
杏园脐腹小蠹空间分布型及易受害寄主生长性状研究
2016-11-24 07:24朱晓锋徐兵强阿布都克尤木卡德尔张青文
植物保护 2016年5期
朱晓锋, 徐兵强, 阿布都克尤木·卡德尔,宋 博, 张青文, 杨 森*
(1.中国农业大学植物保护学院, 北京 100193; 2. 新疆农业科学院植物保护研究所, 特色林果产业国家地方联合工程研究中心, 农业部西北荒漠作物有害生物综合治理重点实验室, 乌鲁木齐 830091)
杏园脐腹小蠹空间分布型及易受害寄主生长性状研究
朱晓锋1,2, 徐兵强2, 阿布都克尤木·卡德尔2,宋 博2, 张青文1*, 杨 森2*
(1.中国农业大学植物保护学院, 北京 100193; 2. 新疆农业科学院植物保护研究所, 特色林果产业国家地方联合工程研究中心, 农业部西北荒漠作物有害生物综合治理重点实验室, 乌鲁木齐 830091)
在脐腹小蠹(ScolytusschevyrewiSemenov)发生期,通过调查杏园脐腹小蠹空间分布及寄主生长性状发现:在杏园脐腹小蠹呈聚集分布,且个体间相互吸引,分布的基本成分是个体群;当种群密度<1.68 头/株时,环境条件是其聚集的主因;当种群密度>4.92 头/株时,脐腹小蠹的聚集习性和环境条件是脐腹小蠹聚集的主因;其为害程度与杏树新生枝条长度、直径、百叶重和叶片含水量呈显著负相关,与叶绿素含量无相关关系;不同受害程度杏树树脂分泌量呈正态分布,零星被害和持续被害杏树树脂分泌量显著大于健康和易被害致死杏树;6月平均新生枝条长度<14.85 cm、直径<0.31 cm、百叶重<52.40 g的杏树易受害,新生枝条长度<12.70 cm、直径<0.29 cm、百叶重<49.36 g的杏树易受害致死。
脐腹小蠹; 空间分布; 易受害寄主; 生长性状
脐腹小蠹(ScolytusschevyrewiSemenov)属鞘翅目(Coleoptera)小蠹科(Scolytidae),1902年记录于中国新疆天山东部,异名ScolytusfrankeiWichmann、EccoptogasteremarginatusWichmann、E.transcaspicusEggers和ScolytusseulensisMurayama[1]。在中国未将脐腹小蠹(S.schevyrewi)和多毛小蠹(S.seulensis)作一归并[2]。脐腹小蠹传统上分布于亚洲,主要分布于中国北部省区,以及韩国、蒙古、俄罗斯、哈萨克斯坦、乌兹别克斯坦、吉尔吉斯斯坦、土库曼斯坦和塔吉克斯坦等地区[2-5]。脐腹小蠹不但可造成榆树的大量死亡[6-7],而且与多种榆树病原菌共存[7-8],并可传播榆枯萎病[9-10]。2003年脐腹小蠹在美国科罗拉多州和犹他州被首次报道[3],2006年传入加拿大[11],截至2011年已在美国28个州和加拿大的4个省有发生[12]。