
大豆中不同除草剂作用靶标酶的miRNA前体克
2016-11-28 01:47陶波刘洋李向勇金龙国邱丽娟
江苏农业科学 2016年9期
陶波+刘洋+李向勇+金龙国+邱丽娟


摘要:对前人已预测的大豆miRNA的4个茎环结构进行引物设计、克隆miRNA前体,先将设计好的接头与大豆小RNA的3′末端连接,然后利用与接头序列互补的引物对小RNA进行反转录,即可得到加接头的第1链cDNA,再用基因特异引物GSP和通用接头引物的组合,经PCR扩增后即可捕获位于已知序列区和接头之间的3′端RNA序列。结合3′端引物的测序结果,初步预测成熟miRNA可能的6个组合是同一家族。
关键词:miRNA;EPSPS(5-烯醇丙酮酰莽草酸-3-磷酸合成酶);基因序列;草甘膦;克隆
中图分类号: S482.4 文献标志码: A
文章编号:1002-1302(2016)09-0024-04
草甘膦是一类具有特异性,且能有效抑制叶绿体酶、5-烯醇丙酮酰莽草酸-3-磷酸合成酶(EPSPS)的除草剂[1-2]。对EPSPS作用的草甘膦能阻断莽草酸酯途径,通过竞争性抑制磷酸烯醇式丙酮酸(PEP)与EPSPS结合,形成稳定的三元络合物,从而阻断PEP与3-磷酸莽草酸(S3P)反应生成EPSPS,最终阻止芳香族氨基酸的生物合成,抑制芳香族氨基酸的生成,导致植物死亡[3-4]。
microRNA(miRNA)是指真核生物中长度约为21~25 nt的小型内源的非编码单链RNA家族。microRNA是Lee等于1993年在线虫中首次发现的。此后,大量的microRNA在植物和动物中被发现和克隆[5],到目前为止,植物中发现的microRNA分子已逾200种[6]。microRNA是近年来非编码RNA研究的热点。它们在组织发育和细胞分化的特定阶段表达,可以在转录后水平对许多基因的表达起着重要的调控作用[7]。miRNA通过2种途径发挥其作用:(1)miRNA与靶mRNA序列不完全互补,它主要通过与靶mRNA的不完全互补配对进而阻抑mRNA的翻译过程,这种途径对mRNA的稳定性无任何影响,属于翻译水平基因调节;(2)miRNA与靶mRNA序列完全互补,它可以通过与小干扰RNA(small interfering RNA,siRNA)类似的途径与靶mRNA完全互补配对进而降解靶mRNA,属于转录后水平基因调节[8]。
在各类小分子RNA中,miRNA具有最广泛的基因调节功能[9],内含子miRNA具有反向调节其宿主基因的功能[10-11],可能有以下几种情况:(1)当宿主基因是内含子miRNA的靶基因时,内含子miRNA直接下调宿主基因的表达,抑制其功能的发挥,这是直接作用。譬如,内含子miR-204被预测可直接靶向其宿主基因[WTBX][STBX]TRPM3[WTBZ][STBZ][12]。(2)内含子miRNA还可以通过下调促进宿主基因功能的靶基因,从而拮抗宿主基因的功能。目前还未见有这方面的试验证据,但不能否定这种情况存在的可能性。文献报道内含子miR-25的可能靶基因是[WTBX][STBX]FHL2[WTBZ][STBZ]基因,而FHL2蛋白则通过与宿主基因编码的蛋白MCM7相互作用从而促进其功能[13]。