
腾格里沙漠南缘沙地一年生植物的生态位分析
2016-11-28 03:30:05蒲万旭张德魁
甘肃科技 2016年16期
蒲万旭,张德魁
(金塔县治沙研究试验站,甘肃 酒泉 735300)
腾格里沙漠南缘沙地一年生植物的生态位分析
蒲万旭,张德魁
(金塔县治沙研究试验站,甘肃 酒泉 735300)
为进一步研究沙地生态系统稳定性提供依据,揭示沙地植被不同演替阶段下一年生植物的生态位变化规律。本文以腾格里沙漠南缘沙地植被为背景,在流动沙地、半流动沙地、半固定沙地和固定沙地4个较典型演替阶段上选取了一年生植物种群为研究对象,计测分析其生态位宽度、生态位重叠和生态位相似比例。结果表明:腾格里沙漠南缘流动沙地上,沙米是一年生植物优势种;半流动沙地上,沙米仍是一年生植物优势种;半固定沙地上,狗尾草和小画眉草是一年生优势植物种;固定沙地上,刺蓬、画眉草和狗尾草是一年生优势植物种。
腾格里沙漠;一年生植物;生态位宽度;生态位重叠
1 概述
“生态位”(niche)是研究植物生态和群落生态的重要理论问题(袁志忠和何丙辉,2004),是指植物种群在时间、空间的位置以及种群在群落中的地位和功能作用(尚玉昌,1988)。生态位宽度可以作为物种对环境资源利用多样性的一种测度,生态位相似比例可以作为物种对环境资源共享程度的一种衡量指标,生态位重叠可以作为植物种间生态学相似性的测度。从环境变化梯度着手,分析植物种群在这些变化梯度上的分布规律是研究植物种群生态位的重要方法(张林静等,2002)。
一年生植物是干旱沙区沙地上分布最广泛的植物种类,并常以先锋植物或建群种植物占据在沙地植被演替的各个阶段,具有特定的生理学特性(张景光等,2002;梁存柱等,2003),在各种类型的沙地生态系统中扮演着重要的生态功能角色(唐进年等,2006)。因此,研究沙地一年生植物的生态位特征,有利于了解沙区一年生植物在沙地植物群落中的功能和地位。本文以腾格里沙漠南缘沙地植被为背景,在流动沙地、半流动沙地、半固定沙地和固定沙地4个较典型演替阶段上(刘华民等,2005;赵丽娅等,2003),选取了植物群落主要一年生植物种群为研究对象,计测分析了它们的生态位宽度、生态位重叠和生态位相似比例,旨在揭示沙地植被不同演替阶段下一年生植物的生态位变化规律,为进一步研究沙地生态系统稳定性提供依据。
