渐变得细短稀疏, 植株矮化, 花瓣、萼片、花葶均变短变小, 但对根系生长无显著影响。
2.2 分布地年平均气温对野生莲瓣兰形态特征的影响
野生莲瓣兰居群分布地年平均气温与莲瓣兰叶长、叶宽等形态指标相关性分析结果见图4。莲瓣兰叶长和株高与分布地年平均气温均呈正相关(P<0.05);莲瓣兰花葶长与年平均气温呈正相关(P<0.01)。叶宽与年平均气温呈正相关但相关不显著。根长与年平均温度负相关, 但相关不显著。综合表1和图4结果, 28个野生居群分布地平均气温范围为10—21℃;表明野生莲瓣兰分布地温度越高, 莲瓣兰的叶片越长, 植株越高、花葶越长, 植株越高大。
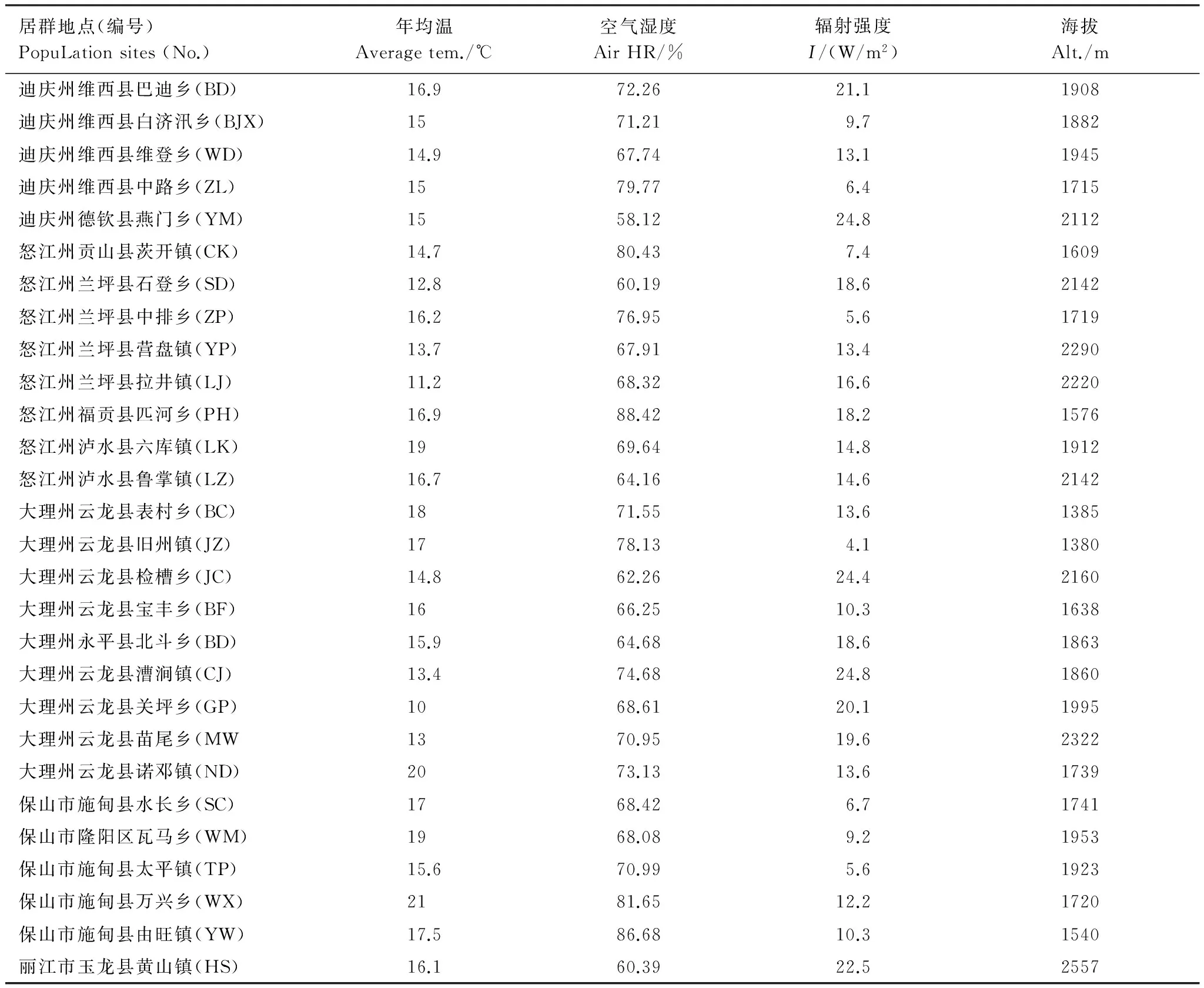
表1 野生莲瓣兰28个居群采样点主要生境因子
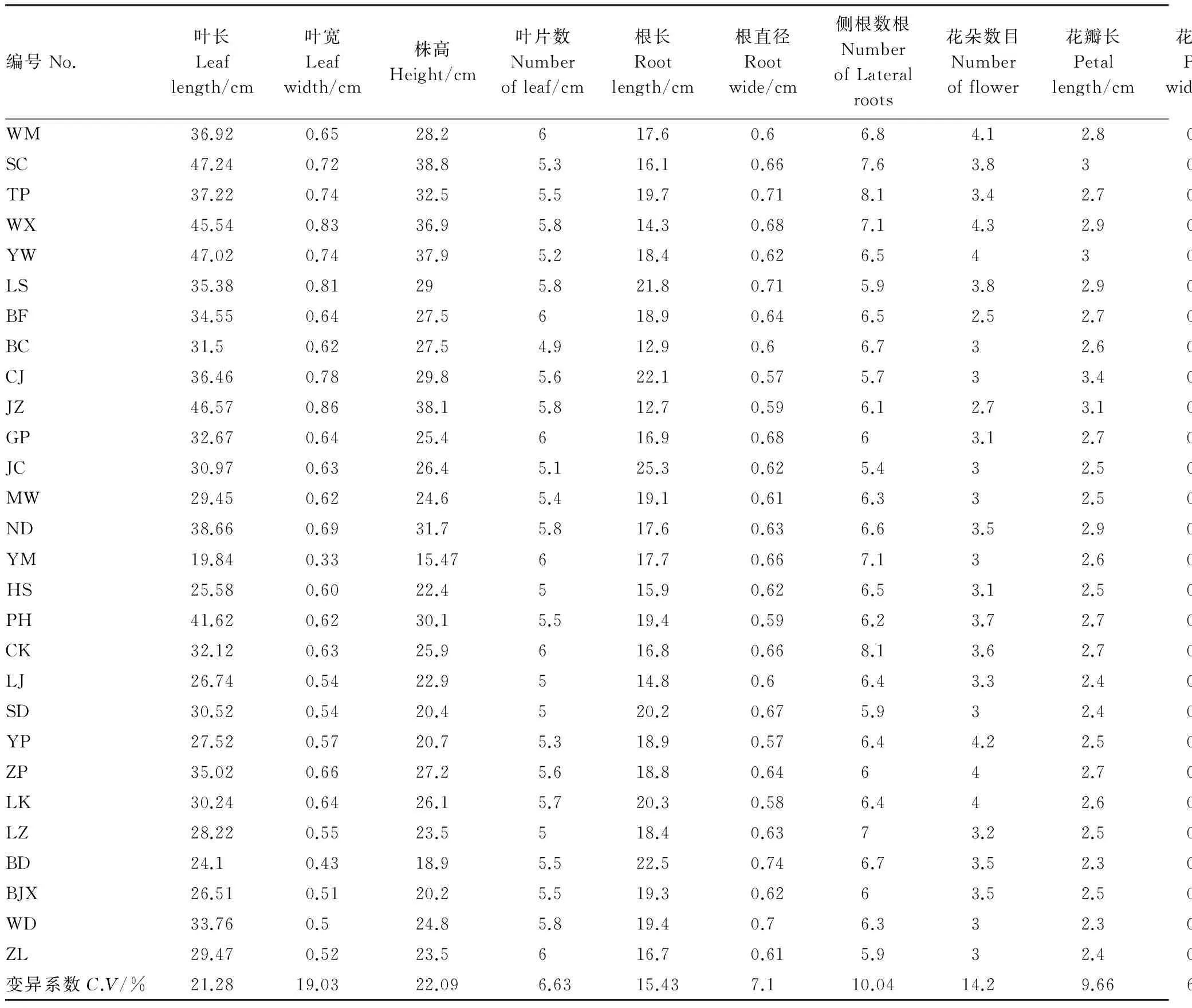
表2 野生莲瓣兰28个居群的形态指标和变异系数
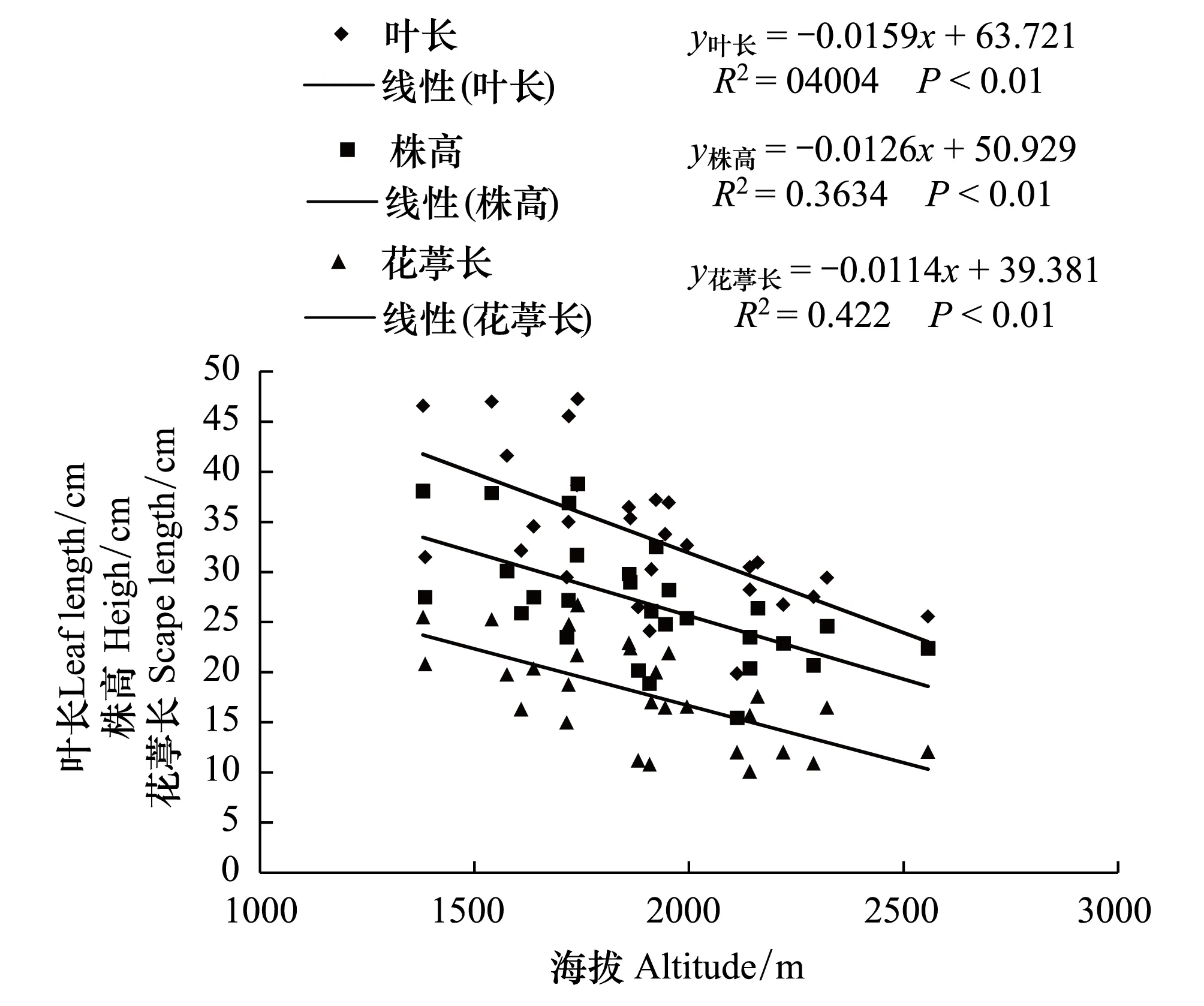
图1 野生莲瓣兰叶长、株高、花葶长与分布地海拔的关系Fig.1 The relationship between leaf length, height, scape length and elevation of wild C. tortisepalum
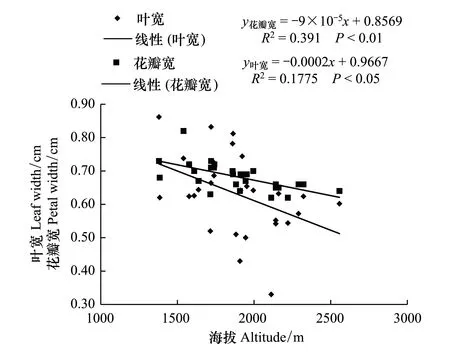
图2 野生莲瓣兰叶宽、花瓣宽与分布地海拔的关系Fig.2 The relationship between leaf width, petal width and elevation of wild C. tortisepalum
