
不同地点杨树无性系树高和胸径变异分析及生长模型构建
2016-11-10 01:16司冬晶张鑫鑫韩冬荟金允哲董元海赵光浩冷伟伟赵曦阳
植物研究 2016年4期
司冬晶 张鑫鑫 韩冬荟 金允哲 董元海 赵光浩 冷伟伟 赵曦阳*
(1.东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040; 2.龙井市林业局,龙井 133400; 3.龙井市开山屯林场,龙井 133400)
不同地点杨树无性系树高和胸径变异分析及生长模型构建
司冬晶1张鑫鑫1韩冬荟1金允哲2董元海2赵光浩3冷伟伟3赵曦阳1*
(1.东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040;2.龙井市林业局,龙井 133400;3.龙井市开山屯林场,龙井 133400)
以不同地点30个白杨杂种无性系为材料,对其不同树龄的树高和胸径进行调查,对获得数据进行变异分析并构建生长曲线模型,方差分析结果表明地点间、无性系间、地点与无性系交互作用间均达到极显著差异水平(P<0.01);树高和胸径表型变异系数变化范围分别为15.34%~23.80%和19.42%~31.88%;所有性状重复力较高,变化范围为0.797 7~0.985 9;模型构建结果表明,根据杨树无性系苗期树高和胸径构建的指数函数模型拟合度r2分别达0.82和0.87,拟合效果较好。利用模型对未测定数据进行估算,无论树高还是胸径,冠县无性系的最初生长量最高,而峰峰的树高和威县的胸径生长速率最快。基因型与环境具有明显的交互作用,不同无性系在相同地点表现差异较大,相同无性系在不同地点也呈现明显差异,表明无性系评价选择需要考虑环境因素,适地适树的进行选择。构建的模型可以为林木生长量预测提供理论依据。
白杨;树高;胸径;生长模型;变异
林木在生长发育过程中,时刻受到环境因素的影响[1~3],特别是在良种选育过程中,优良品种或无性系的筛选均需区域化试验来确定其适应性和适生范围,因此基因型和环境互作一直是育种学家研究的工作热点[4]。赵廷松[5]、黄鹏[6]和李春迤等[7]分别对核桃(Juglansregia)、国外李(Prunussp.)和日本栗(Castaneacrenata)进行品种区域化试验,筛选出在不同地点进行栽培的优良品种。在杨树育种方面,李丕军等[8]和姜岳忠等[9]也分别对杨树无性系进行多点造林,对无性系的遗传稳定性进行探讨,分别获得了适宜不同栽培区域的优良无性系。
林木生长发育是一个复杂的动态过程,为了获得更多与生长相关的信息则需要对植株的生长进程进行测定分析。随着计算机技术以及数学、统计学等多学科在林木育种中的应用,利用数学模型模拟林木生长过程的研究得到了越来越多研究者的重视[10]。树高和胸径是林木生长评价中最重要的两个因素,许多专家对部分树种树高、胸径等进行了多种模型(Exponential,Gompertz,Logistic,Richards,Morgan Mercer Flodin)的构建及生长量预测[11~14],帮助学者更好的了解植物动态生长和发育趋势,但国内外多杨树连续多年数据进行模型构建的研究较少,尤其在品种选育过程中考虑生长过程的更鲜见报道。本文主要以不同地点栽植的30个白杨派杂种无性系为材料,对1~5年的树高和胸径进行测定,对树高和地径生长过程进行数学模型构建,把数学模型的知识与植物生长发育有效结合,为林木生长、发育研究提供理论基础。