2 研究地区与研究方法
2.1 研究区概况
研究区位于腾格里沙漠南缘,由沙漠向绿洲的过渡带上,地理位置37°32′~37°36′N,103°38′~103° 42′。区内气候属大陆性干旱气候,多年平均气温6℃,多年平均降雨量175mm,地下水位埋深平均67m。地貌类型为风沙地貌,土壤类型以风沙土为主,依据植被的不同演替时间阶段,将带内的沙地划分为流动沙地、半流动沙地、半固定沙地和固定沙地4个类型,沙地海拔在1730~1810m的范围内;在流动沙地上,主要植物种是沙蒿(Artemisia desertorum)、沙米(Agriophyllum squarrosum)和沙鞭(Psammochloa villosa),半流动沙地上是沙蒿、沙米、油蒿(Artemisia ordosica)、雾冰藜(Bassia dasyphylla)等,半固定沙地上是油蒿、狗尾草(Setaria viridis)、小画眉草(Eragrostis minor)、雾冰藜等,固定沙地上是油蒿、刺蓬(Cornulaca alaschanica)、小画眉草等,构成的植被中植物种类组成单调,具有典型的沙旱生特性。
2.2 样方的设置与调查方法
采用样方法进行调查。在研究区内,样方分别设置在流动沙地、半流动沙地、半固定沙地和固定沙地上,样方大小1m×2m,每个类型沙地设置6个样方,共计24个,对样方内的所有植物进行调查,调查指标包括种名、冠幅和盖度。由于研究对象主要针对的是草本植物,它们的个体矮小,个体数量变化比较大,采用盖度作为指标能较好地代表和反映该种群
在群落中的实际情况 (王刚等,1984;张林静等,2002),因此本文以盖度作为数量性状进行生态位宽度值和生态位重叠值的计算,进行生态位特征分析。