因此,我们推测内含子miR-25有可能有反向调节宿主基因[WTBX][STBX]FHL2[WTBZ][STBZ]的功能,当然这还有待今后进一步试验证实。拟南芥中的miR160通过负调控生长素响应因子(auxin response factors)ARF10和ARF16的表达而控制根冠细胞的形成[14]。玉米中的GL15活性增高不仅使幼年期叶片增多,而且延缓生殖发育,而miR172对GL15mRNA水平进行负调控,从而促进玉米幼叶向成熟叶片转变[15]。贾小云等发现,miR828可以通过靶向SlMyb7-like负调控花青素合成相关基因的表达,进而负调控番茄植株中花青素的生物合成[2]。这些都表明植物中miRNA具有负反馈调节机制[16]。
本研究对前人预测的除草剂作用靶标酶的大豆miRNA的4个茎环结构进行引物设计,克隆miRNA前体,从而进一步研究它们在植物生长发育中的生物学意义和分子水平上的调节机制,而且还可以有目的、有计划地利用它们进行植物生长发育的调控和生理结构的改造,使植物更大限度地表现出对人类研究有利的性状,从而为人类生产生活造福。
1 材料与方法
1.1 供试材料
大豆基因组EST序列数据库来源:http://www.ncbi.nlm.nih.gov/;RNA-fold predict:http://frontend.bioinfo.rpi.edu/zukerm/home.html;Mipred confirm:http://www.bioinf.seu.edu.cn。
1.2 试验方法
利用生物信息学方法,对不同除草剂作用靶标酶EST序列进行miRNA预测。通过RNA-fold predict软件对原始EST序列预测,初步得到12个microRNA二级结构,利用Mipred confirm(http://www.bioinf.seu.edu.cn)进一步筛选,共筛选到4个miRNA前体,其中2个EPSP合成酶、1个谷氨酰胺合成酶、1个原卟啉原氧化酶。
本试验中,先将设计好的接头与大豆小RNA的3′末端连接,然后利用与接头序列互补的引物对小RNA进行反转录,即可得到加接头的第1链cDNA;再用基因特异引物GSP和通用接头引物的组合,经PCR扩增即可捕获位于已知序列区和接头之间的3′ 端RNA序列。
1.2.1 连接3′接头 3′接头序列:5′-PO4-UUUCACTAGCACGAGCTCGCTACGCTCACTACTCGGCATTATGTACGCTAACTAACTCGTGGTC(idT)-3′。首先将总RNA与3′接头均匀混合,80 ℃下处理2 min后置于冰中冷却。配制反应体系:无核酸酶的水18.6 μL、10×T4 RNA 连接酶缓冲液[不含牛血清白蛋白(BSA-free)]4.0 μL、BSA(0.1%)2.4 μL 、3′接头2.0 μL、总RNA 11 μL、T4 RNA连接酶(40 U/μL)2.0 μL(100 μmol/L),5 ℃反应16 h后,加入2 μL 0.5 mol/L EDTA终止反应;70 ℃下处理10 min以灭活连接酶,之后置于冰中保存。需要注意的是,配制反应体系时须按照以下顺序:无核酸酶的水→10×T4 RNA连接酶缓冲液→0.1% BSA→总RNA & 3′接头混合液。