1 材料与方法
1.1 试验材料与试验林
试验材料主要包括30个白杨杂种无性系(表1)。于2006年春季在河北省邯郸市峰峰矿区苗圃场、河北省邢台市威县苗圃场、山东省聊城市冠县国有苗圃和山东省泰安市宁阳县高桥林场等4个地点营建无性系对比试验林,各地气候条件见表2,试验采用完全随机区组设计,4株小区,单行排列,4次重复,株行距3 m×4 m。
表1试验材料及杂交组合
Table1Experimentmaterialsandhybridizedcombination

无性系Clone杂交组合HybridizedcombinationP20,P22,P23,P26,P28,P30(P.alba×P.glandulosa‘1’)×P.bolleanaP42,P46,P49,P50,P53(P.tomentosa×P.bolleana)×(P.alba×P.glandulosa‘84K’)P63,P64,P67,P69,P76,P77(P.alba×P.tomentosa)×P.bolleanaP78,P83(P.tomentosa×P.bolleana)×P.tomentosa‘LM50’P85,P87,P88(P.alba×P.glandulosa)×P.bolleanaP98,P99,P101P.alba×P.bolleanaP103(P.alba×P.bolleana)×P.bolleanaP104(P.tomentosa×P.bolleana)×P.tomentosa‘LM50’P106,P107(P.tomentosa×P.bolleana)×P.tomentosa‘3’LM50选种无性系SelectedexcellentcloneP.tomentosa‘LM50’
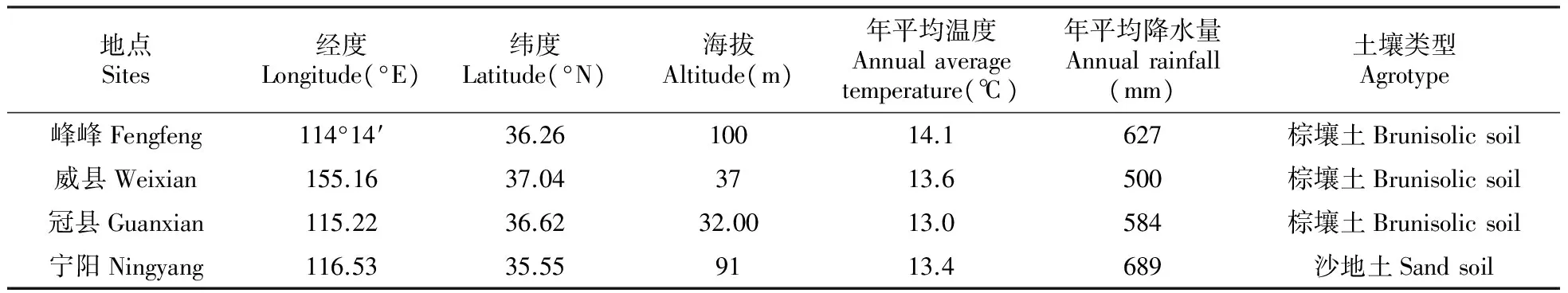
表2 不同地点主要环境因子
表32006~2010年各地点测定树高和地径情况
Table3MeasuringschedulesforHandDBHinfoursitesduring2006-2010

地点Sites树高H胸径DBH2006200720082009201020062007200820092010峰峰Fenggeng—XXXX—XXXX威县Weixian—XXXX—XXXX冠县Guanxian——XXX——XXX宁阳NingyangXXXXXXXXXX
注:X.数据已测;—.没有测定数据。
Note:X and - denotes the trees were measured or not measured,respectively.
1.2 研究方法