2.3 数据处理与分析
4种沙地类型的所有数据资料用 Microsoft Excel电子表格软件和SPSS统计软件做统计分析。
生态位宽度采用香农-维纳 (Shannon-Wiener)生态位宽度公式(楚光明等,2005),计算公式为:
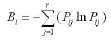
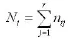
生态位相似比例 (谢少鸿等,2006;奚为民,1993)计测公式:
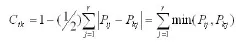
式中,Cik表示物种i与k的相似程度,且有Cik=Cki,Pij和Pkj分别为种i和种k在资源位j上的数量特征值。
种间的生态位重叠采用Pianka(1973)公式计测:
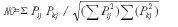
式中,Pij和分别为种i和种k在资源位j上的数量特征值(唐启义和冯明光,1997;楚光明等,2005)。
3 结果与分析
3.1 4种沙地类型的差异显著性检验
4种沙地类型的盖度数量性状原始数据经Microsoft Excel电子表格整理后,首先用SPSS统计软件做方差齐次检验 (Test of Homogeneity of Variances),Levene统计量为1.88,第1自由度等于3和第2自由度等于20下对应的概率值 (P值)为0.166,大于0.05,因此认为原数据是方差齐的;其次做沙地类型对盖度的单因素方差分析(ANOVA)(表1),表中显著性水平均为0,说明各沙地类型间至少有1组或4组间差异显著;再进一步做沙地类型间的逐对比较(LSD法)(表2),表2的比较结果说明,4种类型沙地两两各组均值间均有显著性差异,表中是用“*”标示的组均值在0.05水平上有显著性差异。
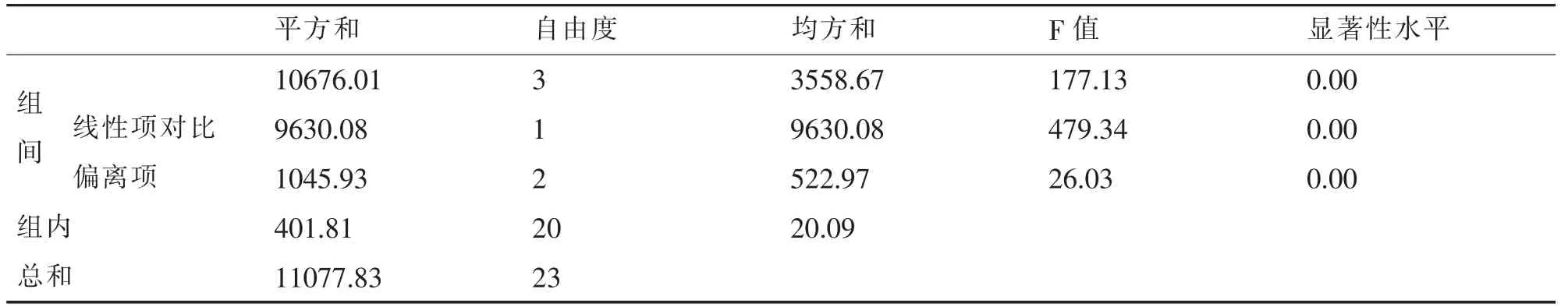
表1 方差分析表
