
全基因组范围禾本科植物GPX基因家族的进化分析和表达模式研究
2016-11-11 02:27:22刘翠晶杨美英
西北农林科技大学学报(自然科学版) 2016年10期
杨 雪,刘翠晶,杨美英
(吉林农业大学 a 生命科学学院,b 中药材学院,吉林 长春 130118)
全基因组范围禾本科植物GPX基因家族的进化分析和表达模式研究
杨雪a,刘翠晶b,杨美英a
(吉林农业大学 a 生命科学学院,b 中药材学院,吉林 长春 130118)
【目的】 研究禾本科植物谷胱甘肽过氧化物酶(GPX)基因家族的序列及其在正常生长和胁迫条件下的表达差异,为揭示GPX基因在植物抵抗逆境胁迫中的作用奠定基础。【方法】 利用生物信息学方法,分析水稻、短柄草、高粱中18个GPX基因的序列特点、基因结构和系统进化关系;采用反转录PCR(RT-PCR)方法,分析18个GPX基因在正常生长以及1-氯-2,4-二硝基苯(CDNB)、双氧水(H2O2)、莠去津(Atrazine)和水杨酸(SA)处理后,水稻、短柄草和高粱的根、茎、成熟叶、幼叶、叶鞘,以及正常生长条件下发芽2 d后幼苗根和芽中的表达情况。【结果】 从水稻、短柄草和高粱基因组中分别鉴定出6,5,7个GPX基因,这些基因编码长度为168~251个氨基酸的蛋白,分子质量介于18.44~27.42 ku。系统发生分析发现,3个物种至少有7个最近的共同祖先GPX基因,物种分化之后水稻和短柄草分别丢失1个和2个GPX基因。序列相似性分析发现,3个物种GPX蛋白具有较高的序列相似性,介于38.0%~95.2%,且不同基因簇中GPX序列的分化速率不同。3个物种GPX基因具有保守的基因结构,但SbGPX7发生了内含子插入事件,预示着其与其他GPX基因之间功能的分化。表达模式分析发现,3个物种的18个GPX基因中有15个在所有检测样品中均为组成型表达,3个GPX基因(水稻2个、高粱1个)为选择性表达,表达模式的分化预示着功能的分化。【结论】 作为植物保护细胞免受氧化损伤的重要酶类,禾本科3个物种的GPX基因在进化过程中发生了功能分化。
谷胱甘肽过氧化物酶;禾本科;基因进化分析;基因表达模式
植物体在光合作用和呼吸作用过程中会产生活性氧,生物和非生物胁迫也会促进植物体产生活性氧,活性氧的不断积累对生物体是有毒性的,大量的活性氧能够破坏蛋白、脂质和核酸等生物大分子[1-2]。在长期的进化过程中,植物已经发展出有效的酶和非酶机制来保护细胞免受氧化损伤,其中清除活性氧的酶类主要包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)和谷胱甘肽过氧化物酶(GPX)等[2]。GPX是植物用来清除活性氧的重要酶类,它通过催化过氧化氢、有机氢过氧化物和脂质过氧化物的还原来保护细胞免受氧化损伤[3]。前人的研究还发现,GPX通过在气孔关闭过程中作为传感器介导脱落酸和过氧化氢信号传导来调节植物的蒸腾作用,在植物响应干旱胁迫过程中发挥着重要作用[4]。
在植物中,对GPX的研究主要集中于双子叶植物上,如拟南芥、菠菜、番茄和烟草等[4-8],对单子叶植物GPX的研究较少。禾本科是单子叶植物中的重要科,其中很多物种是重要的粮食作物,如小麦、玉米、高粱和水稻;有些是重要的牧草,如鸭茅、黑麦草、苏丹草和短柄草等,但干旱、低温等逆境胁迫限制了它们的高产、稳产。本研究以重要的粮食作物水稻(OryzasativaL.ssp.japonica)、优良牧草短柄草(Brachypodiumdistachyon)及重要粮食作物和能源植物高粱(Sorghumbicolor)为研究材料,通过在水稻、短柄草和高粱全基因组[9-11]中搜索其GPX基因,对该基因家族的系统发生关系、序列特征、基因结构及其在不同胁迫处理条件下的表达模式进行详细分析,以期为深入揭示GPX基因在禾本科植物抵抗逆境胁迫中的作用奠定基础。
1 材料与方法
1.1材料