脐腹小蠹以繁殖能力强和对寄主反应速度快的优势,已在美国的部分州取代当地普遍发生的欧洲榆小蠹[Scolytusmultistriatus(Marsham)],上升为主要害虫[13]。脐腹小蠹寄主广泛,可为害胡颓子科的沙枣(ElaeagnusangustifoliaLinn.)、杨柳科的垂柳(SalixbabylonicaLinn.)、豆科的柠条锦鸡儿(CaraganakorshinskiiKom.)和蔷薇科的多种果树,如杏(ArmeniacavulgarisL.)、桃(AmygdaluspersicaLinn.)、樱桃[Cerasuspseudocerasus(Lindl.) G.Don]、梨(Pyrussp.)、苹果(MaluspumilaMill.)、李(PrunussalicinaLindl.)、扁桃(A.communisLinn.)等,以及榆科的多种榆树,如白榆(UlmuspumilaL.)、春榆(U.propinquaKoidz.)、黑榆(U.davidianaPlanch.)、黄榆(U.macrocarpaHance)、美国榆(U.americanaL.)和欧洲白榆(UlmuslaevisPall.)等[2, 14-15]。在中国脐腹小蠹严重为害榆树、杏树,可造成98%的榆树和52%的杏树死亡[5, 16]。
昆虫种群的空间分布型是种群在栖息地内因生物的和非生物的环境间相互作用,造成种群在一定空间内个体扩散分布的一定形式。研究种群分布型,不仅有助于确定或改进抽样设计方案,而且也可以对研究资料提出适当的数据统计方法,对于了解昆虫的猖獗、扩散行为和种群管理也有一定意义[17]。明确脐腹小蠹空间分布型不但对该虫生态学研究提供依据,而且对该虫的种群控制提供技术资料。在新疆早在1962年就有关于果树小蠹发生为害的报道[18],随后多数科研工作者的研究主要在脐腹小蠹的形态、发生为害规律、天敌资源等方面[14, 16, 19-28],近几年又有关于寄主挥发物对脐腹小蠹引诱的研究报道[29-30]。据前人研究表明,脐腹小蠹主要为害树势较弱的寄主和枝干[16,23,25],易在弱势杏树和有腐烂病、伤口的枝干上定居繁衍[14]。本文在前期研究掌握杏园脐腹小蠹生物学特性和为害规律的基础之上[14],通过棋盘式调查不同种群密度的杏园脐腹小蠹种群空间分布以及易受害杏树外在生长性状,从理论上明确其空间分布型和易受害杏树的生长特征,为杏园脐腹小蠹的防治及研究提供依据。
1 材料与方法
1.1 调查地概况
调查地设在喀什疏勒县塔孜洪乡3村2组,2004年3月15日定植杏园,面积13.5 hm2,品种为‘赛买提杏’(Prunusarmeniaca‘Saimaity’),株行距为3 m×4 m,主要栽培模式为杏园套种冬小麦,夏季直播玉米。
1.2 空间分布型