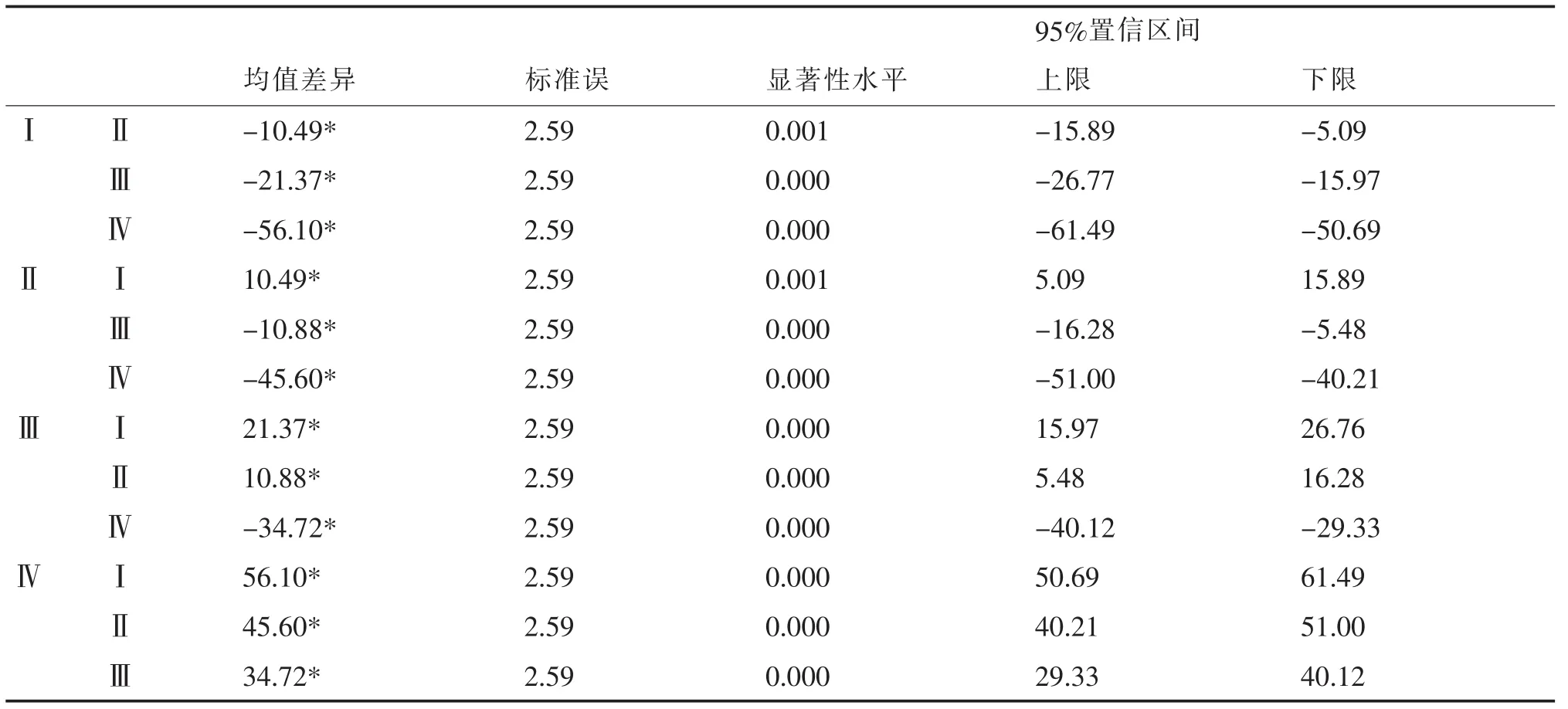
表2 LSD法均值多重比较表
3.2 生态位宽度
生态位宽度主要反映物种对资源利用的程度,物种生态位宽度越大,说明该物种的生态适应能力越强、生存机会越大、分布越广(张继义等,2003)。从表3中可知,在腾格里沙漠南缘的流动沙地中,主要植物种有沙蒿、沙米和沙鞭三种植物,一年生植物沙米的生态位宽度值 (Bi= 2.6340)。
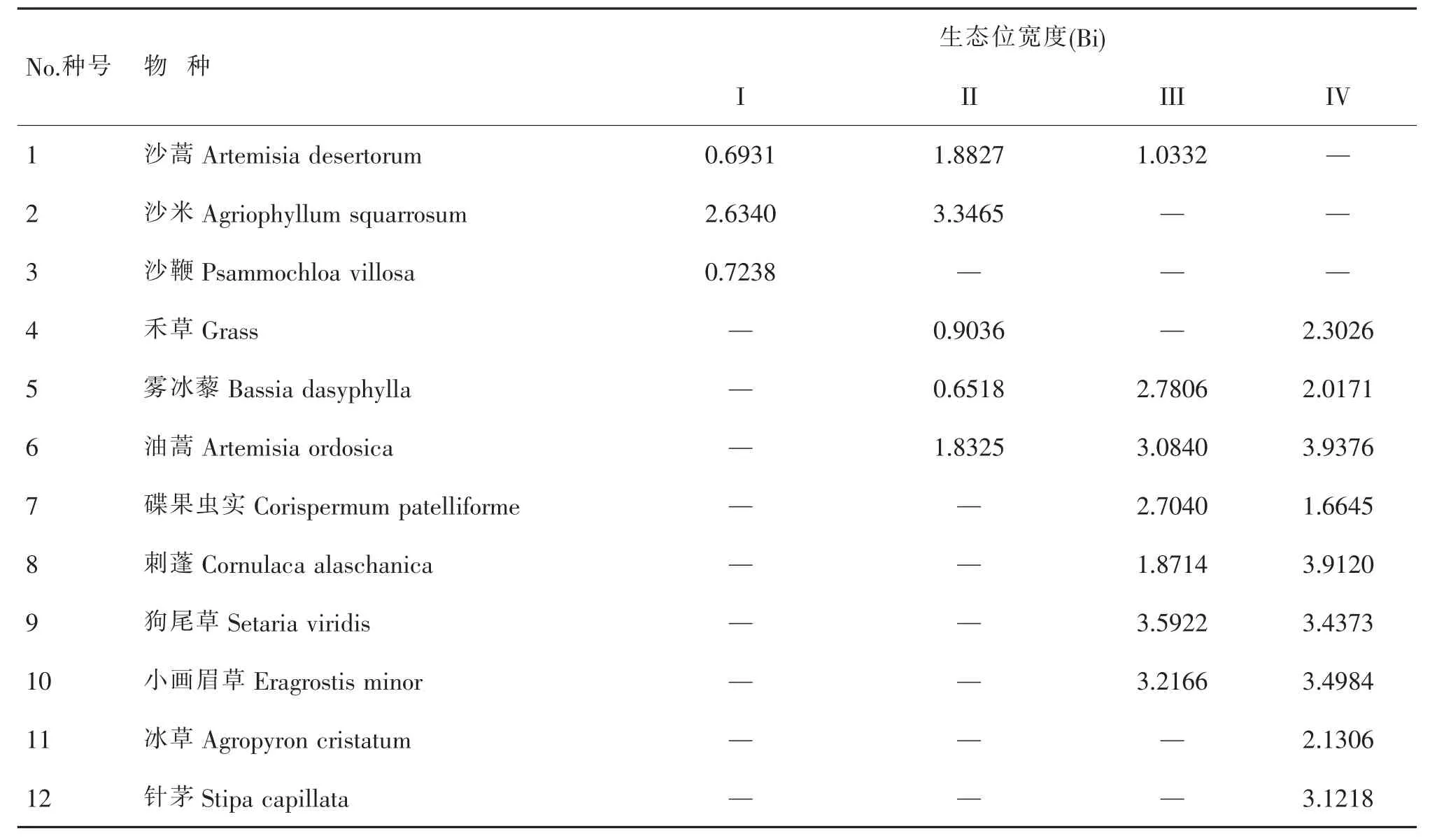
表3 不同沙地一年生植物的生态位宽度值
要远远大于沙鞭的生态位宽度值 (Bi=0.7238)和半灌木植物沙蒿的生态位宽度值(Bi=0.6931);在半流动沙地中,生态位宽度值最大的仍是沙米(Bi= 3.3465),其次是沙蒿 (Bi=1.8827)和油蒿 (Bi= 1.8325),雾冰藜的生态位宽度值最小(Bi=0.6518);在半固定沙地中,狗尾草的生态位宽度值最大(Bi= 3.5922),其次是小画眉草(Bi=3.2166)和油蒿(Bi= 3.0840),生态位宽度值最小的是刺蓬(Bi=1.8714)和沙蒿(Bi=1.0332);在固定沙地中,生态位宽度值最大的是油蒿(Bi=3.9376)和刺蓬(Bi=3.9120),生态位宽度值最小的是雾冰藜 (Bi=2.0171)和碟果虫实(Corispermum patelliforme)(Bi=1.6645)。
表3中还可以看出,对于同一种一年生植物,在不同类型的沙地中它的生态位宽度值会有较大的变化,如沙米,仅存在于流动沙地和半流动沙地中,并且生态位宽度值在这两种类型的沙地中均最大,半流动沙地上沙米对环境因子的利用要比流动沙地上高,这是由于沙米种子的定居和发芽需要借助别的植物对流沙具有一定的固定才能进行,但随着生境的变化,到半固定沙地和固定沙地中就退出生长;雾冰藜从半流动沙地有生长,到半固定沙地对环境因子的利用达到最高,再到固定沙地,它对环境因子的利用又趋减弱;碟果虫实和狗尾草只有在半固定沙地和固定沙地上生长,且在半固定沙地上对环境因子的利用要比在固定沙地上高;刺蓬和小画眉草也只有在半固定沙地和固定沙地上生长,但它们在固定沙地上对环境因子的利用要比半固定沙地上高。