总 RNA 提取试剂盒Aurum Total RNA Kit购自Bio-Rad公司,反转录试剂盒RNA PCR Kit (AMV) version 3.0、TaqDNA聚合酶和其他PCR试剂购自TaKaRa公司,双氧水(H2O2)、1-氯-2,4-二硝基苯(1-chloro-2,4-dinitrobenzene,CDNB)、水杨酸(salicylic acid,SA)购自Sigma公司,除草剂莠去津(Atrazine)购自山东滨农科技有限公司。
水稻、短柄草和高粱均采用基因组测序种,分别为OryzasativaL.ssp.japonica、Brachypodiumdistachyon和Sorghumbicolor。
1.2水稻、短柄草和高粱GPX基因序列的鉴定
以拟南芥的8个GPX蛋白序列为模板,利用TBLASTN检索Phytozome v9.0 (http://www.phytozome.net/) 中水稻、短柄草和高粱的全基因组数据库,获得与拟南芥8个GPX序列均具有较高相似性的序列。通过搜索3个物种的EST库检验获得自动注释基因的剪接方式,对于无对应EST的基因,通过与有EST基因的序列比对检验基因的剪接方式。确认拼接方式正确的基因序列,在http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi上进行CDD 保守结构域分析,分析其编码蛋白是否具有保守的GPX结构域。
1.3系统发生关系和序列相似性分析
根据CDD预测的GPX结构域,截取3个物种GPX基因中对应GPX结构域的核苷酸序列,在Bioedit中翻译为氨基酸序列,然后进行氨基酸序列比对,并进行手动调节,获得氨基酸序列的比对结果。将氨基酸序列反转为核苷酸序列,利用MEGA 4.0中的neighbour-joining(NJ) 法以Kimura 2-parameter模型构建进化树,Bootstrap值为1 000,外类群为EcGPX(Locus tag:UTI89_C1903)。将对比后的GPX蛋白序列在Alignment中生成序列相似性矩阵,获得任意两个基因所编码蛋白的序列相似性。
1.4水稻、短柄草和高粱GPX基因的表达模式分析
为研究GPX基因在水稻、短柄草和高粱不同时期、不同部位中的表达特点,以及对不同胁迫的应答反应,利用基因特异性RT-PCR引物扩增目的片段。将测序种水稻和高粱种植在10 cm×10 cm的方盆中,30 ℃、光照14 h/黑暗10 h条件下,分别培养2个月和1个月,生长至幼苗期;测序种短柄草种植在10 cm×10 cm的方盆中,23 ℃、光照14 h/黑暗10 h条件下培养1个月,生长至幼苗期。对处于同一时期的水稻、高粱和短柄草植株,分别进行1.0 mmol/L CDNB、体积分数5.0% H2O2、体积分数1.5% Atrazine和1.0 mmol/L SA浇灌根部、喷洒地上部处理,每种处理3个重复。经CDNB、H2O2和Atrazine处理12 h以及SA处理24 h后,提取3种植物根、茎、成熟叶、幼叶和叶鞘总RNA;同时提取同一时期正常生长的3种植物根、茎、成熟叶、幼叶和叶鞘总RNA,以及发芽2 d后幼苗根和芽总RNA。以总RNA为模板反转录合成cDNA。序列比对后根据基因序列分别设计特异性引物 (表1),进行RT-PCR扩增。在进行PCR扩增时,模板cDNA上样量为1 μL,以各物种Actin基因作为内标。PCR反应程序为:94 ℃预变性3 min;94 ℃ 30 s,60 ℃ 40 s,72 ℃ 1 min,35个循环;最后72 ℃延伸3 min。PCR结束后进行10 g/L琼脂糖凝胶电泳检测,电泳上样量2 μL。为了进一步验证所扩增到的是否为目的基因,对PCR产物进行测序。
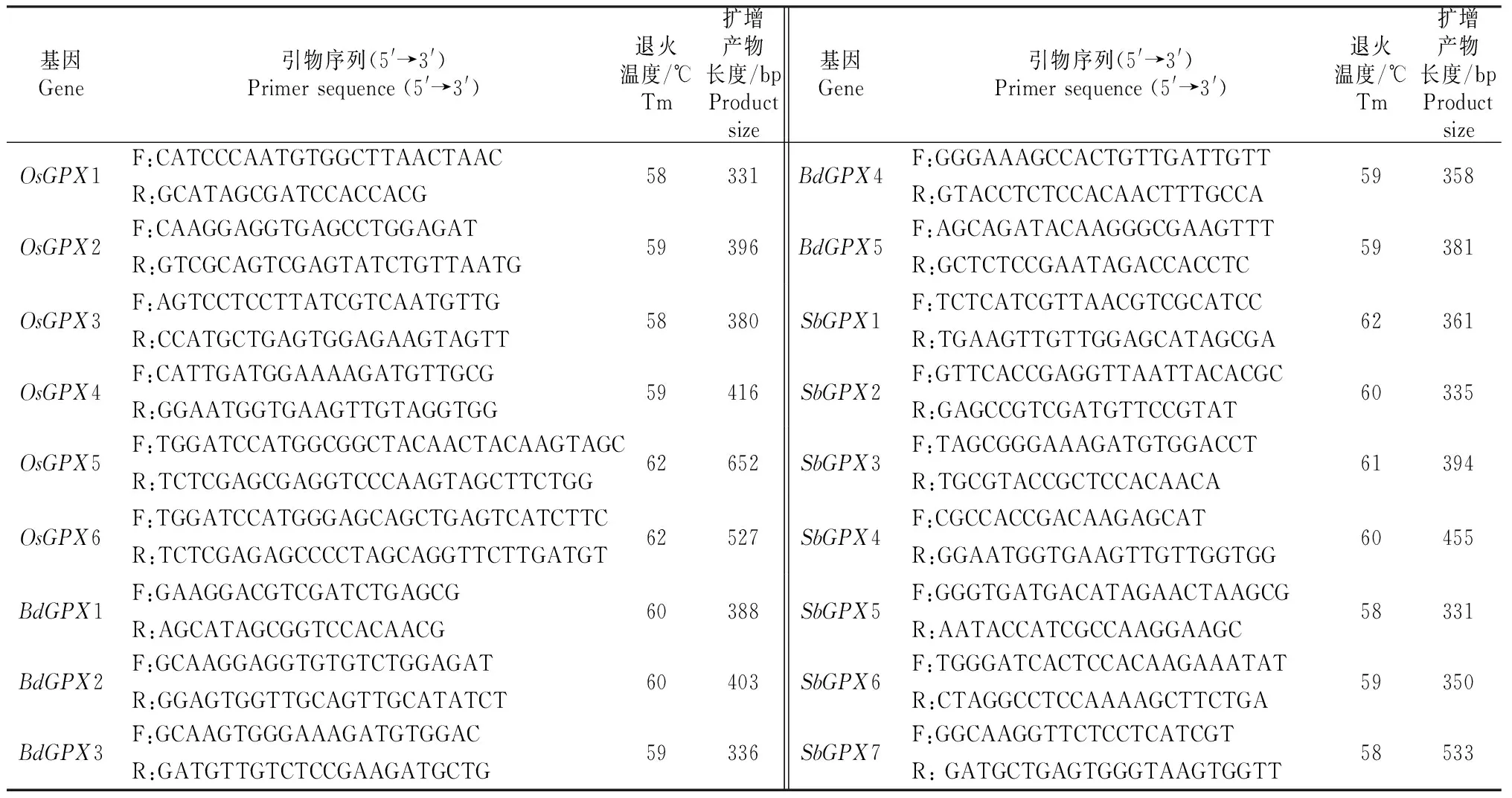
表 1 检测水稻、短柄草和高粱GPX基因表达所用的引物Table 1 Primers used to detect expression of GPX genes in O.sativa,B.distachyon and S.bicolor
注:Os代表水稻,Bd代表短柄草,Sb代表高粱。下表同。
Note:Os,BdandSbrepresentO.sativa,B.distachyonandS.bicolor,respectively.The same below.