1.2.1 调查方法
在调查区,于2013年7月中旬在脐腹小蠹成虫发生高峰期选取6块脐腹小蠹为害程度不同的地块,每调查地约1 hm2,按照棋盘式取样法每块地选取25株,统计每株脐腹小蠹成虫数量。
1.2.2 分析方法
采用张孝羲主编的《昆虫生态及预测预报》介绍的聚集指标法、线性回归方程法和聚集均数(λ)概念分析脐腹小蠹空间分布和聚集原因[17]。
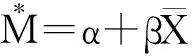

1.3 生长性状
1.3.1 调查方法
2013年6月1日在调查地选取新生枝条长度在0 ~ 50cm未被害的杏树25株,定株测量与观察杏树新生枝条长度、直径、叶绿素含量、百叶重、叶片含水量、韧皮部含水量、树脂分泌量和脐腹小蠹为害情况,杏树被害死亡后停止测量。分析脐腹小蠹为害程度与杏树生长性状之间的关系。测量方法如下。
新生枝条长度、直径和叶绿素含量:将杏树分为东、西、南、北4个方位,每方位分上、下2层,共8个采样点,每采样点随机选取顶端生长旺盛的新生枝条2枝和10片叶,用卷尺、游标卡尺和叶绿素测量计(日本柯尼卡美能达叶绿素计Spad502plus)测量1年新生枝条长度、直径(距基部5cm)以及叶绿素含量。每10d测量1次,共测量7次。

韧皮部含水量:在脐腹小蠹为害期,对定株调查的25株杏树分别取距地1.5 m处被害和未被害杏树主干或主枝10 ~ 20 cm2的韧皮部3块,称取鲜重后,置于75℃的烘箱中,每30 min称重1次,出现恒重时结束测定,并计算含水量,计算方法同上。每20 d测量1次,共测量3次。
树脂分泌量:在脐腹小蠹为害期,对定株调查的25株杏树在主干距地1 m处,使用直径6 mm打孔器打孔至木质部,并保持打孔角度方向一致,打孔24 h后称量树脂分泌量,每30 d测量一次,共测量2次。
1.3.2 分析方法
调查结束后根据每株杏树被脐腹小蠹的为害程度进行分级,并将测量数据分为被害前和被害后进行平均,再利用统计分析软件SPSS 11.5对数据进行被害前后的方差分析、LSD检验和双变量的回归分析。被害程度分级标准如下:
3级:脐腹小蠹虫口数量≥50头/株,整株寄主迅速(约15 d)被害致死,且脐腹小蠹定殖;
2级:脐腹小蠹虫口数量≥50头/株,持续被害,寄主部分枝干被害致死,且脐腹小蠹定殖;
1级:脐腹小蠹虫口数量<50头/株,寄主零星被害,脐腹小蠹未定殖;
0级:脐腹小蠹寄主未受害。
2 结果与分析
2.1 空间分布型
2.1.1 聚集指标测定
2.1.2 线性回归方程检验
2.1.3 聚集原因分析
脐腹小蠹在6块杏园的聚集均数(λ)见表1,在种群密度大于4.92头/株时,聚集均数(λ)分别为6.67、2.25、22.34、2.45,均大于2,说明聚集是由脐腹小蠹聚集习性和环境条件共同引起的,随着为害程度的加剧,聚集均数呈上升趋势;当平均种群密度小于1.68头/株时,聚集均数(λ)分别为0.16和0.76,均小于2,说明聚集是由环境条件引起的。