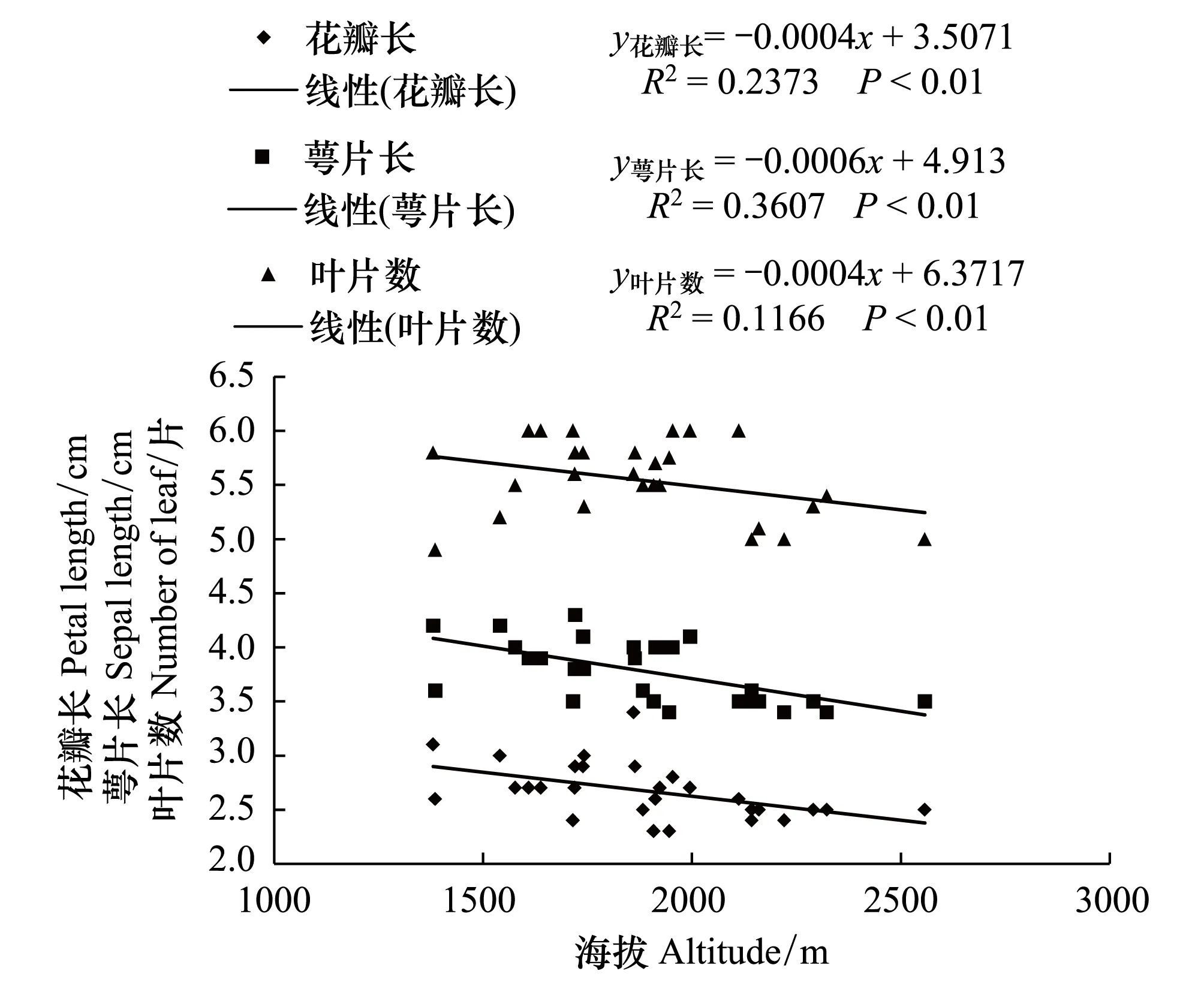
图3 野生莲瓣兰花瓣长、萼片长、叶片数与分布地海拔的关系Fig.3 The relationship between petal length, sepal length, number of leaf and elevation of wild C. tortisepalum

图4 野生莲瓣兰叶长、株高、花葶长与分布地年平均温度的关系Fig.4 The relationship between leaf length, height, scape length and average temperature of wild C. tortisepalum
2.3 分布地相对空气湿度和辐射强度对野生莲瓣兰形态特征的影响
对野生莲瓣兰居群所在地的空气湿度与莲瓣兰形态指标进行了相关性分析(图5,图6)。从图可知, 野生莲瓣兰的叶宽、株高、花葶长均与空气湿度呈显著正相关(P<0.05);叶长与空气湿度呈显著的正相关(P<0.01)。综合表1、图5和图6结果, 28个野生居群分布地空气湿度范围为60.39%—88.42%;表明野生莲瓣兰分布地空气湿度越大, 莲瓣兰的叶片越长越宽, 植株越高, 花葶越长。
对不同野生莲瓣兰居群所在地的辐射强度与形态指标进行了相关性分析, 如图7所示, 叶长、株高均与辐射强度呈显著负相关(P<0.05);根长与辐射强度呈显著正相关(P<0.05)。综合表1和图7结果, 28个野生居群分布地辐射强度范围为4.1—24.8 W/m2;表明分布地辐射强度越大, 叶片越短, 植株越矮, 根系越发达。