2 结果与分析
2.1水稻、短柄草和高粱GPX基因的序列挖掘
根据已知的8个拟南芥GPX蛋白序列,利用TBLASTN检索Phytozome v9.0中水稻、短柄草和高粱的全基因组数据库,分别从各物种全基因组中获得6,5,7个与拟南芥GPX序列均具有较高相似性的蛋白序列。获得序列经过CDD保守结构域分析,确定其编码蛋白具有保守的GPX结构域。
水稻的6个GPX基因分布于5条染色体上,编码长度为168~238个氨基酸的蛋白,蛋白分子质量介于18.48~25.84 ku;短柄草的5个GPX基因分布于3条染色体上,编码长度为168~240个氨基酸的蛋白,蛋白分子质量介于18.48~25.90 ku;高粱的7个GPX基因分布于4条染色体及1条未能组装到染色体的片段(scaffold10)上,编码长度为168~251个氨基酸的蛋白,蛋白分子质量介于18.44~27.42 ku(表2)。
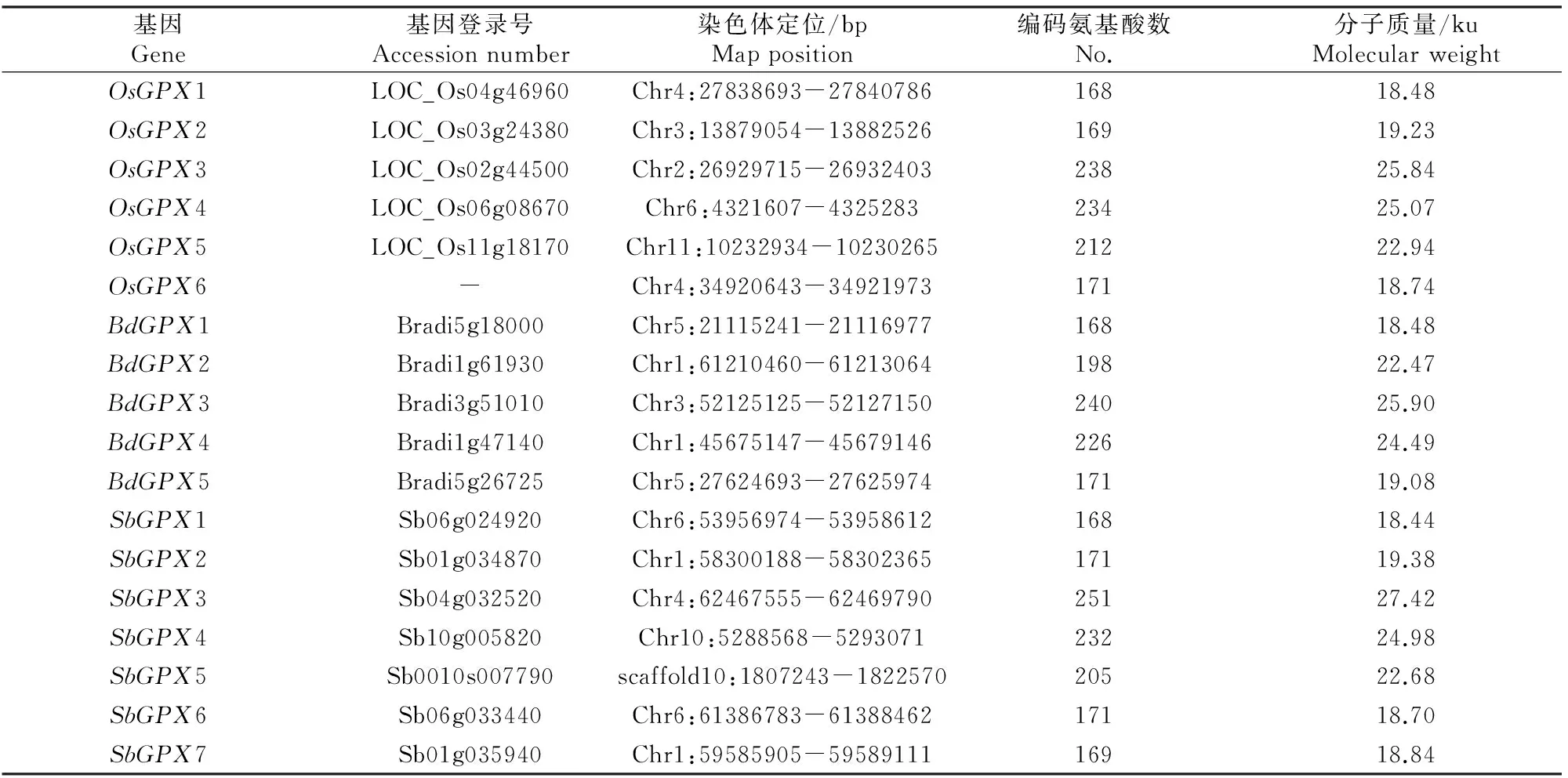
表 2 水稻、短柄草和高粱基因组中鉴定出的GPX基因Table 2 GPX genes identified from O.sativa,B.distachyon and S.bicolor genome
注:-表示基因组未标注。
Note:- represents the gene was not annotation in genome.