于2006年秋季开始对不同地点各无性系树高(H)和胸径(DBH)进行测定,具体测定次数见表3。测定过程中对各无性系进行全林测定,剔除死亡或者风折等特异性数据后进行参数计算。本研究利用双因素方差分析方法进行无性系、地点,以及交互作用分析;同时,对不同无性系的生长进程进行指数方程拟合。
1.3 分析方法
所有数据利用SPSS(Statistical Program for Social Sciences)19.0软件[15]和R软件[16]进行分析。
(1)方差分析线性模型为[17]
yij=μ+αi+βj+αβi(j)+εij
(1)
式中,μ为总体平均值,αi为无性系效应,βj为地点效应,αβi(j)为无性系与地点的交互效应,eij为环境误差;
(2)无性系重复力采用公式[18]
R=1-1/F
(2)
式中:F为方差分析的F值。
(3)表型变异系数采用公式[19]
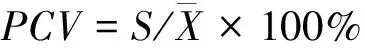
(3)

(4)曲线模拟过程
利用yijk(t)代表第t年第i个无性系在第k个环境下,无性系j的i性状的测定结果,而t=1,2,…,Tk代表在k环境条件下不同测定的时间点,因此生长性状的模型yijk(t)可以表示为以下方程:
yijk(t)=μjk(t)+ejk(t)
(4)
式中:μjk(t)代表在t时间,k地点上,无性系j的期望平均值,而ejk(t)代表环境误差。假设误差项ejk(t)遵从正态分布,不同时间点(t,t′)的误差项相关系数表示为ρ(t,t′),那么在环境k条件下方差和协方差可以利用(Tk×Tk)协方差矩阵∑k表示。
1.3.1 似然函数
令yijk(t)=(Yijk(l),…,yijk(Tk))作为在不同时间,k地点,无性系j的i性状测定结果的向量函数,假设,基因型、基因型与环境互作同时控制植物生长过程,那么不同无性系在不同地点生长参数则会不同,生长性状(y)似然函数可以表示如下:
(5)
这里Ω是方程(4)协方差矩阵中∑k和构成平均向量μjk参数的未知参数向量,而在k环境条件下无性系j的i性状在时Tk间点的多元正态分布密度函数可以表示为:
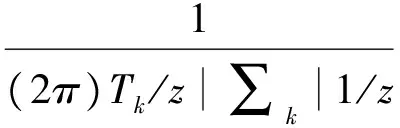
(6)
式中:μjk代表k环境条件下无性系j期望平均向量。
1.3.2 构建平均值曲线
生长轨迹可以描述为具有生物学意义的数学函数,例如logistic曲线已经被看作为成年树生长模型。本研究构建了平滑曲线模型如下:
μjk(t)=αjk×e(bjkt)
(7)
这里t代表不同的测定时间点,Ωmjk=(ajk,bjk)参数描述当t=0时无性系j在地点k的最初生长量(ajk)和生长速率(bjk)。因此生长曲线μjk(t)是由参数Ωmjk决定的。优于重复方差分析模型,该模型可以利用简单增加时间点来估计丢失时间点期望平均值。此外,指数模型只需要估计在Tk时间点的μjk平均向量的两个参数,而重复方差分析估计需要评价Tk平均值。
1.3.3 协方差矩阵模型