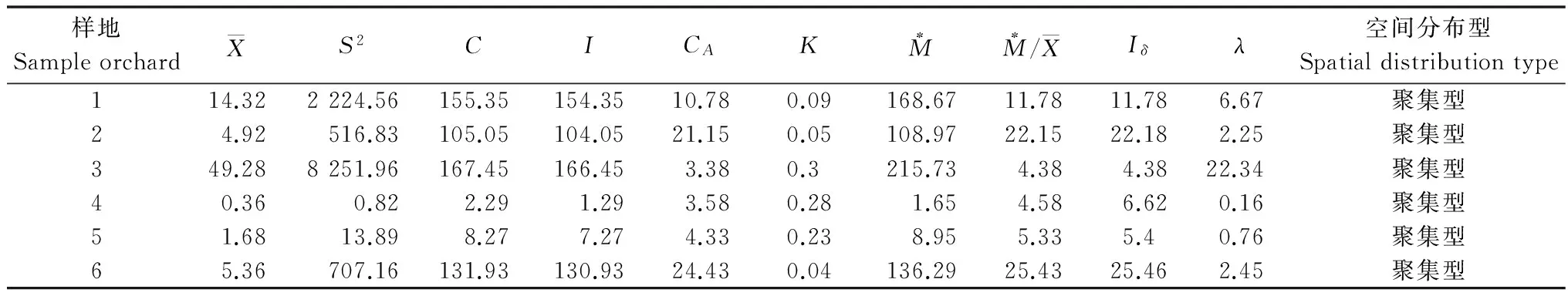
表1 杏园脐腹小蠹成虫聚集指标Table 1 Aggregation indexes of Scolytus schevyrewi in apricot orchard
2.2 脐腹小蠹为害与杏树生长性状的相关性
通过调查脐腹小蠹为害前后杏树外在生长性状,结果表明(见表2):6月杏树被害前平均新生枝条长度、新生枝条直径和百叶重与脐腹小蠹的后期为害程度呈显著或极显著负相关关系,完全被害致死杏树被害前平均新生枝条长度(±SE)为(7.05±4.30)cm、直径为(0.26±0.01)cm,与未被害杏树新生枝条长度(22.99±2.51)cm和直径(0.37±0.02)cm有显著性差异,平均百叶重、叶片含水量、叶绿素含量在不同被害程度杏树间无显著差异;杏树被害后平均新生枝条长度、新生枝条直径、百叶重和叶片含水量与被害程度呈显著或极显著负相关关系,完全被害致死杏树平均新生枝条长度、直径、百叶重和叶片含水量与未被害杏树有显著性差异,随被害程度的加重韧皮部含水量逐渐减少,但无显著性负相关关系,不同被害程度杏树树脂分泌量呈正态分布,未被害和易被害致死杏树树脂分泌量较少,分别为(7.03±5.20)g和(14.00±11.00)g;持续被害和零星被害杏树树脂分泌量较多,分别为(166.67±58.36)g和(167.33±122.47)g,两者之间有显著性差异。
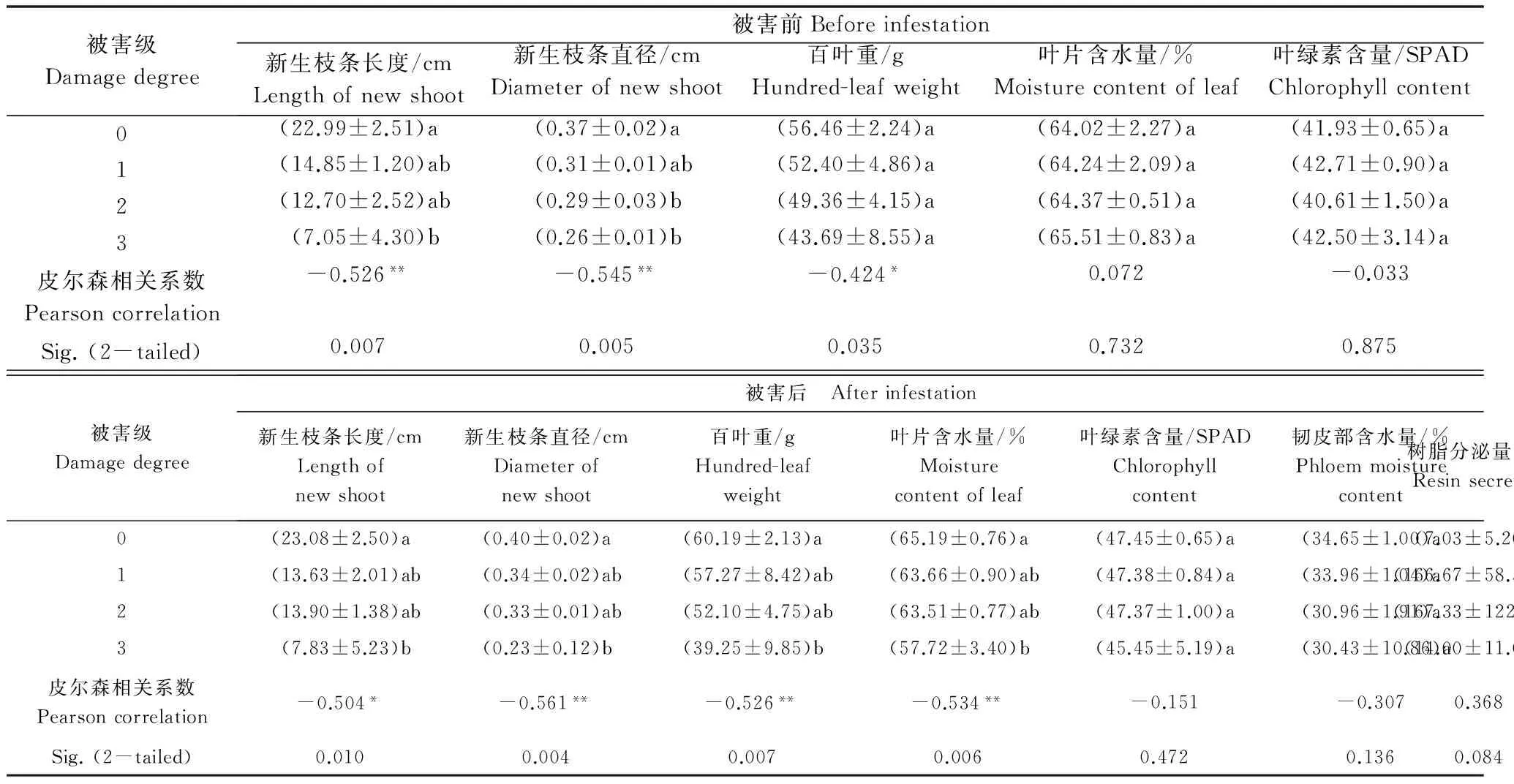
表2 杏树生长性状与脐腹小蠹为害相关性分析1)Table 2 Correlation analysis of the growth characters of apricot and the infestation of Scolytus schevyrewi
1) 利用SPSS 11.5统计软件进行分析,同列标有相同字母表示无显著性差异(LSD,P>0.05);皮尔森相关系数标有*或**的表示有显著性或极显著性相关关系(P<0.05或0.01)。
Means were compared by ANOVA followed by the LSD test (SPSS11.5 for Windows). The data with the same letter are not significantly different in the same column.*or**indicated significant correlation at the 0.05 or 0.01 level (2-tailed).