2.2水稻、短柄草和高粱GPX基因的命名及系统发生关系分析
根据Margis等[2]的方法将水稻6个GPX基因分别命名为OsGPX1/2/3/4/5/6,以大肠杆菌GPX为外类群,对水稻、短柄草和高粱的GPX基因进行系统发生分析,根据与水稻GPX基因的同源关系,将短柄草的5个GPX基因分别命名为BdGPX1/2/3/4/5,高粱的7个GPX基因分别命名为SbGPX1/2/3/4/5/6/7。
根据3个物种的系统发生树,鉴定出代表水稻、短柄草和高粱最近共同祖先GPX基因的节点,这些节点都具有高的支持率(不小于87%),如图1中圆点所示,并且推断这些节点代表了3个物种分化的节点,每个节点对应一个水稻、短柄草和高粱的直系同源基因簇。例如,在a1基因簇中,包含OsGPX3、BdGPX3和SbGPX3基因,推测在3个物种分化前的祖先物种中存在1个这一簇所对应的最近的共同祖先基因。基因组中也会发生基因丢失,因此某些祖先基因在祖先物种中存在,后来在物种分化后的某一物种中丢失,例如,在a5基因簇中,包含OsGPX5和SbGPX5基因,推测短柄草丢失了对应的基因。根据鉴定出的直系同源基因簇数目,推测这3个物种的祖先物种中至少存在7个GPX基因,而在3个物种分化之后,水稻、短柄草和高粱中都没有发生GPX基因家族的扩张,且水稻丢失了a2基因簇对应的基因,短柄草丢失了a2和a5基因簇对应的2个基因。
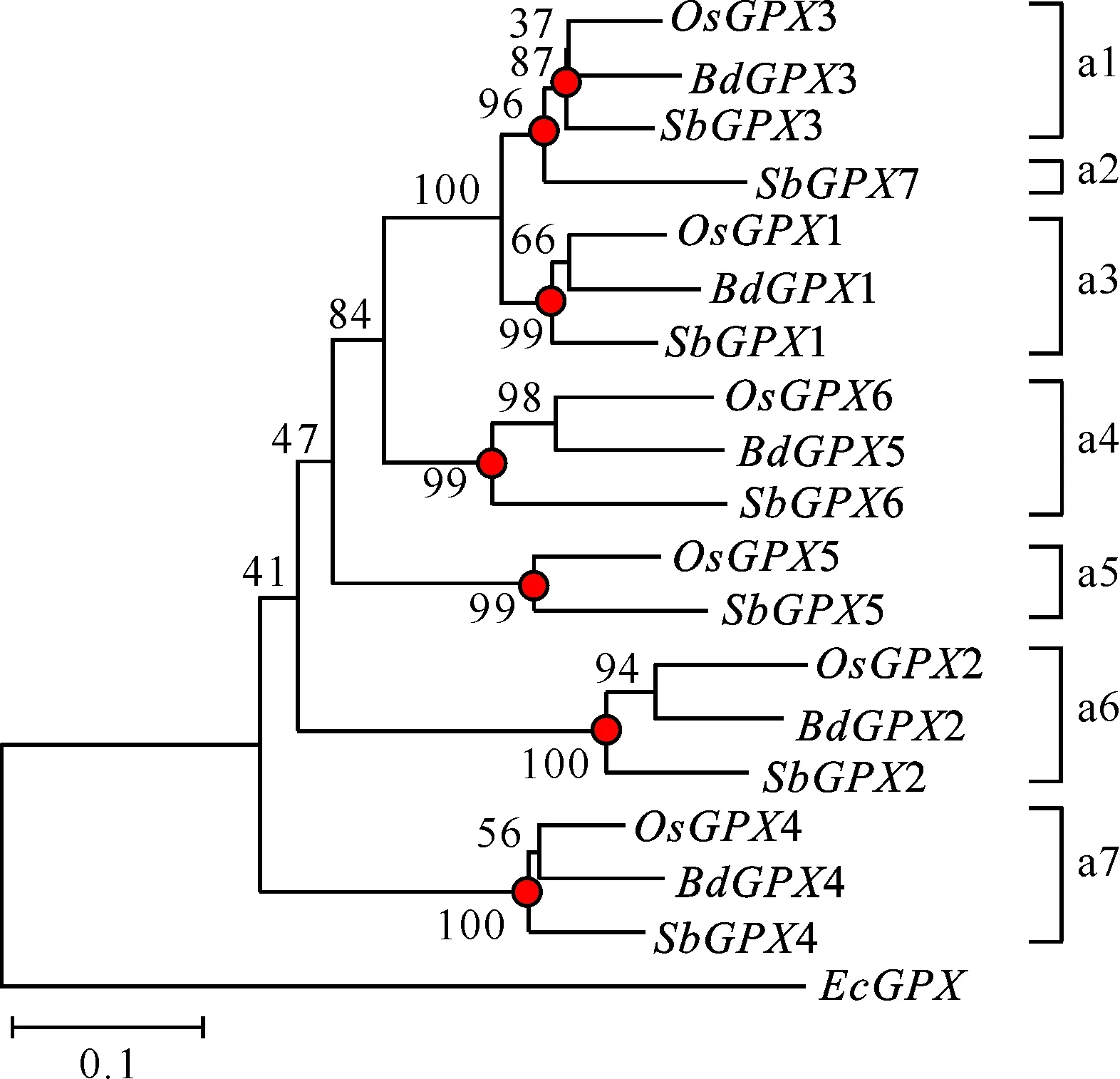
图 1水稻、短柄草和高粱GPX基因的系统发生树枝上的数字代表经过1 000次重复算出的支持率(%);圆点代表3个物种分化前最近的共同祖先基因
Fig.1Phylogenetic tree ofO.sativa,B.distachyonandS.bicolorGPXNumbers on branches indicate the bootstrap percentages calculated from 1 000 replicates.The nodes that represent the most recent common ancestral genes before theO.sativa,B.distachyonandS.bicolorsplit are indicated by black circles
2.3水稻、短柄草和高粱GPX蛋白序列的分析
分别分析3个物种中GPX蛋白的序列相似性,结果见表3。由表3可以看出,在水稻中,OsGPX2与OsGPX4之间的序列相似性最低,只有42.3%;OsGPX1与OsGPX5/6之间的序列相似性最高,为73.2%。在短柄草中,BdGPX2与BdGPX3之间的序列相似性最低,只有38.0%;BdGPX1与BdGPX5之间的序列相似性最高,为71.5%。在高粱中,SbGPX2与SbGPX3之间的序列相似性最低,只有43.6%;SbGPX1与SbGPX7之间的序列相似性最高,为80.4%。在3个物种的所有GPX中,BdGPX2与BdGPX3之间的序列相似性最低,为38.0%;OsGPX1与SbGPX1之间的序列相似性最高,为95.2%。