纵向设计需要考虑不同时间点之间性状的相互关系。测量每个单株的过程中忽略时间效应,这样获得的生长参数将会有偏差。最常用的方法是满足第一序列的自回归模型,假定剩余方差在不同时间点是固定的,表示为σ2,并且在不同时间点t1和t2之间的相关性可以表达为ρ(t1,t2)=ρ|t1-t2|。在实际应用中,假设不同时间点固定剩余方差和固定之间的相关性可能无法控制。因此,本研究使用的transform-both-sides(TBS)方法[20]进行解决。模拟和实例分析表明,该TBS模型可以提高参数估计的精度和计算效率。此外,通过分析表型数据而获得生长参数的生物信息学的解释。
1.3.4 假设检验
根据方差分析模型,可以进行很多具有生物学意义的假设检验。首先,可以检验不同无性系生长曲线是否相同,假设检验可以按照以下公式:
H0:(a1,b1)=(a2,b2)=…=(ai,bi)
H1:thenullhypothesisnottrue
如果接受H0,那么不同无性系生长曲线相同,如果接受H1,则不同无性系生长曲线不同。
另外一个假设是如果想检验基因型与环境之间是否存在显著互作关系,那么假设可以写成以下公式:
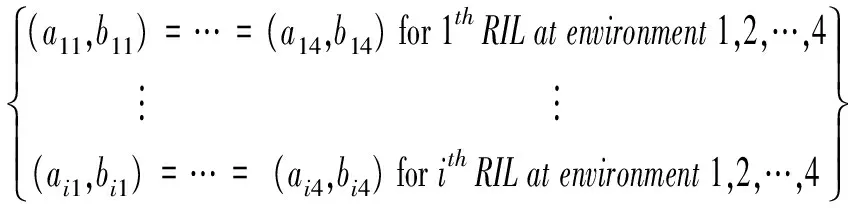
在上述假设条件下,接受H1,则相同的无性系在不同环境条件下生长曲线不同。
1.3.5 模型应用于参数估计
利用TBS的方法,平均曲线(方程1)转化为线性混合效应模型:log(y)=log(a)+b×t+log(e)。利用SAS v9.3中的MIXED程序构建log-linear模型,利用赤池信息量(Akaike information criterion,AIC)或贝叶斯信息量(Bayesian information crietreion,BIC)等评估模型拟合度和协方差矩阵。利用三型平方和进行假设检验并且利用最大似然法评价方差模型中的参数(a,b)。
2 结果与分析
2.1不同时间、不同地点无性系树高、胸径总体平均值变化及变异参数
不同地点无性系树高、胸径总体平均值见表4,2006年,宁阳所有无性系树高和胸径分别为4.32 m和3.01 cm,而到2010年,所有无性系树高和胸径分别达7.85 m和9.19 cm;从峰峰来看,2007年树高和胸径分别为6.44 m和5.88 cm,至2010年树高和胸径平均值分别达9.91 m和10.62 cm;威县2007年树高和胸径分别为5.46 m和4.47 cm,而2010年达9.62 m和10.02 cm;冠县只测定3年数据,2008年树高和胸径分别为6.70 m和6.69 cm,2010年树高和胸径分别达9.82 m和10.54 cm;从变异系数来看,随着树龄的增长,不同地点树高和胸径均呈递增趋势,变化范围分别为15.34%~23.80%和19.42%~31.88%;所有性状重复力变化范围为0.797 7~0.985 9。
表4不同地点30个杨树无性系树高、胸径变异参数
Table4VariationparametersofHandDBHof30poplarclonesindifferentsites