3 讨论
目前小蠹对寄主的选择理论主要有2种:一是初级引诱,小蠹受到伤害或病变树木挥发的几种化学信息素的引诱,第二是自由选择,小蠹在飞行中随机遭遇到寄主,并进行短期的嗅觉和味觉尝试。最初落于寄主并寻找合适枝干进行钻蛀侵害的个体称之为“先锋个体”,先锋个体若要成功定殖一株活的寄主之上,需要释放足够的信息素来招引更多的同种个体来克服寄主的抗性[31-32]。本研究发现:在杏园不同种群密度脐腹小蠹呈聚集分布,且个体间相互吸引,分布的基本成分是个体群,其聚集程度与种群密度有关;当种群密度<1.68头/株时,环境条件是引起脐腹小蠹聚集的主因;当种群密度>4.92头/株时,脐腹小蠹的聚集习性和环境因素是脐腹小蠹聚集的原因;杏树新生枝条长度、直径和百叶重与脐腹小蠹后期为害程度呈显著负相关。此外,据任龙研究表明杏树挥发物己醛、庚醛、壬醛、α-蒎烯对脐腹小蠹有一定的引诱效果,但引诱效果不明显[30];范丽华通过分析脐腹小蠹成虫后肠和虫粪的挥发物及田间生物测定推断十八烷、十九烷、十二酸和十四酸是脐腹小蠹聚集信息素的主要成分[33]。由此我们推测,在杏园脐腹小蠹受到易受害杏树(特别是新生枝条长度<14.85 cm的杏树)挥发物的引诱或偶然遭遇到易受害杏树,其先锋个体经过初步侵害,判断为合适的寄主便释放聚集信息素,招引更多的同种个体共同进攻易受害杏树。由于部分易受害杏树(新生枝条长度在12.70 ~14.85 cm)具有一定的抵抗力,呈现少量脐腹小蠹虫零星侵害和持续侵害造成部分枝干死亡,而衰弱的杏树(特别是新生枝条长度<12.70 cm)由于树体抗性不强,迅速被大量聚集的脐腹小蠹为害致死。被害后杏树树体24 h树脂分泌量也初步证明了这一点。据束庆龙等研究表明松树泌脂速度与植株的抗病性呈显著的负相关,泌脂速度可作为松树对松材线虫病的抗性指标之一[34]。杏树泌脂速度与植株抗虫性的研究尚未见报道,但据本文研究结果杏树树脂分泌量因脐腹小蠹的为害程度而异,呈正态分布,零星被害和持续被害并造成部分枝干被害致死的杏树树脂分泌量显著大于未受害和易被害致死杏树,因此推测在易受害杏树中新生枝条长度在12.70 ~ 14.85 cm的杏树具有一定的抵抗力,呈现少量小蠹虫零星侵害和持续侵害造成部分枝干致死,而衰弱的杏树(新生枝条长度<12.70 cm),抵抗力较弱,呈现出一旦被脐腹小蠹为害,便迅速死亡。适合脐腹小蠹定殖的寄主在杏园的不均匀分布以及脐腹小蠹释放聚集信息素可能是造成脐腹小蠹聚集分布的主要因素。作者认为通过健康栽培提高杏树生长势是预防脐腹小蠹暴发危害的关键,研究易受害杏树挥发物以及脐腹小蠹聚集信息素对脐腹小蠹的引诱作用,是诱杀防治脐腹小蠹的理论基础。本文已明确脐腹小蠹为害程度与杏树新生枝条长度、直径以及百叶重有显著负相关关系,上述指标可用于杏树脐腹小蠹为害风险评估,而脐腹小蠹的入侵机制和杏树的抗性机制有待进一步研究。
[1] Wood S L, Bright D E.A catalog of Scolytidae and Platypodidae (Coleoptera), part 2: taxonomic index volume A[J]. Great Basin Naturalist Memoirs, 1992(13): 1-833.
[2] 黄复生,李兆麟,殷惠芬. 中国经济昆虫志第二十九册(鞘翅目小蠹科)[M]. 北京: 科学出版社, 1984: 1-205.
[3] Negrón J F, Witcosky J J, Cain R J, et al. The banded elm bark beetle:a new threat to elms in north America [J]. American Entomologist, 2005, 51(2): 84-94.