图5 野生莲瓣兰叶长、株高、花葶长与分布地空气湿度的关系 Fig.5 The relationship between leaf length, height, scape length and air humidity of wild C. tortisepalum
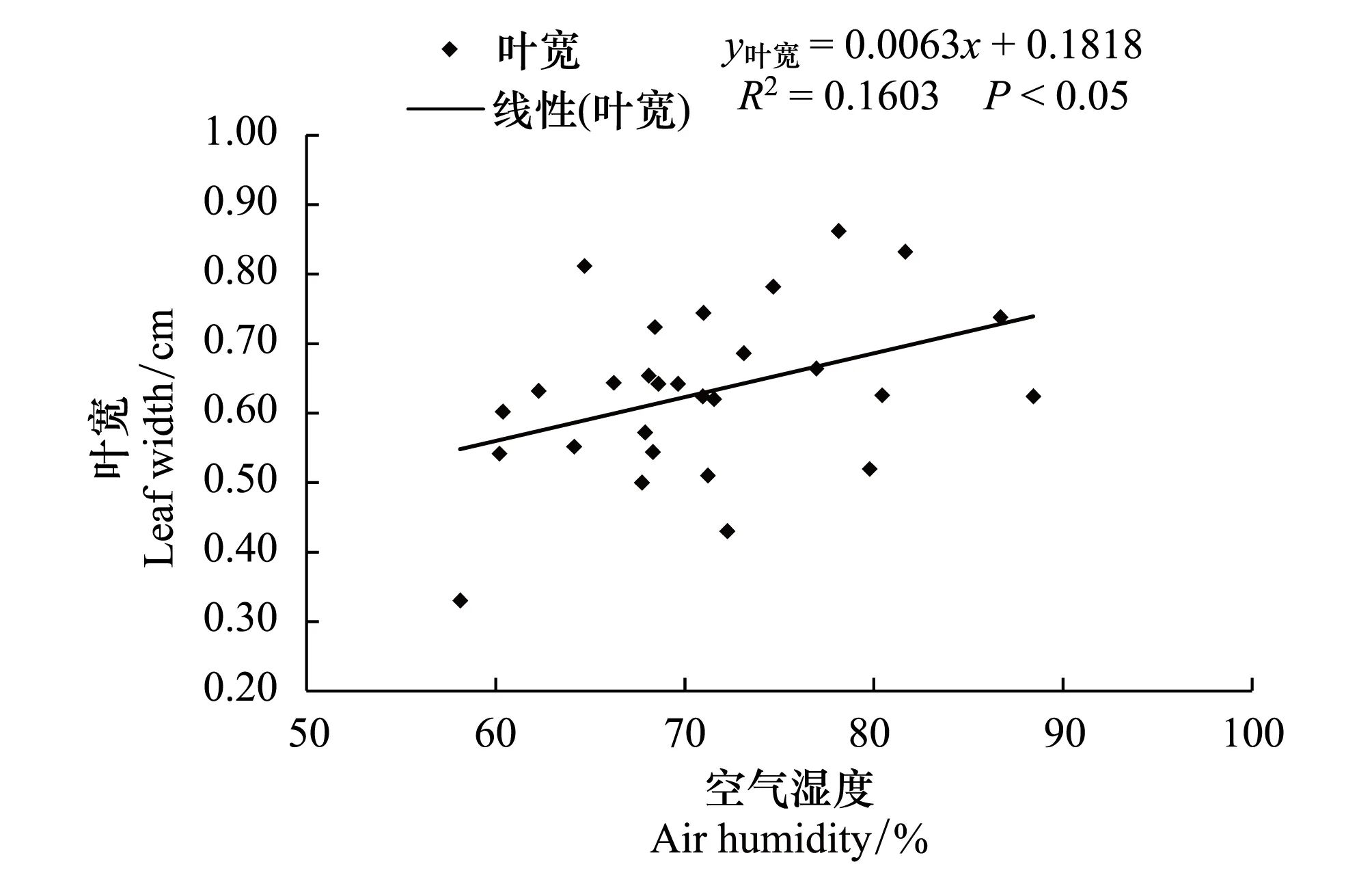
图6 野生莲瓣兰叶宽与布地空气湿度的关系 Fig.6 The relationship between leaf width and air humidity of wild C. tortisepalum

图7 野生莲瓣叶长、株高、根长与兰分布地辐射强度的关系 Fig.7 The relationship between leaf length, height, root length and radiation intensity of wild C. tortisepalum
2.4 野生莲瓣兰形态特征的变异系数分析
云南野生莲瓣兰28个野生居群的形态变异系数见表2。按变异系数从高到低排序, 结果为:花葶长、株高、叶长、叶宽、根长、花朵数、萼片宽、侧根数、花瓣长、萼片长、根直径、叶片数、花瓣宽。 莲瓣兰叶长、叶宽、株高、根长、花葶长的变异系数超过了15%, 其中花葶长的变异系数最大, 为27.77%。这说明云南滇西北莲瓣兰的叶长、叶宽、株高、根长、花葶长变异程度高, 容易受到环境影响。
3 结论与讨论
3.1 野生莲瓣兰在滇西北海拔为1380—2557 m的区域都有分布;随着分布地海拔的升高, 野生莲瓣兰株型矮化, 形态变小。