1.2.2 连接5′接头 5′接头序列:5′-GGAATTCGGATC-3′,通过TaKaRa生产的末端转脱氧核苷酸转移酶作用,使5′接头poly(G)连接到cDNAs的3′末端,随后利用BamHⅠ(dC)16引物进行PCR扩增,结果5′接头被克隆到质粒pTZ18上。
1.2.3 合成cDNA cDNA文库合成反应体系:10×逆转录酶缓冲液8.0 μL、MgCl2(25 mmol/L)16 μL、dNTP(10 mmol/L)8.0 μL、AMV逆转录酶(20 U/μL)4.0 μL、RNasin(RNA酶抑制剂,40 U/μL) 2.0 μL、 RT引物(100 μmol/L)4.0 μL、连接有3′接头的总RNA38 μL。首先将反应液与RT引物均匀混合,80 ℃下处理2 min后置于冰中冷却。RT引物序列:5′-GACCACGAGTTAGTTAGCGTACAT-3′。然后参照配制反应体系(Promega公司试剂盒),按照以下程序合成cDNA文库:30 ℃ 10 min→42~55 ℃1 h→95 ℃ 5 min→5 ℃ 5 min。
1.2.4 PCR检测反应体系和反应程序 递减PCR(Touchdown PCR)是在引物的最高退火温度与最适退火温度之间,以一定温度梯度逐步递减至最适退火温度,即通过在PCR的前几个循环使用严谨退火条件以提高扩增产物特异性的方法。例如循环可设在比估算的退火温度高约5 ℃的退火温度下开始,然后每个循环降低1~2 ℃,直到降至退火温度(也可低于退火温度5 ℃)。特异性最高的目的模板会被优先扩增,这些产物在随后的循环中继续扩增并占据优势。
1.2.5 割胶回收、转化连接、测序 DNA纯化后利用琼脂糖凝胶DNA回收方法割胶回收。在微量离心管中配制下列DNA溶液(全量为5 μL):pMD19-T载体1 μL;Control Insert(TaKaRa提供的DNA片段) 1 μL;ddH2O 3 μL。加入5 μL(等量)的 Solution I,16 ℃反应30 min。全量(10 μL)加入至100 μL JM109感受态细胞中,冰中放置30 min。42 ℃加热45 s 后,再在冰中放置1 min。加入890 μL SOC培养基,37 ℃ 振荡培养60 min。在含有X-Gal、IPTG、Amp的L-琼脂平板培养基上培养,形成单菌落。计数白色、蓝色菌落;挑选白色菌落,使用PCR法确认载体中插入片段的大小;将培养液送出测序。
2 结果与分析
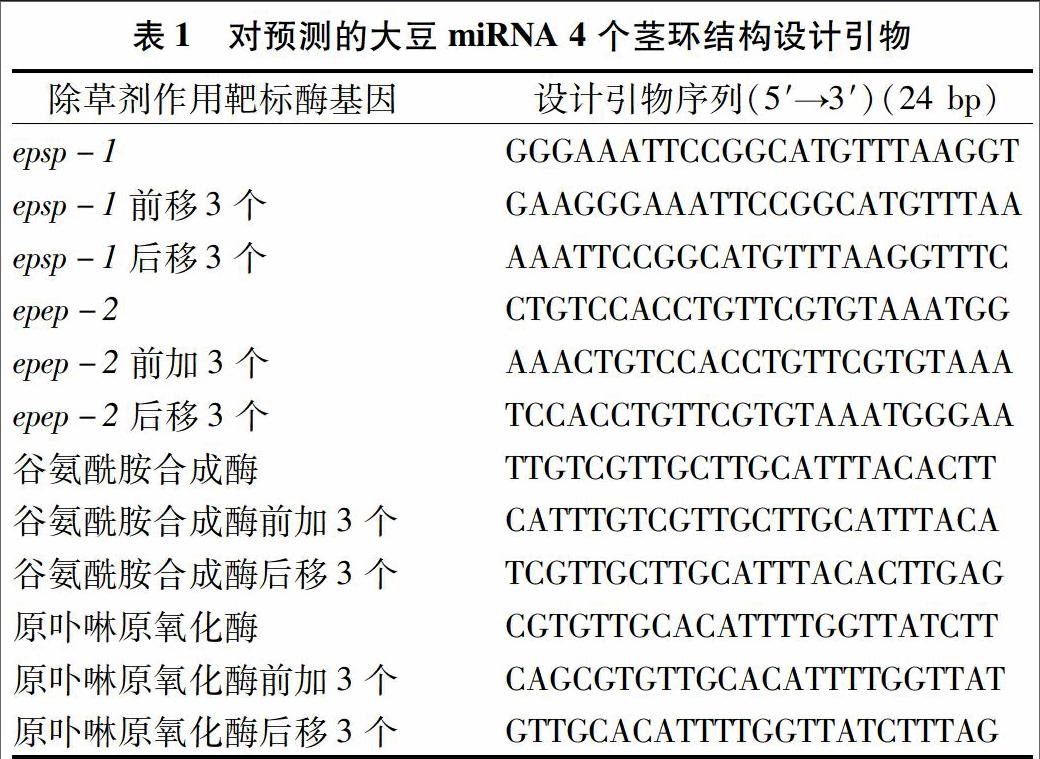
2.1 对已预测的4个miRNA前体序列进行引物设计
确定3′端对预测的茎环结构设计3′端引物。设计原则:在4个茎环上游,根据碱基配对比例设计1条引物,分别前移3个碱基、后移3个碱基,设计3条引物(表1)。
由于microRNA本身序列较短,加接头的目的是与 microRNA 连接便于扩增与识别,3′接头(64 bp)为:5′-PUUUCACTAGCACGAGCTCGCTACGCTCACTACTCGGCATTATGTACGCTAACTAACTCGTGGTCX-3′。
2.2 小RNA的回收、转化连接、PCR检测及测序