因此,从生态位宽度来看,流动沙地和半流动沙地上,一年生植物沙米是优势种;在半固定沙地上,一年生植物碟果虫实和狗尾草是优势种;在固定沙地上,一年生植物刺蓬和小画眉草是优势种。
3.3 生态位相似比例
从表4中列出的不同类型沙地一年生植物的生态位相似比例值可知,在流动沙地,3种植物的生态位相似比例在 1.8000~2.2193之间;在半流动沙地,5种植物的生态位相似比例,有1对 等 于 7.8282,1对 等 于 6.9434,1对 等 于4.9048,其余7对都介于1.7888~2.9053之间;在半固定沙地。
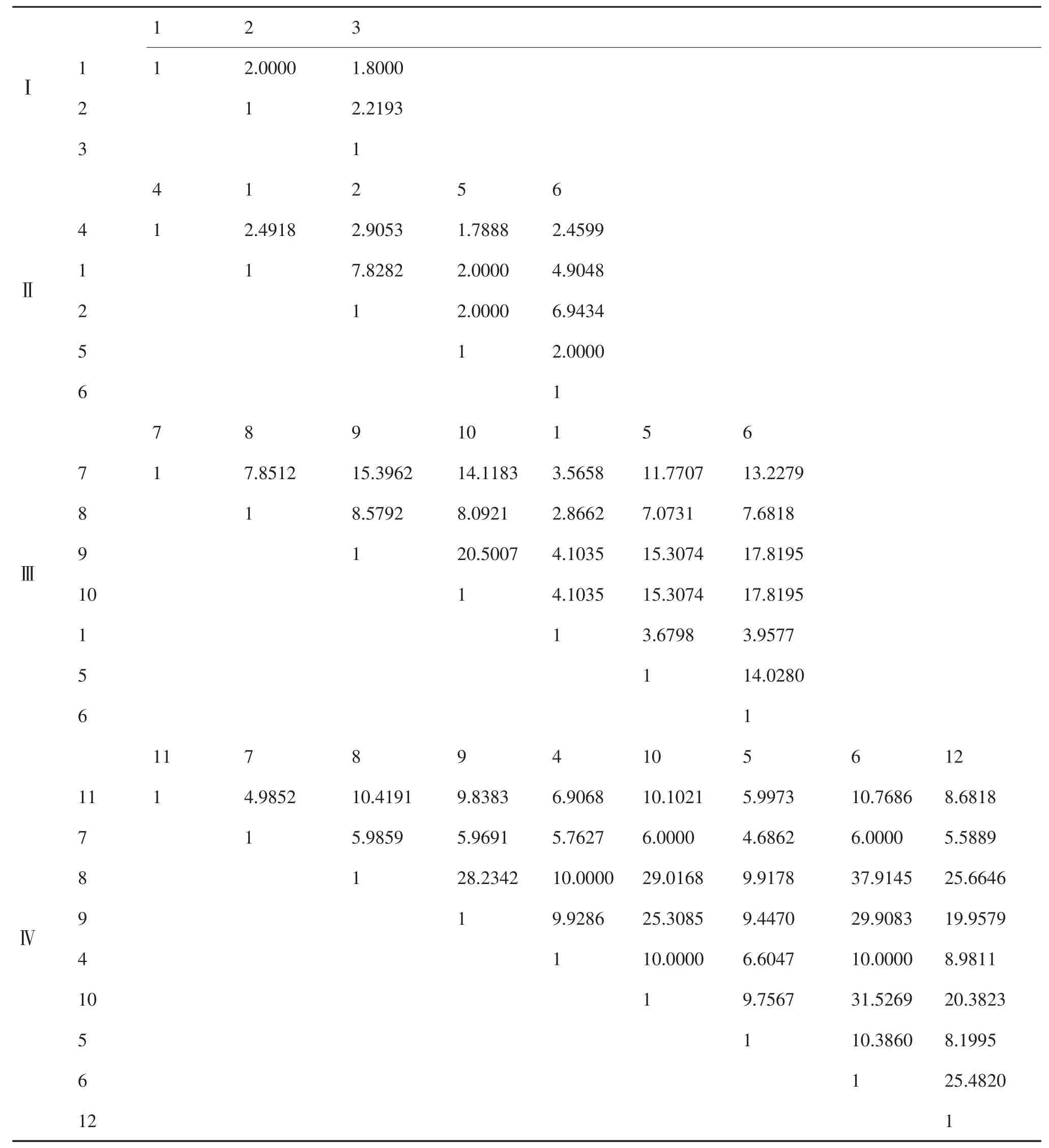
表4 不同沙地一年生植物的生态位相似比例
生态位相似比例值大于20.0000的有1对,介于10.0000~20.0000之间的有9对,介于5.0000~10.0000之间的有 5对,下剩 6对介于2.8662~5.0000之间;在固定沙地上,生态位相似比例值大于30.0000的有2对,介于20.0000~30.0000的有7对,介于10.0000~20.0000的有8对,介于5.0000~10.0000的有17对,下剩2对介于4.5000~5.0000之间,反映出一年生植物种利用资源的相似程度大。不同的一年生植物有各自适宜的不同沙地类型,在各自适宜的沙地类型中,一年生植物之间或与其它植物之间有较大的生态位相似比例,表明一年生植物在不同类型的沙地中,有明显的优势种群。另外,表3中的生态位宽度值和表4中的生态位相似特征值反映出,生态位宽度值较大的两个植物种,种对相似性比例值一般较高,如半固定沙地中的狗尾草与小画眉草、固定沙地中的刺蓬与油蒿、小画眉草与油蒿等,它们的生态位宽度值大,种
对的相似性比例也高。
3.4 生态位重叠