本研究通过对云南野生莲瓣兰28个野生居群的研究发现, 在滇西北野生莲瓣兰分布地海拔范围为1380—2557 m。随着分布地海拔的升高, 野生莲瓣兰叶长、叶宽、叶片数、株高、花瓣长、花瓣宽、花朵数、萼片长、花葶长都呈现了不同程度的减小趋势。这与叶广[15]和潘光华[16]等对野生莲瓣兰植株高度和叶片宽度与海拔呈负相关关系的研究结果吻合, 但其研究均未涉及野生莲瓣兰花部特征与分布地海拔关系。本研究结果表明, 滇西北野生莲瓣兰分布地海拔幅度远超过叶广[15]等研究报道的1700—2300 m, 以及贾琳[17]等研究报道的1500—2500 m。表明野生莲瓣兰在滇西北有更宽的海拔适应范围;海拔高度不仅影响野生莲瓣兰株型, 而且对莲瓣兰花部特征影响也较大。这些结果对了解野生莲瓣兰资源现状极为重要。
3.2 野生莲瓣兰分布地年均温、空气湿度等生态因子对莲瓣兰形态影响较大, 且呈规律性变化。
植被与气候之间是相互影响的耦合关系[18], 气温和降水长时间的作用是形成区域植被的主要原因[19]。本研究结果表明, 莲瓣兰对生境的年均温、空气湿度和辐射强度就较为敏感, 表现为年均温越高, 空气湿度越大,莲瓣兰的叶片越长越宽, 植株越高、花葶越长;而辐射强度越大, 叶片越短, 植株越矮, 根系越发达。这与刘仲健[20]对墨兰、李艳[21]等对中国兰叶片生长的研究和Mokany[22]等对草原灌丛的研究结果相似。莲瓣兰是一种阴生且喜潮湿的植物, 当空气湿度降低时, 莲瓣兰的叶长、叶宽、株高会有显著的短、细、矮的变化。这与秦栋[23]等对阴生植物蓝果忍冬(LoniceracaeruleaL.)的研究结果:即RH75%处理下, 蓝果忍冬生长良好, 随着空气湿度的下降, 植株株高及相对生长量呈下降趋势相一致。