地点Sites树龄Age树高H胸径DBH平均值Average变异系数PCV重复力R平均值Average变异系数PCV重复力R峰峰Fengfeng20076.4419.700.96295.8823.640.967820087.6320.380.97457.9826.680.983720098.9520.690.97599.5927.820.984920109.9123.400.980910.6229.180.9859冠县Guanxian20086.7019.430.96946.9919.420.966120098.1219.500.97248.6722.950.968520109.8219.890.973010.5425.040.9805宁阳Ningyang20064.3215.340.81503.0121.480.814820075.2117.510.89595.1819.820.922820085.8318.470.92376.5621.450.941220096.8820.360.95458.3223.960.960220107.8523.360.96619.1926.750.9685威县Weixian20075.4619.410.79774.4724.110.823220086.8220.440.86526.7925.130.863220098.8621.980.90308.6127.690.884120109.6223.800.904810.0231.880.9035
2.2 方差分析结果
相同时间不同地点30个白杨杂种无性系方差分析见表5。相同树龄条件下,在无性系间、地点间及无性系与地点的交互作用间,树高和胸径均达极显著差异水平(P<0.01)。
表5不同无性系间、地点间树高和胸径方差分析表
Table5VarianceAnalysisforHandDBHamongdifferentsitesandclones

性状Traits树龄Age变异来源VariancesourcedfMSFP树高H20062007200820092010无性系Clones292.4285.406<0.01地点Sites2116.586163.867<0.01无性系Clones2918.23325.627<0.01地点×无性系Sites×Clones583.1744.462<0.01地点Sites3145.900236.742<0.01无性系Clones2940.49965.715<0.01地点×无性系Sites×Clones872.8454.617<0.01地点Sites3221.076248.283<0.01无性系Clones2967.53075.841<0.01地点×无性系Sites×Clones873.5263.960<0.01地点Sites3232.504210.277<0.01无性系Clones2986.29878.048<0.01地点×无性系Sites×Clones875.6585.117<0.01胸径DBH20062007200820092010无性系Clones291.6115.401<0.01地点Sites2123.764166.486<0.01无性系Clones2921.29528.645<0.01地点×无性系