[4] Douglas H, Bouchard P, Anderson R S, et al. New Curculionoidea (Coleoptera) records for Canadа[J]. ZooKeys, 2013, 309: 13-48.
[5] 张培,吴晶,余本渊,等. 我国榆树害虫发生及研究现状[J]. 江苏林业科技, 2014, 41(1): 46-49.
[6] 李清西,蒋新民,刘芳政. 榆树脐腹小蠹生物学习性初步研究[J]. 八一农学院学报, 1987, 33(3): 12-18.
[7] 徐阳,朱晓梅,刘雪峰. 榆树脐腹小蠹坑道真菌研究[J]. 森林工程, 2013, 29(4): 35-38, 59.
[8] 沙依拉·帕那西,朱晓梅,徐阳,等. 脐腹小蠹伴生菌致病性测定[J]. 安徽农业科学, 2014, 42(10): 2949-2951.
[9] Jacobi W R, Koski R D, Negron J F.Dutch elm disease pathogen transmission by the banded elm bark beetle [J]. Forest Pathology, 2013, 43(3): 232-237.
[10]Jacobi W R, Koski R D, Harrington T C, et al. Association of ophiostoma novo-ulmi withScolytusschevyrewi(Scolytidae) in Colorado [J]. Plant Disease, 2007, 91(3): 245-247.
[11]Humble L M, John E, Smith J, et al. First records of the banded elm bark beetle,ScolytusschevyrewiSemenov (Coleoptera: Curculionidae: Scolytinae), in British Columbia [J]. Journal of the Entomological Society of British, 2010, 107: 21-24.
[12]Lee J C, Negrón J F, Mcelwey S J, et al. Biology of the invasive banded elm bark beetle (Coleoptera: Scolytidae) in the western United States [J]. Annals of the Entomological Society of America, 2011, 104(4): 705-717.
[13]Lee J C, Seybold S J.Host acceptance and larval competition in the banded and European elm bark beetles,ScolytusschevyrewiandS.multistriatus(Coleoptera: Scolytidae): potential mechanisms for competitive displacement between invasive species [J]. Journal of Insect Behavior, 2010, 23(1): 19-34.
[14]李宏,朱晓锋,阿布都克尤木,等. 喀什地区多毛小蠹发生与为害规律[J]. 植物保护, 2009, 35(6): 135-139.
[15]虞国跃,胡亚莉,王合,等. 榆树脐腹小蠹的识别与防治[J]. 植物保护, 2014, 40(6):196-198.
[16]李中焕,托乎提·阿木提,胡卫江. 多毛小蠹虫的发生及综合防治技术[J]. 新疆农业科学, 1993(5): 215-216.
[17]张孝羲. 昆虫生态及预测预报[M]. 第3版. 北京: 中国农业出版社, 2002: 1-323.
[18]杜逾. 喀什市果树小蠹虫初步调查[J]. 新疆农业科学, 1963(8): 313-314.
[19]刘复玳,冯远斌. 多毛小蠹药剂防治试验[J]. 甘肃农大学报, 1987(3): 59-62.
[20]韩峻,乔善元,邓小山,等. 桃树多毛小蠹的发生规律及防治方法的研究[J]. 辽宁农业科学, 1989(5): 42-44.
[21]李江霖,张涛,李新唐,等. 新疆果树多毛小蠹生物学特性及防治[J]. 植物保护, 1995, 21 (1): 8-10.
[22]赵林忠,马俊华,朱明成,等. 杏树多毛小蠹的发生和防治研究[J]. 植保技术与推广, 1995(2): 11-12.
[23]朱承美,曲爱军,路丛山,等. 多毛小蠹虫形态和生物学观察[J]. 落叶果树, 1997(S1): 30-31.
[24]杨虹民,周树星. 桃多毛小蠹虫的发生与防治[J]. 北方果树, 1998(1): 35.
[25]王树杞,敖贤斌,于丽辰,等. 多毛小蠹的生物学特性及防治[J]. 中国果树, 1998(2): 11-13.
[26]张鲁豫,赵莉,李军如,等. 轮台县杏树多毛小蠹生物学特性研究[J]. 新疆农业科学, 2011, 48 (9): 1655-1660.