本研究结果表明, 叶长、株高均与辐射强度有较显著的负相关关系。 根长与辐射有显著的正相关关系, 这说明辐射强度越高, 越不利于野生莲瓣兰的生长。本研究对28个野生莲瓣兰居群分布地辐射强度测定的时间主要集中在11:00—16:00, 强度范围在4.1—24.8 W/m2之间。植物的生长一般会随着光强的增加而增加[24], 然而造成莲瓣兰随光强的增加而减弱的原因可能有以下几方面:(1)强光会导致生境的水分过度蒸腾影响莲瓣兰的生长;(2)强光会伤害莲瓣兰叶片光合系统, 有实验发现阴生植物的叶片在全光照下会发生凋落;(3)强光导致叶片气孔关闭, 减弱蒸腾作用, 影响光合作用的干物质累积[25-26];(4)强光会导致植物分配干物质时, 优先向根分配, 造成叶片得到的干物质量相对减少, 从而影响植物的光合作用及生长[27]。吴能表[28]等研究的阴生植物少花桂(CinnamomumpauciflorumNees), 其幼苗随着生境光照强度的增强, 遮荫条件下根系相对不发达, 与本研究结果相似。向芬[29]等研究发现河谷外阳生生境下的吉首蒲儿根(SinoseneciojishouensisD.G.Zhang, Y.Liu & Q.E.Yang)的比叶面积要显著低于其它阴生生境, 这说明辐射加强, 吉首蒲儿根的叶片会变小, 变厚, 已应对不利的生境胁迫, 这与莲瓣兰对辐射胁迫的应对策略相一致。
3.3 野生莲瓣兰居群形态变异较大, 生态适应幅度变宽, 需加强就地和迁地保护。