进一步分析3个物种中直系同源基因(图1中同一簇即为直系同源基因)的蛋白序列相似性(表3)可见,BdGPX3与SbGPX3的序列相似性最低,为65.7%;OsGPX1与SbGPX1的序列相似性最高,为95.2%。
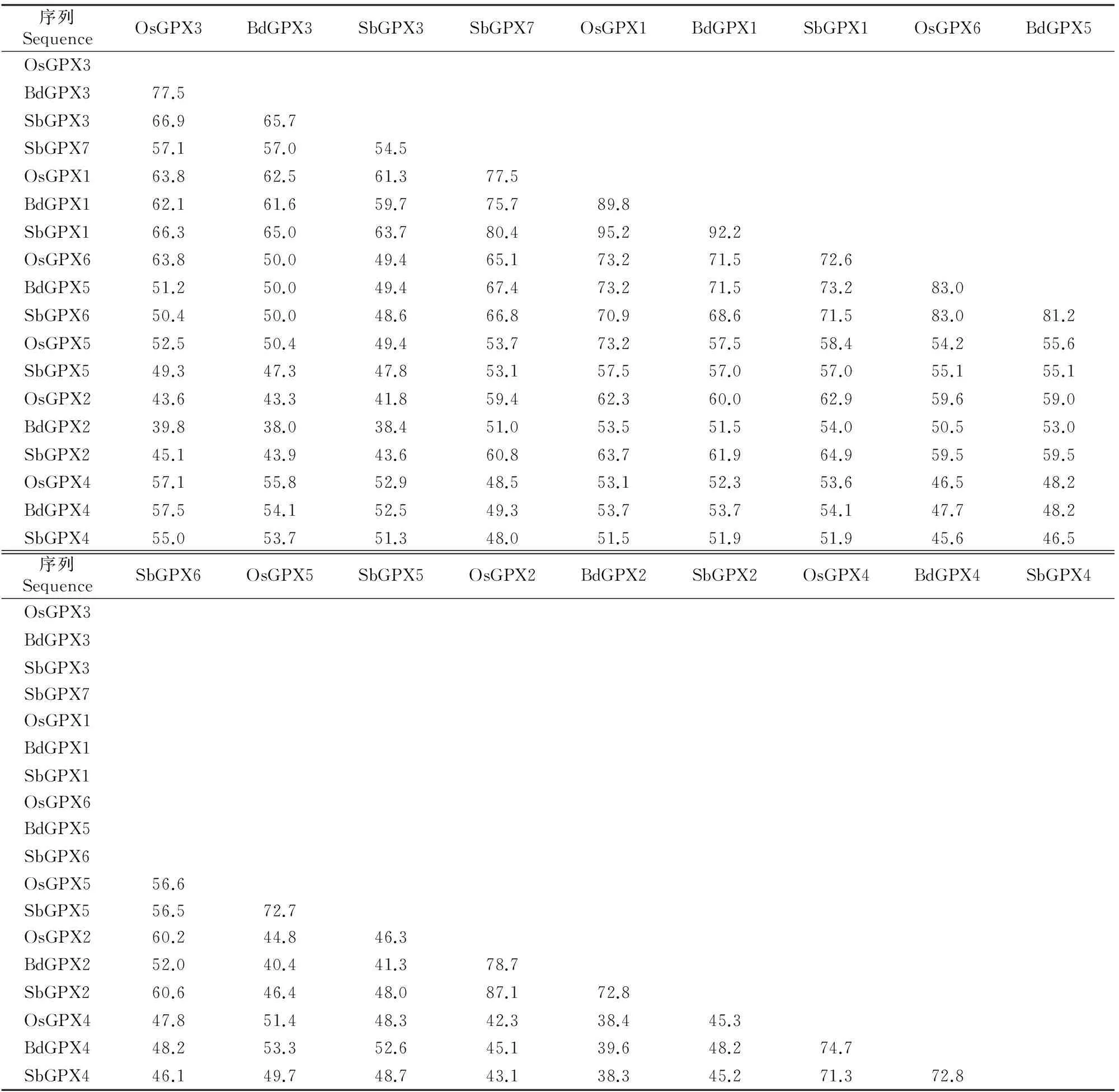
表 3 水稻、短柄草和高粱GPX蛋白的序列相似性分析Table 3 GPX protein sequences identity of O.sativa,B.distachyon and S.bicolor
通过对3个物种GPX蛋白的序列比对发现,在3个物种的所有GPX蛋白中,均存在3个保守的半胱氨酸(如图2箭头所示),且在GPX结构域区,GPX蛋白序列高度保守,序列相似性介于59.8%~97.4%,而在非结构域区,序列差异则较大。

图 2水稻、短柄草和高粱GPX蛋白序列的比对分析
在所有GPX蛋白中均保守的氨基酸残基用黑色背景标示;GPX蛋白的3个保守半胱氨酸残基用黑色箭头指示
Fig.2Sequence alignment of GPX proteins ofO.sativa,B.distachyonandS.bicolorConserved residues in all GPX proteins are marked in black.The three conserved cysteine residues of plant GPX proteins are marked with black arrows
2.4水稻、短柄草和高粱GPX基因的结构分析
分析3个物种GPX基因的结构发现,GPX基因的内含子数目非常保守,除SbGPX7有6个内含子外,其他所有GPX都只有5个内含子,而且所有GPX基因在结构域区的内含子数目和外显子长度都高度保守,除SbGPX2第5个外显子比其他基因对应外显子的蛋白序列多1个氨基酸外,其他相对应的外显子长度都完全保守;对应内含子的长度差异则比较大,例如,OsGPX5第3个内含子长度为138 bp,而SbGPX5第3个内含子长度为5 499 bp。在GPX蛋白N端非GPX结构域区,外显子的长度差别较大,例如OsGPX3第1个外显子的非结构域部分长度为234 bp,而OsGPX1第1个外显子非结构域部分长度只有24 bp。在GPX蛋白C端非GPX结构域区,外显子的长度则比较相近,除BdGPX2为90 bp外,其他都在6~16 bp(图3)。
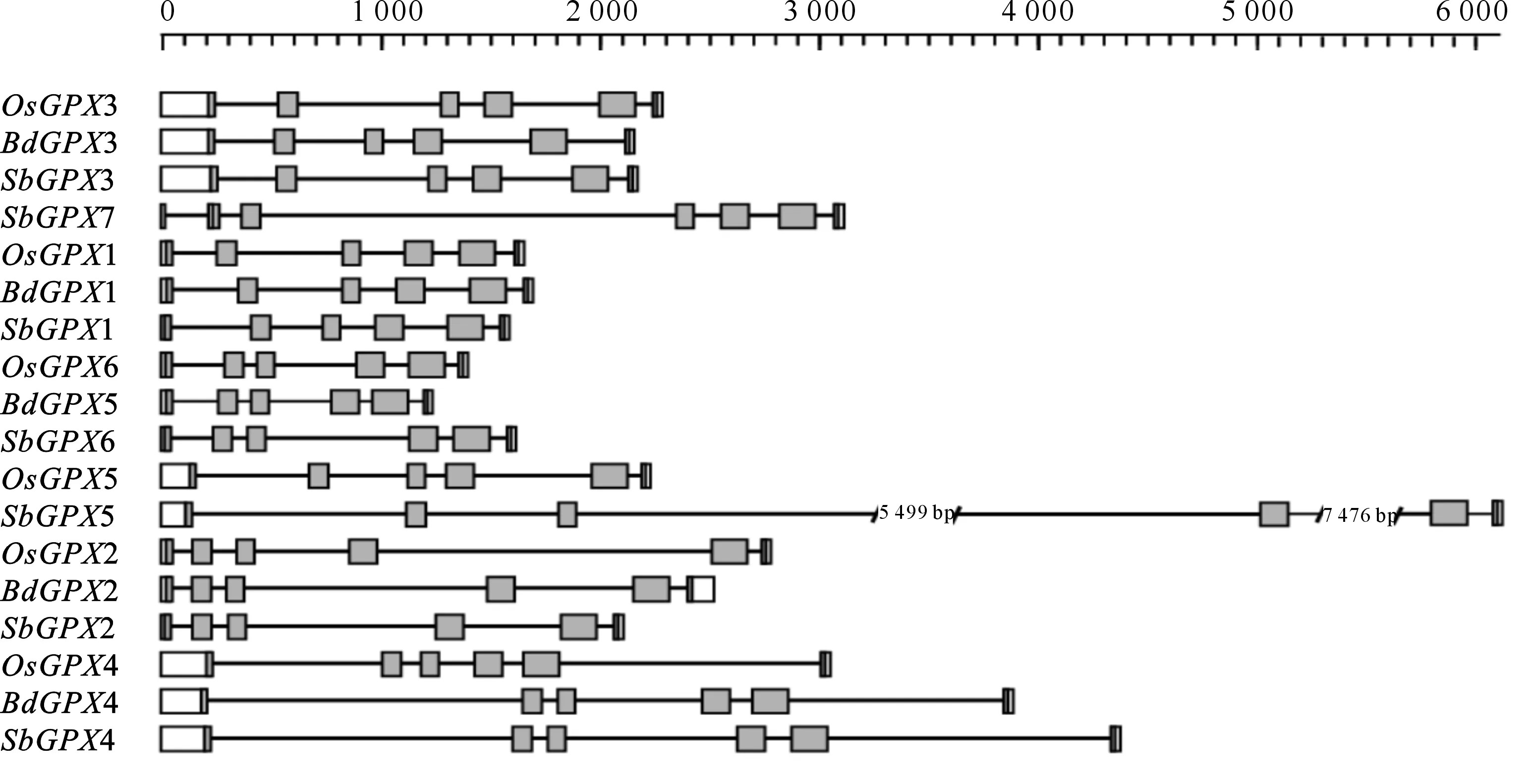
图 3水稻、短柄草和高粱GPX的基因结构灰色方框代表外显子,横线代表内含子
Fig.3Structures ofGPXgenes ofO.sativa,B.distachyonandS.bicolorExons are indicated by gray boxes,while introns are indicated by lines