Sites×Clones582.3643.180<0.01地点Sites3110.659118.149<0.01无性系Clones2966.36070.852<0.01地点×无性系Sites×Clones874.0774.353<0.01地点Sites385.12760.020<0.01无性系Clones29122.69286.505<0.01地点×无性系Sites×Clones875.5073.883<0.01地点Sites3110.73164.039<0.01无性系Clones29195.212112.898<0.01地点×无性系Sites×Clones876.7533.905<0.01
2.3 生长曲线的拟合结果
4个地点不同无性系在不同年份下树高和胸径的曲线模拟图见图1,统计检验表明树高和胸径的方程拟合程度非常高,拟合系数r2分别达0.82和0.87,具体树高和胸径的曲线模型参数分别见表6~7,根据曲线模型推测冠县白杨杂种无性系最初树高平均值最大,达到4.591 m,其次是宁阳(4.140 m)和威县(3.431 m),最小为峰峰(3.017 m);峰峰树高生长速率最大,达到0.742,其次是威县(0.658)和冠县(0.480),最小值是宁阳(0.355);从胸径来看,峰峰、威县、冠县和宁阳白杨杂种无性系最初胸径平均值分别为2.984、2.407、3.812和3.028 cm(t检验结果表明宁阳推测值与实测值差异未达显著水平P>0.05);不同地点生长速率也不同,威县白杨杂种无性系胸径生长速率最快,达到0.919,峰峰其次(0.769)和宁阳(0.697),冠县最低,只有0.632。
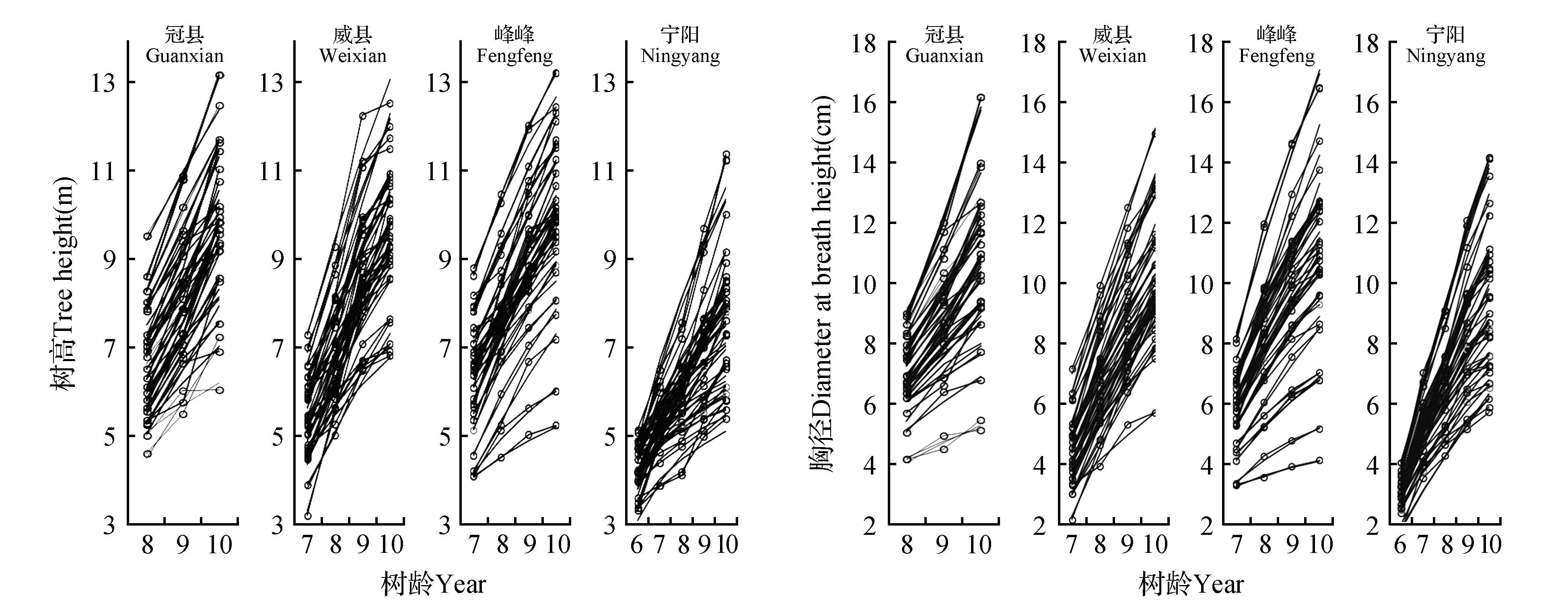
图1 4个地点不同无性系树高和胸径拟合图 (实线为观测值,虚线为模拟值)Fig.1 Matching figure of H and DBH of different clones at 4 different sites (full lines were observed values and imaginary lines were simulation values)
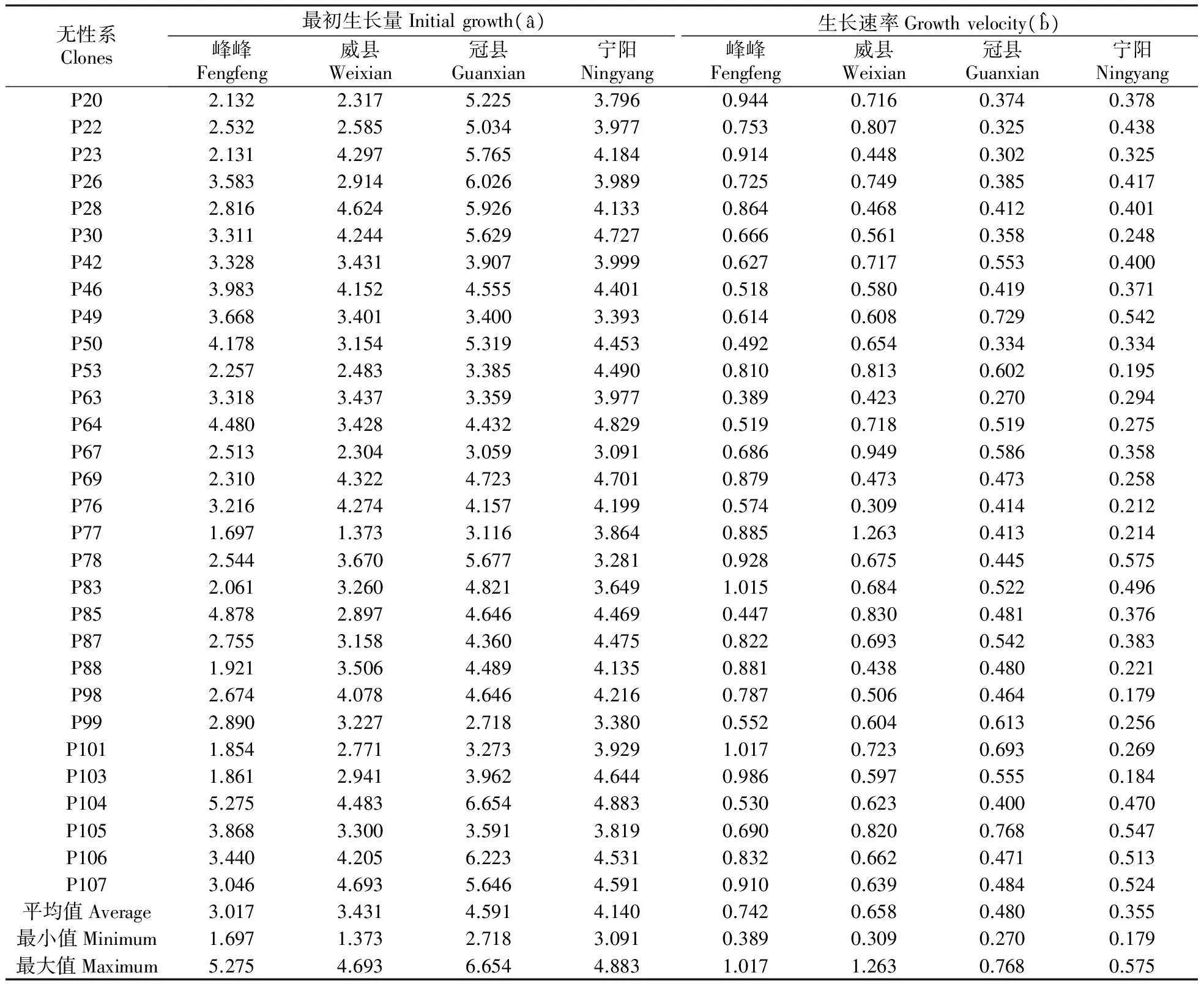
无性系Clones最初生长量Initialgrowth(^a)生长速率Growthvelocity(^b)峰峰Fengfeng威县Weixian冠县Guanxian宁阳Ningyang峰峰Fengfeng威县Weixian冠县Guanxian宁阳NingyangP202.1322.3175.2253.7960.9440.7160.3740.378P222.5322.5855.0343.9770.7530.8070.3250.438P232.1314.2975.7654.1840.9140.4480.3020.325P263.5832.9146.0263.9890.7250.7490.3850.417P282.8164.6245.9264.1330.8640.4680.4120