对设计的12条引物进行PCR扩增、回收、转化连接和PCR检测。图1显示,每个引物挑5个克隆,目标片段大小约85 bp(3′接头64 bp,小RNA 20 bp左右),每个引物选1管菌液送去测序。
测序结果表明:引物EPSP-2-1在3′端扩出TGG,而引物EPSP-2-2在3′端扩出AA,而且碱基TGG和AA均存在于已经预测的小RNA前体序列中,如果这种3′端多态性确实存在,那么这个茎环结构上的miRNA初步认为是家族式的。
为了进一步验证这种多态性的可靠性,在2个cDNA文库中去扩增:“B Lib.”文库(含豆粒)和“R Lib.”(不含豆粒)文库(图2),每个平板挑10个克隆,进行测序。
测序结果表明:在上述2个文库中,每个文库挑10个克隆,可以看出3′端序列存在多态性,且重复性较高,3′端端点有4种可能(表3)。
测序结果表明:在上述2引物中,每个引物挑5个克隆,可以看出5′端序列存在多态性,5′端序列有5种可能,详见表4。
后续的试验将主要依赖核酸杂交技术。利用试验认定是同一家族的可能6个microRNA组合基因序列作为探针进行Northern Blotting,可以在RACE的基础上进一步验证microRNA是否表达。
3 讨论
本试验从4个靶标酶入手,首先通过软件预测miRNA的二级结构,可知miRNA的二级结构是茎环结构。根据已知miRNA的二级结构碱基配对比例,通过筛选,挑选出可能的miRNA前体,miRNA前体在形成成熟miRNA的过程中,茎环结构中的一条茎被降解,另一条茎形成成熟的miRNA[17]。尽管从理论上来说,成熟的miRNA产生是随机选择的结果,但由于2条链的稳定性有所不同,导致机会的不等[18]。而本研究只是对4个茎环中其中1条茎设计引物,没有对另外的茎设计引物,所以不能排除其他3个茎环结构中也含有成熟的miRNA。本研究确定6个miRNA组合,可能是同一家族。植物miRNA具有簇集现象,多数miRNA不是分散分布的,而是2个或多个基因由1个前体miRNA加工而来,成簇排列的基因协同表达,显现出在基因表达方面的数量优势[19]。
后续试验将主要依赖核酸杂交技术[20]。利用试验认定为同一家族可能的6个microRNA组合基因序列作为探针进行Northern Blotting,可以在RACE的基础上进一步验证microRNA的表达与否。由于microRNA基因的表达比蛋白基因更迅速,且不受翻译过程影响,对目标基因表达的调控效率更高,我们可以把它作为一种强有力的工具,借助它来完成突变体的构建与特定基因表达的调控。虽然对植物microRNA的研究取得了很大进展,但是植物miRNA在转录和转录后水平上其本身如何被调控及其被调控后的结果是怎样的,这些问题都需要人们去研究和探索[21]。随着植物microRNA越来越多地被发现,可以继续研究它们在植物生长发育中的生物学意义和在分子水平上的调节机制,而且还可以有目的、有计划地对它们进行植物生长发育的调控和生理结构的改造,使植物更大限度地表现出对我们有利的性状,从而为人类的生产生活造福。
参考文献:
[1]代俊萍,蔡民华,张飞雄. 植物miRNA研究进展[J]. 首都师范大学学报,2004,25(增刊1):44-46.