2.5水稻、短柄草和高粱GPX基因的表达模式
通过反转录PCR(RT-PCR),分析在正常生长和胁迫处理条件下,水稻、短柄草和高粱GPX基因在根、茎、成熟叶、幼叶、叶鞘以及未经胁迫处理的幼苗根、芽中的表达模式,结果见图4。
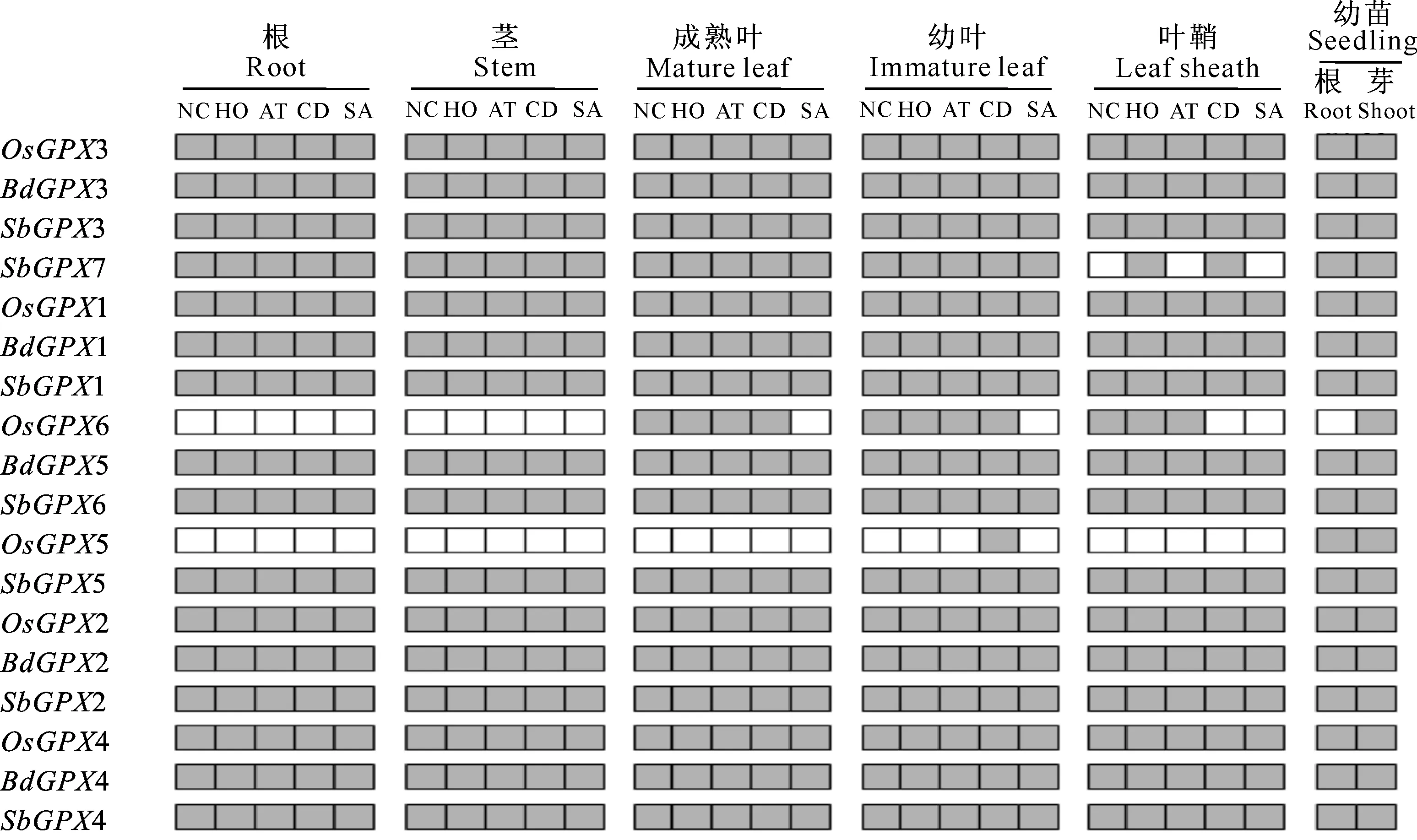
图 4水稻、短柄草和高粱GPX基因的表达模式
灰格表示检测到基因表达,白格表示未检测到基因表达。NC表示对照组(正常生长未经胁迫处理),HO表示H2O2处理,AT表示莠去津处理,CD表示CDNB处理,SA表示水杨酸处理
Fig.4Expression patterns ofGPXgenes ofO.sativa,B.distachyonandS.bicolor
The gray box indicates positive detection of gene expression in the corresponding tissue under normal growth conditions (NC),CDNB (CD),H2O2(HO),atrazine (AT) and salicylic acid (SA) treatments
图4显示,短柄草5个GPX基因在检测的所有样品中都表达;水稻6个GPX基因中,有4个 (OsGPX1/2/3/4) 在检测的所有样品中都表达,有2个 (OsGPX5/6) 是选择性表达,其中OsGPX5只在幼苗根和芽以及CDNB处理下的幼叶中表达,OsGPX6在植株的根和茎、幼苗根、SA处理的成熟叶和幼叶,以及CDNB和SA处理条件下的叶鞘中均不表达;高粱7个GPX基因中,有6个 (SbGPX1/2/3/4/5/6) 在检测的所有样品中都表达,而SbGPX7为选择性表达,在正常生长、Atrazine和SA处理条件下的叶鞘中不表达。
由图4还可见,在3个物种中,直系同源基因的表达模式也发生了分化,例如BdGPX5、SbGPX6和OsGPX6是直系同源基因(图1中a4簇),BdGPX5和SbGPX6在检测的所有样品中均表达,而OsGPX6选择性表达;OsGPX5和SbGPX5是直系同源基因(图1中a5簇),SbGPX5在检测的所有样品中均表达,而OsGPX5则表现为选择性表达。
3 讨 论
GPX是植物体内保护细胞免受氧化损伤的重要酶类[2]。水稻是重要的粮食作物,短柄草是优良的牧草,高粱是重要的粮食作物和能源植物,但在农业生产中由于干旱和低温等逆境胁迫限制了其高产、稳产,因此在这3个物种中研究与抗性相关的GPX基因具有重要的理论意义与应用价值。
本研究从水稻、短柄草和高粱基因组中分别搜索到6,5,7个GPX基因,通过系统发生分析推测3个物种至少有7个最近的共同祖先GPX基因,在3个物种分化以后,水稻和短柄草中分别丢失了1个和2个GPX基因。基因重复和丢失是基因功能变化的有力来源,根据Wapinski等[12]的研究,基因重复和丢失的可能性与基因功能密切相关,且与胁迫相关的基因更容易出现重复和丢失,而与生长发育相关的基因数目更加稳定。GPX基因是抵抗氧化胁迫的基因,其基因数目的变化预示了进化过程中GPX基因在3个物种中功能的变化。