[27]张鲁豫,赵莉,范毅,等. 轮台县杏树小蠹虫天敌种类及小蠹蒲螨生物学特性研究初探[J]. 新疆农业大学学报, 2011, 34 (6): 507-511.
[28]朱晓锋,崔荣,毕海燕,等. 新疆小蠹虫主要寄生性天敌蒲螨生物学特性初步研究[J]. 新疆农业科学, 2011, 48 (2): 316-319.
[29]隋学良. 多毛小蠹和皱小蠹生物学特性及诱捕技术研究[D]. 北京:北京林业大学, 2011.
[30]任龙. 几种蔷薇科植物挥发物对多毛小蠹虫化学通讯影响的研究[D]. 北京:北京林业大学, 2013.
[31]Byers J A, Zhang Q H.Chemical ecology of bark beetles in regard to search and selection of host Trees[M].Beijing and Springer-Verlag Berlin Heidelberg: Higher Education Press, 2011:150-190.
[32]Byers J A.Host-tree chemistry affecting colonization in bark beetles [C]. New York: Chapman and Hall, 1995: 154-213.
[33]范丽华,牛辉林,张金桐,等. 脐腹小蠹聚集信息素的提取鉴定和引诱效果[J]. 生态学报, 2015, 35(3): 892-899.
[34]束庆龙,汤坚,黄长春,等. 松树泌脂速度对松材线虫病的抗性影响[J]. 安徽农业大学学报, 2006, 33(1): 1-4.
(责任编辑:田 喆)
Spatial distribution type ofScolytusschevyrewiSemenov and the growth characters of the vulnerable host in apricot orchard
Zhu Xiaofeng1,2, Xu Bingqiang2, Abudukeyim Kader2, Song Bo2, Zhang Qingwen1, Yang Sen2
(1. College of Plant Protection, China Agricultural University, Beijing 100193, China; 2. National Local Joint Engineering Research Center of Special Forestry and Fruit Industry; Key Laboratory of Integrated Pest Management on Crops in Northwestern Oasis, Ministry of Agriculture; Institute of Plant Protection, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China)
During the emergence period ofScolytusschevyrewiSemenov, the spatial distribution type ofS.schevyrewiand host growth characters were studied in apricot orchards. The results showed that the spatial distribution type ofS.schevyrewiwas aggregative distribution, with individuals attracting each other, and the individual group was the basic component in its distribution. When the population density<1.68 individuals/plant, the environmental conditions were the main cause for the aggregation; when the population density>4.92 individuals/plant, the environmental factors and aggregation behavior were the main cause. The damage degree ofS.schevyrewihad significantly negative correlation with the length and diameter of new shoots, weight per hundred leaves and leaf water content. However, it had no correlation with chlorophyll content of apricot leaves. Resin secretion of apricot was in a normal distribution. Resin secretion of the vulnerable apricots under sporadic or continued damage was significantly greater than that of healthy and vulnerable dead apricots. The apricots with new shoot length less than 14.85 cm, new shoot diameter less than 0.31 cm, and hundred-leaf weight less than 52.40 g were easy to be infested byS.schevyrewi. The apricots with new shoot length less than 12.70 cm, new shoot diameter less than 0.29 cm, hundred-leaf weight less than 49.36 g were easy to be infested and die.
Scolytusschevyrewi; spatial distribution; vulnerable host; growth character
2015-11-13
2015-12-28
新疆维吾尔自治区公益性科研院所基本科研业务经费(KY2015062); 新疆维吾尔自治区重大专项(201130102-3)
S 436.62
A
10.3969/j.issn.0529-1542.2016.05.013
* 通信作者 E-mail:zhangqingwen@263.net,yangsenxj@126.com
猜你喜欢
散文诗世界(2022年1期)2022-01-13
小品文选刊·印象大同(2021年8期)2021-09-10
教育·校长参考(2020年6期)2020-06-23
学苑创造·A版(2019年10期)2019-11-18
广东第二课堂·初中(2019年4期)2019-05-02
小小说大世界(2018年5期)2018-07-02
读友·少年文学(清雅版)(2018年11期)2018-03-07
家庭医学(2017年9期)2017-10-24
妇女生活(2016年10期)2016-10-12
中国果业信息(2014年4期)2014-01-24