表5列出了一年生植物在各类型沙地的生态位重叠。在流动沙地上,重叠较大的种对有沙米和沙蒿(4.6678),沙米和沙鞭(4.6157),沙米在流动沙地上的生态位最宽,它与其他种间的生态位重叠也大;在半流动沙地上,沙米与沙蒿生态位重叠最大(10.1103),其次是沙米与油蒿(10.0535)。

表5 不同类型沙地一年生植物的生态位重叠值
表明沙米仍是半流动沙地上的优势种;在半固定沙地上,狗尾草和小画眉草的生态位重叠最大(17.4361),其次是小画眉草与油蒿(16.5108)、狗尾草与油蒿(16.3624),表明狗尾草与小画眉草这两个优势种在适应半固定沙地生境方面具有相似的特性;在固定沙地上,刺蓬与油蒿的生态位重叠值最大(37.6122),其次是小画眉草与油蒿(35.6992)、狗尾草与油蒿 (33.0439),说明在适应固定沙地生境方
面,刺蓬、小画眉草具有相似的特性。从物种竞争的角度看,生态位重叠越大表明物种之间的竞争排斥作用越强烈,但一年生植物能够同时出现在某一类型沙地植物群落中,且形成共优状态,如半固定沙地上的狗尾草与小画眉草,说明它们在其他资源轴上生态位特化.使它们能够彼此共存。
4 结论与讨论
1)通过对腾格里沙漠南缘不同类型沙地上一年生植物生态位特征的研究,生态位特征值在一定程度上可以表征植物种对环境的适应性及利用资源的能力,即在某一小环境中,物种的生态位越宽,该物种利用小环境资源的能力越强。根据生态位的大小,沙米为流动沙地和半流动沙地上的一年生植物优势种,狗尾草与小画眉草是半固定沙地上的优势种,刺蓬与小画眉草是固定沙地上的优势种。随着生境的变化,这些优势植物种的生态位特征值会有较大变化,即一年生植物在沙地不同的小生境上表现出较大的分布分化,这有助于它们更充分利用资源。
2)不同类型沙地上优势种对的生态位相似性比例值较大,反映出沙地上一年生植物优势种利用资源的相似程度较大,这与沙地中一年生植物种群的位置、不同类型沙地的生境、一年生植物利用资源上的分化有关。