在本研究的28个野生莲瓣兰居群进行形态指标的分析发现, 叶长、叶宽、株高、根长、花葶长的变异系数均超过了15%。这与各居群生境遭到人为干扰导致小气候改变有关。祁福云[30]指出, 野生莲瓣兰生境年均温10.5—15.3 ℃, 年降雨量900—1000 mm, 空气湿度60%—80%, 海拔1500—2500 m。本研究结果表明, 滇西北野生莲瓣兰分布地的年均温10—21 ℃, 年降雨量570—1726 mm, 空气湿度58.12%—88.42%, 海拔1380—2557 m。对比发现, 本研究中野生莲瓣兰的生态因子幅度明显扩大, 而且个别调查样地的莲瓣兰植株数量少, 个体矮小, 有明显的退化表征。敖素燕[31]等研究指出, 在野生莲瓣兰种内花部性状较其他性状保持较高的稳定性, 在种内变异系数均未超过15%, 花葶长的变异系数为8.80%。而本研究中花葶长的变异系数最高值可达27.77%, 其他花部形状变异系数未超过15%。造成这一现象的原因可能是各生境辐射强度相差较大, 莲瓣兰为适应环境而发生适应性的改变。
莲瓣兰是适应范围较为狭窄的植物, 由于遭到乱采乱挖导致其生境被破坏, 造成莲瓣兰生态适应幅度变宽。本研究发现部分莲瓣兰居群植株数量少, 植株矮化趋势明显, 表明这些样地并不是其最适宜的生境, 植株有消亡的危险。因此应对这些居群进行就地保护, 恢复其生境。同时, 需要采用自然分株法[32]对其进行迁地保护。
[1] 王鸿泽, 唐敏, 毕玉芬.云南野生莲瓣兰资源研究进展和科研思考. 云南农业大学学报, 2014, 29(3):443-447
[2] 饶箐, 王金亮, 郭树荣. 维西野生莲瓣兰生境变化研究初探. 中国野生植物资源, 2008, 27(4): 17-19.