本研究对3个物种直系同源基因蛋白序列的相似性进行分析发现,不同基因簇的序列相似性不同,例如,OsGPX3/BdGPX3、OsGPX3/SbGPX3和BdGPX3/SbGPX3的序列相似性分别为77.5%,66.9%和65.7%,而OsGPX1/BdGPX1、OsGPX1/SbGPX1和BdGPX1/SbGPX1的序列相似性分别为89.8%,95.2%和92.2%。不同基因簇序列相似性的不同预示功能分化速率的差异,序列相似性更高的基因簇中的基因在3个物种中的功能可能更加保守,而序列相似性低的基因簇中的基因在3个物种中的功能可能已经出现了较大的分化[13]。
通过序列比对,本研究在GPX蛋白序列中鉴定出3个保守的半胱氨酸(Cys),根据Navrot等[1]的研究结果,第122和170位的Cys是GPX的催化位点,在催化过程中,首先是第122位Cys残基与过氧化物反应,形成Cys-SOH,然后Cys-SOH与第170位Cys残基形成一个分子内二硫键,再通过与Trx反应将二硫键还原,使GPX回到还原态,当2个Cys中的任意一个突变之后,GPX蛋白都不能持续发挥催化功能;而第151位Cys残基可影响GPX蛋白对过氧化物底物的结合能力,当这个位点的Cys残基突变以后,GPX对过氧化底物的结合能力将明显降低。
本研究发现,水稻、短柄草和高粱GPX基因的结构非常保守,除SbGPX7有6个内含子外,其他所有GPX都只含有5个内含子,而且所有GPX基因结构域区的内含子数目和外显子长度都高度保守。SbGPX7多出的1个内含子可能是内含子插入的结果,从而引起SbGPX7与其他GPX基因结构的分化。基因结构的分化对重复基因分化具有重要作用[14],SbGPX7与其他GPX基因的结构分化预示其功能的分化。
对3个物种GPX基因的表达模式研究发现,水稻、短柄草和高粱分别有4,5,6个GPX基因在所有检测的样品中均表达,说明这些基因可能是组成型表达基因,预示这些基因在正常条件及胁迫条件下的所有部位中均发挥作用。而水稻OsGPX5、OsGPX6和高粱SbGPX7为选择性表达基因,可能只在某些条件下或某些发育阶段的某些部位发挥作用。表达的时空差异预示着水稻的2个和高粱的1个基因功能发生了分化。
[1]Navrot N,Collin V,Gualberto J,et al.Plant glutathione peroxidases are functional peroxiredoxins distributed in several subcellular compartments and regulated during biotic and abiotic stresses [J].Plant Physiology,2006,142(4):1364-1379.