3)生态位重叠体现了物种对同等级资源的利用程度以及空间配置关系,一般而言,2个生态位宽的物种之间生态位重叠值高(陈波与周兴民,1995;郭全邦等,1997),而2个生态位较窄的物种之间或1个生态位窄的物种与1个生态位宽的物种之间的重叠值则较低 (刘金福和洪伟,1999;柳江等,2002)。腾格里沙漠南缘一年生植物种之间的生态位重叠情况与前人的这些研究结论相一致。生态位重叠值较大的种对之间存在有两种可能,一种是植物种间共享资源的同时存在竞争关系,二是植物种间资源利用相似的同时彼此促进关系 (李瑞等,2006)。在流动沙地和半流动沙地上,后者的可能性大,如沙米与沙蒿之间,沙米的定居和生长需要借助沙蒿对流沙的适量固定,沙米的生长又为沙蒿的更新创造条件,二者之间存在彼此的相互促进关系,在半固定和固定沙地上,更多倾向于前者,如狗尾草、画眉草、刺蓬等在共享资源的同时又相互之间存在竞争关系,特别是相互之间对水分的竞争更为强烈。
4)生态位与植物种群是一一对应的,某一特定的植物种群要求某一特定的生态位,反过来某一特定的生态位只能容纳某一特定的植物种群(袁志忠和何丙辉,2004)。通过对物种生态位宽度、生态位相似性比例以及生态位重叠的研究分析,有助于我们了解一年生植物在不同类型沙地中对资源利用的差异。对于沙地生境条件而言,植物在生长中可有效利用的资源有限,优势植物种在创建植物群落过程中形成了各自内部独特的生境条件,这一独特的生境条件就决定了不同的群落内植物种类的组成,所形成的小环境内,这些植物种的生活力及生态适应能力较强,一年生植物的生活周期短、繁殖比较迅速,因而表现出较强的优势,但只要生境稍有改变,它们的生态位就会有较大的变化,远离小生境,生态位迅速变窄,导致生态位重叠程度的降低,种间对资源的利用上出现分化。
[1] 陈波,周兴民.三种蒿草群落中若干植物种的生态位宽度与重叠分析[J].植物生态学报,1995,19(2):158-169.
[2] 楚光明,潘存德,晋瑜,等.克拉玛依农业综合开发区外围荒漠植被主要植物种的生态位分析[J].新疆农业科学, 2005,42(1):9~13.