[3] 黄永艺, 唐敏, 张志荣, 李枝林, 毕玉芬. 云南莲瓣兰主栽品种SSR指纹图谱的构建和遗传差异分析. 热带亚热带植物学报, 2015, 23(3): 236-244.
[4] Tang M, Zeng C X, Bi Y F, Yang J B. Microsatellite markers for the Chinese endangered and endemic orchidCymbidiumtortisepalum(Orchidaceae). American Journal of Botany, 2012, 99(1): e11-e13.
[5] 杨冬之, 刘海娟, 罗毅波, 耿盼盼, 郭彩茹, 周波. 国兰品种分类研究. 安徽农业科学, 2007, 35(29): 9242-9243, 9245-9245.
[6] 敖素燕. 国兰表型性状与核型分析[D]. 临安: 浙江农林大学, 2014.
[7] 刘昳雯, 许春梅, 王丹, 李枝林. 莲瓣兰“滇梅”根状茎的增殖与分化技术研究. 安徽农业科学, 2014, 42(10): 2890-2892.
[8] 李海峰, 杨兴彪, 程清泉. 珍稀濒危莲瓣兰组织培养试管苗移栽的研究. 北方园艺, 2012, (5): 96-98.
[9] 贾琳, 史云东, 虞泓, 李淑英. 莲瓣兰形态变异研究. 植物科学学报, 2013, 31(1): 93-98.
[10] 季祥彪, 王国鼎, 康继川, 乔光, 文晓鹏. 贵州12种兰属植物资源形态学和RAPD标记的比较分析. 种子, 2008, 27(2): 56-60.
[11] 陈定谋, 陈浩. 云南莲瓣兰生育习性、常见品种及栽培管理. 昆明: 云南科技出版社, 2010.
[12] 关文灵, 李枝林, 余朝秀, 王玉英, 程利霞. 云南野生兰花资源的多样性及其引种驯化. 西南农业学报, 2006, 19(4): 688-691.
[13] 刘仲健, 刘可为, 陈利君, 雷嗣鹏, 李利强, 施晓春, 黄来强. 濒危物种杏黄兜兰的保育生态学. 生态学报, 2006, 26(9): 2791-2800.
[14] 罗毅波, 贾建生, 王春玲. 中国兰科植物保育的现状和展望. 生物多样性, 2003, 11(1): 70-77.
[15] 叶广, 唐敏, 王鸿泽, 索默, 车伟光, 陈贤, 毕玉芬. 云南迪庆州野生莲瓣兰形态分异与海拔的关系. 云南农业大学学报, 2015, 30(1): 84-89.
[16] 潘光华. 云南兰花的识别鉴赏与收藏. 昆明: 云南科技出版社, 1994.
[17] 贾琳, 史云东, 虞泓, 周元清. 莲瓣兰繁育方式研究. 玉溪师范学院学报, 2006, 22(12): 84-86.
[18] 郝永萍, 陈育峰, 张兴有. 植被净初级生产力模型估算及其对气候变化的响应研究进展. 地球科学进展, 1998, 13(6): 564-571.
[19] Liang N S, Maruyama K. Interactive effects of CO2enrichment and drought stress on gas exchange and water-use efficiency inAlnusfirma. Environmental and Experimental Botany, 1995, 35(3): 353-361.
[20] 刘仲健, 陈利君, 刘可为, 李利强, 张玉婷, 黄来强. 气候变暖致使墨兰(Cymbidiumsinense)野外种群趋向灭绝. 生态学报, 2009, 29(7): 3443-3455.
[21] 李艳. 几种中国兰的叶片生长动态及栽培基质研究[D]. 南京: 南京林业大学, 2008.
[22] Mokany K, Raison R, Prokushkin AS. Critical analysis of root: s hoot ratios in terrestrial biomes. Global Change Biology, 2006, 12: 84-96.
[23] 秦栋, 段国晟, 霍俊伟, 睢薇. 空气湿度对蓝果忍冬相对生长和外部形态的影响. 北方园艺, 2011, (16): 45-48.