[2]Margis R,Dunand C,Teixeira F K,et al.Glutathione peroxidase family:an evolutionary overview [J].FEBS Journal,2008,275(15):3959-3970.
[3]Milla M A R,Maurer A,Huete A R,et al.Glutathione peroxidase genes inArabidopsisare ubiquitous and regulated by abiotic stresses through diverse signaling pathways [J].The Plant Journal,2003,36(5):602-615.
[4]Miao Y,Lü D,Wang P,et al.AnArabidopsisglutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses [J].The Plant Cell Online,2006,18(10):2749-2766.[5]Chang C C C,Slesak I,Jordá L,et al.Arabidopsischloroplastic glutathione peroxidases play a role in cross talk between photooxidative stress and immune responses [J].Plant Physiology,2009,150(2):670-683.
[6]Sugimoto M,Furui S,Suzuki Y.Molecular cloning and characterization of a cDNA encoding putative phospholipid hydroperoxide glutathione peroxidase from spinach [J].Biosci Biotechnol Biochem,1997,61(8):1379-1381.
[7]Chen S,Vaghchhipawala Z,Li W,et al.Tomato phospholipid hydroperoxide glutathione peroxidase inhibits cell death induced by bax and oxidative stresses in yeast and plants [J].Plant Physiology,2004,135(3):1630-1641.
[8]Criqui M C,Jamet E,Parmentier Y,et al.Isolation and characterization of a plant cDNA showing homology to animal glutathione peroxidases [J].Plant Mol Biol,1992,18(3):623-627.
[9]Goff S A,Ricke D,Lan T H,et al.A draft sequence of the rice genome (OryzasativaL.ssp.japonica) [J].Science,2002,296(5565):92-100.
[10]Vogel J P,Garvin D F,Mockler T C,et al.Genome sequencing and analysis of the model grassBrachypodiumdistachyon[J].Nature,2010,463(7282):763-768.
[11]Paterson A H,Bowers J E,Bruggmann R,et al.TheSorghumbicolorgenome and the diversification of grasses [J].Nature,2009,457(7229):551-556.
[12]Wapinski I,Pfeffer A,Friedman N,et al.Natural history and evolutionary principles of gene duplication in fungi [J].Nature,2007,449(7158):54-61.
[13]Duan Z H,Hughes B,Reichel L,et al.The relationship between protein sequences and their gene ontology functions [J].BMC Bioinformatics,2006,7(1):1-11.
[14]Xu G,Guo C,Shan H,et al.Divergence of duplicate genes in exon-intron structure [J].Proceedings of the National Academy of Sciences,2012,109(4):1187-1192.
Genome-wide evolutionary analysis and expression of glutathione peroxidase genes in Gramineae
YANG Xuea,LIU Cuijingb,YANG Meiyinga
(aCollegeofLifeSciences,bCollegeofChineseMedicinalMaterials,JilinAgriculturalUniversity,Changchun,Jilin130118,China)
【Objective】 The sequences and expression patterns of glutathione peroxidase (GPX) genes in Gramineae were investigated to provide information for studying the role ofGPXin stress tolerance.【Method】 Sequence characteristics,gene structures and phylogenetic relationship of 18GPXsfromOryzasativa,BrachypodiumdistachyonandSorghumbicolorwere analyzed by bio-informatics tools.Gene expression patterns were analyzed by reverse transcription PCR (RT-PCR) in roots,stem,leaves,and leaf sheath exposed to various treatments (1-chloro-2,4-dinitrobenzene (CDNB),H2O2,atrazine and salicylic acid (SA)) for 12 or 24 h and in roots and shoots of seedlings 2 d after germination under normal condition.【Result】 The numbers ofGPXgenes identified fromO.sativa,B.distachyonandS.bicolorgenomes were 6,5 and 7,respectively.These genes encoded proteins with 168-251 amino acids and calculated molecular mass of 18.44-27.42 ku.Phylogenetic analysis indicated that there were at least 7 most recent common ancestralGPXgenes before split ofO.sativa,B.distachyonandS.bicolorand one and twoGPXgenes were lost inO.sativaandB.distachyonafter species divergence.Sequence similarity analysis showed GPX proteins had high sequence similarities of 38.0%-95.2%,and GPXs of different clades had different divergent rates.AllGPXgenes of the three species had conserved structures, and an insertion of intron occurred inSbGPX7,indicating functional divergence betweenSbGPX7 and otherGPXs.Reverse transcription PCR revealed that 15 of the 18GPXgenes were constitutive expression genes,while the other three were selectively expressed either in a specific tissue and/or in response to a specific treatment.Expression divergence indicated functional divergence.【Conclusion】GPXsin Gramineae,as key enzymes that protect cellular components from oxidative damage,had functional differences after split ofO.sativa,B.distachyonandS.bicolor.
glutathione peroxidase;Gramineae;gene evolutionary analysis;gene expression pattern
时间:2016-09-0709:03DOI:10.13207/j.cnki.jnwafu.2016.10.025
2015-04-15
吉林农业大学科研启动基金项目(201407)
杨雪(1984-),女,吉林长春人,讲师,博士,主要从事植物基因家族的功能进化研究。
E-mail:xueyang840316@163.com
Q811.4
A
1671-9387(2016)10-0176-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160907.0903.050.html
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
古今农业(2022年2期)2022-08-15 01:39:52
青年文学家(2022年1期)2022-03-11 12:27:39
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01 08:10:30
核农学报(2020年6期)2020-06-30 06:15:38
生物工程学报(2019年6期)2019-07-10 08:38:38
生物学通报(2019年1期)2019-02-15 16:33:43
生物学通报(2018年12期)2018-10-10 06:52:36
植物研究(2018年4期)2018-07-24 01:06:24
云南中医学院学报(2012年4期)2012-07-31 18:06:20