[3] 郭全邦,刘玉成,李旭光.缙云山森林次生演替序列优势种群生态位[J].西南师范大学学报(自然科学版),1997,(22):3-78.
[4] 李瑞,张克斌,杨俊杰,等.宁夏盐池荒漠化草原人工封育区生态位研究[J].干旱区资源与环境,2006,20(4):171-175.
[5] 梁存柱,刘钟龄,朱宗元,等.阿拉善荒漠区一年生植物层片物种多样性及其分布特征[J].应用生态学报,2003,14 (6):897-903.
[6] 刘华民,朴顺姬,王立新,等.不同演替阶段褐沙蒿种群特征研究[J].生态学杂志,2005,24(5):497-502.
[7] 刘金福,洪伟.格氏栲群落生态学研究—格氏栲林主要种群生态位的研究[J].生态学报,1999,19(3):347-352.
[8] 柳江,洪伟,吴承祯,等.退化红壤区植被恢复过程中灌木层主要种群的生态位特征[J].植物资源与环境学报,2002,11(2):11-16.
[9] 尚玉昌.现代生态学中的生态位理论[J].生态学进展, 1988,5(2):77-84.
[10]唐进年,彭金凤,徐先英,等.一年生荒漠植物的生态功能及在沙地植被恢复与重建中的作用[J].甘肃科技,2006.22 (10):212-215. [11]唐启义,冯明光.实用统计分析及其计算机处理平台[J].北京:中国农业出版社,1997.
[12]王刚,赵松岭,张鹏云.关于生态位定义的探讨及生态位重叠计测公式改进的研究[J].生态学报,1984,4(2):119-127.
[13]王琳,张金屯.濒危植物矮牡丹的生态位研究[J].生态学杂志,2001,20(4):65~69.
[14]奚为民.怀柔山区灌丛群落优势种群生态位的研究[J].植物生态学与地植物学学报,1993,17(4):324-330.
[15]谢少鸿,陈玉军,陈远合,等.广东南澳岛台湾相思林主要种群生态位研究[J].生态科学,2006,25(4):343-345.
[16]袁志忠,何丙辉.生态位理论及其在植物种群研究中的应用[J].福建林业科技,2004,31(2):123-127.
[17]张桂莲,张金屯.关帝山神尾沟优势种生态位分析[J].武汉植物学研究,2002,20(3):203-208.
[18]张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征[J].生态学报,2003,23(12): 2741-2746.
[19]张景光,周海燕,王新平,等.沙坡头地区一年生植物的生理生态特性研究[J].中国沙漠,2002,2(4):350-353.
[20]张林静,岳明,张远东,等.新疆阜康绿洲荒漠过渡带主要植物种的生态位分析[J].生态学报,2002,22(6):969-972.
[21]张林静,岳明,赵桂仿.生态位不同计测方法在绿洲荒漠交错带应用的比较分析[J].生态学杂志,2002,21(4):71-75.
[22]赵丽娅,李锋瑞,王先之.草地沙化过程地上植被与土壤种子库变化特征[J].生态学报,2003,23(9):1745-1756.
[23]Pianka ER.1973.The structure of lizard Communities.Ann RevSyst,4:53~74.
S728.4
猜你喜欢
军事文摘(2021年18期)2021-12-02 01:27:58
内蒙古林业(2021年6期)2021-06-26 10:42:32
幼儿智力世界(2021年4期)2021-05-17 17:10:29
作文周刊·小学一年级版(2020年40期)2020-10-19 04:42:20
阅读与作文(小学低年级版)(2018年9期)2018-10-26 10:53:24
阅读与作文(小学低年级版)(2018年10期)2018-10-25 12:26:00
科普童话·百科探秘(2018年7期)2018-09-13 11:12:08
意林·少年版(2017年15期)2017-09-05 19:46:51
小学生作文选刊·低年级版(2016年6期)2016-06-24 08:40:46
小星星·阅读100分(低年级)(2014年1期)2014-02-19 00:32:19