[24] Gardiner E S, Hodges J D. Growth and biomass distribution of cherrybark oak (QuercuspagodaRaf.) seedlings as influenced by light availability. Forest Ecology and Management, 1998, 108(1/2): 127-134.
[25] Gamper R, Mayr S, Bauer H. Similar susceptibility to excess irradiance in sun and shade acclimated saplings of Norway spruce [Piceaabies(L.) Karst.] and stone pine (PinuscembraL.). Photosynthetica, 2001, 38(3): 373-378.
[26] Hanba Y T, Kogami H, Terashima I. The effect of growth irradiance on leaf anatomy and photosynthesis inAcerspeciesdiffering in light demand. Plant, Cell & Environment, 2002, 25(8): 1021-1030.
[27] 林波, 刘庆. 四种亚高山针叶林树种的表型可塑性对不同光照强度的响应. 生态学报, 2008, 28(10): 4665-4675.
[28] 吴能表, 谈锋, 肖文娟, 王小佳. 光强因子对少花桂幼苗形态和生理指标及精油含量的影响. 生态学报, 2005, 25(5): 1159-1164.
[29] 向芬, 周强, 田向荣, 陈功锡, 肖艳. 不同生境吉首蒲儿根叶片形态和叶绿素荧光特征的比较. 生态学报, 2014, 34(2): 337-344.
[30] 祁福云. 维西莲瓣兰资源保护与开发对策. 林业调查规划, 2004, 29(2): 105-108.
[31] 敖素燕, 范义荣, 段艳皊, 郭雨楠, 宁惠娟. 31个国兰品种的数量性状分析. 江苏农业科学, 2014, 42(9): 131-134.
[32] 韩冬. 国兰市场调查及国兰资源保护研究[D]. 北京: 北京林业大学, 2009.
Relationship between morphology and elevation in nativeCymbidiumtortisepalumin Yunnan Province, China
SUO Mo1, BI Yufen1,*, WANG Hongze1, TANG Min2, YE Guang2
1CollegeofAnimalScienceandTechnology,YunnanAgriculturalUniversity,Kunming650201,China2CollegeofLandscapeandHorticulture,YunnanAgriculturalUniversity,Kunming650201,China
Cymbidiumtortisepalumis an endemic and endangered orchid species in southwestern China. Twenty-eight native populations ofCymbidiumtortisepalumin northwest Yunnan Province were studied from August to October in 2013 and 2014. NativeCymbidiumtortisepalumwere distributed in elevations 1380—2557 m; The number of leaves, scape length, petal length, petal width, and sepal length were all negatively correlated with altitude (P<0.01); leaf length (P<0.05), height (P<0.05), and scape length (P<0.01) were all positively correlated with the average annual temperature. Leaf width (P<0.05), height (P<0.05), scape length (P<0.05), and leaf length (P<0.01) were all positively correlated with air humidity. Moreover, it was observed that the higher the altitude, the smaller the plant; the higher the average temperature or air humidity, the longer and wider the leaf blade; the taller the plant, the longer the scape; the stronger the radiation intensity, the shorter the blade; and the more dwarfed the plant, the more developed the root. The great changes among populations in wildCymbidiumleaf length, leaf width, plant height, root length and scape length. The coefficient of variation was greater than 15%, which indicates that there is a wide range of ecological adaptability, a large variation in morphology and the plant dwarfing serious, it is urgent to strengtheninsituandexsituconservation. The effects of ecological factors, including altitude and temperature, on the characteristics of wildCymbidiumtortisepalumprovide a theoretical basis for the further protection of wildCymbidiumtortisepalum.
wild cymbidium; altitude; annual average temperature; humidity; radiation intensity
国家自然科学基金项目(31260487)
2015- 07- 26;
日期:2016- 04- 19
10.5846/stxb201507261561
*通讯作者Corresponding author.E-mail: biyufenynnd@ sina. com
索默, 毕玉芬, 王鸿泽, 唐敏, 叶广.云南野生莲瓣兰形态分异与海拔的关系.生态学报,2016,36(19):6170- 6177.
Suo M, Bi Y F, Wang H Z, Tang M, Ye G.Relationship between morphology and elevation in nativeCymbidiumtortisepalumin Yunnan Province, China.Acta Ecologica Sinica,2016,36(19):6170- 6177.
猜你喜欢
花卉(2024年7期)2024-04-11 03:05:36
陶瓷科学与艺术(2022年11期)2022-12-24 08:10:20
理财·收藏版(2021年8期)2021-10-12 07:13:59
江苏陶瓷(2020年1期)2020-03-20 09:59:56
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
江苏农业学报(2018年5期)2018-09-10 22:36:44
江苏农业科学(2016年10期)2017-02-05 14:45:09
家家乐·上半月(2013年6期)2013-04-29 00:44:03