.401P303.3114.2445.6294.7270.6660.5610.3580.248P423.3283.4313.9073.9990.6270.7170.5530.400P463.9834.1524.5554.4010.5180.5800.4190.371P493.6683.4013.4003.3930.6140.6080.7290.542P504.1783.1545.3194.4530.4920.6540.3340.334P532.2572.4833.3854.4900.8100.8130.6020.195P633.3183.4373.3593.9770.3890.4230.2700.294P644.4803.4284.4324.8290.5190.7180.5190.275P672.5132.3043.0593.0910.6860.9490.5860.358P692.3104.3224.7234.7010.8790.4730.4730.258P763.2164.2744.1574.1990.5740.3090.4140.212P771.6971.3733.1163.8640.8851.2630.4130.214P782.5443.6705.6773.2810.9280.6750.4450.575P832.0613.2604.8213.6491.0150.6840.5220.496P854.8782.8974.6464.4690.4470.8300.4810.376P872.7553.1584.3604.4750.8220.6930.5420.383P881.9213.5064.4894.1350.8810.4380.4800.221P982.6744.0784.6464.2160.7870.5060.4640.179P992.8903.2272.7183.3800.5520.6040.6130.256P1011.8542.7713.2733.9291.0170.7230.6930.269P1031.8612.9413.9624.6440.9860.5970.5550.184P1045.2754.4836.6544.8830.5300.6230.4000.470P1053.8683.3003.5913.8190.6900.8200.7680.547P1063.4404.2056.2234.5310.8320.6620.4710.513P1073.0464.6935.6464.5910.9100.6390.4840.524平均值Average3.0173.4314.5914.1400.7420.6580.4800.355最小值Minimum1.6971.3732.7183.0910.3890.3090.2700.179最大值Maximum5.2754.6936.6544.8831.0171.2630.7680.575