[2]贾小云,刘 慧,沈 洁,等. MicroRNA828负调控缺磷胁迫诱导的番茄花青素生物合成[J]. 中国农业科学,2015,48(15):2911-2924.
[3]金龙国,王 川,刘进元. 植物MicroRNA[J]. 中国生物化学与分子生物学报,2006,22(8):609-614.
[4]卢加举,廖文彬,朱白婢,等. MicroRNA的研究进展[J]. 分子植物育种,2006,6(4):73-77.
[5]吴 丹,胡 兰. MicroRNA及其研究进展[J]. 中国畜牧兽医,2006,33(1):40-42.
[6]Barrel D P.MicroRNAs:genomics,biogenesis,mechanism,andfunction[J]. Cell,2004,116(2):281-297.
[7]Bartel B,Bartel D P.MicroRNAs:at the root of plant development[J]. Plant Physiology,2003,132(2):709-717.
[8]Chen C Z.MicroRNAs as oncogenes and tumor suppressors[J]. The New England Journal of Medicine,2005,353(17):1768-1771.
[9]Deo M,Yu J Y,Chung K H.Detection of mammalian microRNAexpression by in situ hybridization with RNA oligonucleotides[J].Developmental Dynamics,2006,235(9):2538-2548.
[10]Herrmann K M,Weaver L M.The shikimate pathway[J]. Annual Review of Plant Physiology and Plant Molceular Biology,1999,50(4):47-50.
[11]Gruys K J,Marzabadi M R,Pansegrau P D,et al.Steady-statekinetic evaluation of the reverse reaction for Escherichia coli5-enolpyruvoylshikimate-3-phosphate synthase[J]. Archives ofBiochemistry and Biophysics,1993,304(2):345-351.
[12]Gruys K J,Walker M C,Sikorski J A.Substrate synergism and the steady-statekinetic reactionmeehanism for EPSP synthase from Escherichia coli[J]. Biochemistry,1992,31(24):5534-5544.
[13]Lauter N,Kampani A,Carlson S,et al.MicroRNA172 down-regulates glossy15 topromote vegetative phase change in maize[J]. Proceedings of the National Academy of Sciences of the United States of America,2005,102(26):9412-9417.
[14]Li S C,Tang P,Lin W C.IntronicmicroRNA:discovery and biological implications[J]. DNA and Cell Biology,2007,26(4):195-207.
[15]Petrocca F,Visone R,Onelli M R,et al.E2F1-regulated micro RNAs impair TGFβ-dependent cell-cycle arrest and apoptosis in gastric cancer[J]. Cancer Cell,2008,13(3):272-286.
[16]Schmid J,Amrhein N.Molecular organization of the shikimate pathway in higher plants[J]. Phytochemistry,1995,39(4):737-749.
[17]Du T,Zamore P D.Microprimer:the biogenesis and function of microRNA[J]. Development,2005,132(21):4645-4652.
[18]Wang J W,Wang L J,Mao Y B,et al.Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis [J]. The Plant Cell,2005,17(8):2204-2216.
[19]Xie Z X,Kasschau K D,Carrington J C.Negative feedback regulation of Dicer-Like1 in Arabidopsis by microRNA-guided mRNAdegradation[J]. Current Biology,2003,13(9):784-789.
[20]肇 莹,肖 军,杨 镇,等. 转导芦苇总DNA耐盐变异水稻SSH文库的构建及部分基因特异表达[J]. 江苏农业科学,2015,43(3):13-16.
[21]Xie Z,Allen E,Fahlgren N,et al.Expression of Arabidopsis miRNA gene[J]. Plant Physiology,2005,138(4):2145-2154.
猜你喜欢
天津医科大学学报(2021年1期)2021-12-05
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
猪业科学(2021年3期)2021-05-21
幽默大师(2020年10期)2020-11-10
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
生物学通报(2018年12期)2018-10-10
猪业科学(2018年4期)2018-05-19
现代检验医学杂志(2016年5期)2016-08-20
茶叶通讯(2014年2期)2014-02-27