从无性系来看,无性系P46在4个地点胸径最初生长量最相近(冠县4.414 cm,威县4.408 cm,峰峰4.392 cm,宁阳3.934 cm),无性系P77在4个地点均呈现最低值(冠县2.500 cm,威县0.777 cm,峰峰2.689 cm,宁阳1.882 cm)。对于胸径生长速率来说,无性系P106在4个地点呈现最高生长速率(冠县0.787,威县0.990,峰峰1.154,宁阳0.822),而无性系P63在4个地点生长速率最低(冠县0.241,威县0.627,峰峰0.511,宁阳0.543)。
总的来说不同地点间无性系生长状况差异极显著(P<0.01),宁阳各无性系树高和胸径的生长量比其他三个地点无性系树高和胸径的生长量低(图1),从树高来看,冠县的最初生长量最高(4.591 m),峰峰生长速率最大(0.742),从胸径来看,同样是冠县地点最初生长量最高(3.812 cm),威县生长速率最大(0.919)(表6~7)。
2.4 基因型与环境互作关系
不同性状、不同时间基因型与环境之间的交互作用差异达极显著水平(表4)。无性系P107在峰峰、威县和宁阳地点树高和胸径均最大,在冠县也较高,但是低于P106。无性系P77在峰峰地点胸径最小,但是在威县,冠县和宁阳,无性系P63胸径最小;在峰峰、威县和冠县无性系P63树高最低,但是在宁阳,无性系P99树高最小。对无性系P107和无性系P63进行作图比较(图2),无性系P107在不同地点,不同时间树高和胸径明显高于无性系P63,在冠县地点无性系P107生长显著高于无性系P63,明显可以看出2个无性系在生长过程中基因型与环境之间的互作。
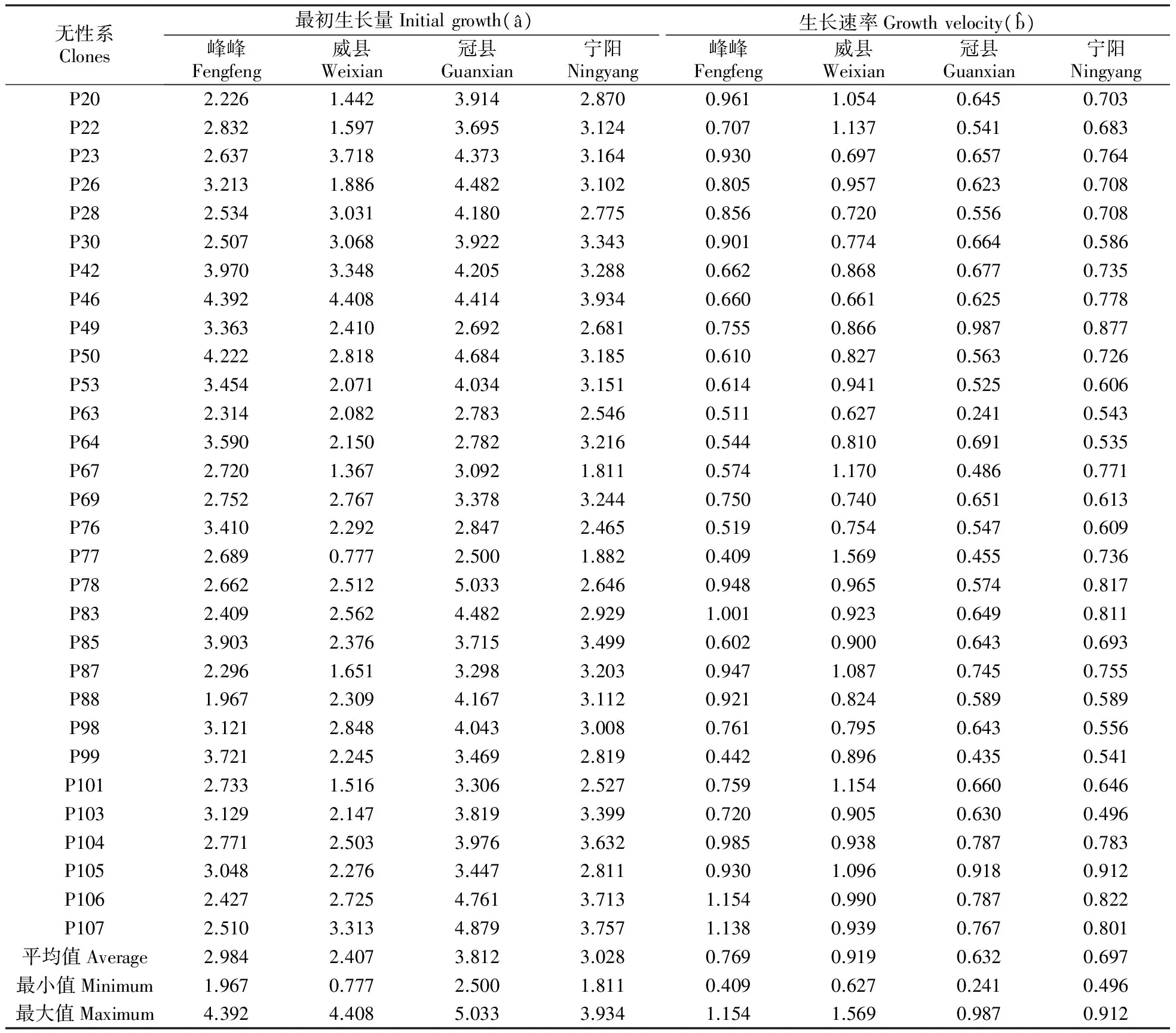
表7 不同地点各无性系胸径指数函数参数估计值
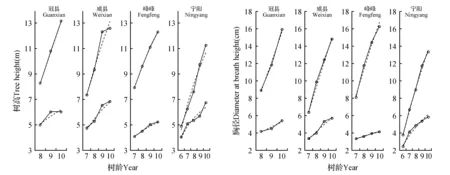
图2 无性系P63(下)和无性系P107(上)在4个地点的树高、胸径拟合图 (实线为测定值,虚线为模型估算值)Fig.2 Matching figure of H and DBH for clone P63(down) and P107(up) at four environments (full lines were observed values and imaginary lines were simulation values)
3 讨论
区域化试验是基因型与环境互作研究的热点,是对树种进行评价的理论基础及确定品种推广价值和适应范围的重要依据[21]。本研究中相同树龄条件下,不同无性系间、地点间及地点与无性系交互作用间均差异显著,表明各无性系生长受遗传与环境因素共同作用,由于地理位置、土壤环境、气候等差异导致相同无性系在不同地点变异较大,这与王军辉[22]在桤木(Alnuscremastogyne),Pliura[23]在杨树上的研究结果相近。
重复力和表型变异系数是探讨无性系遗传和变异的重要参数[24],表型变异系数越大,表明可选择空间越大,重复力越高,暗示性状无性扩繁表现越稳定。本研究中树高和胸径的表型变异系数变化范围分别为15.34%~23.80%和19.42%~31.88%,结果与Pliura[23]对魁北克杨树无性系及Liu[25]对转基因杨树的研究结果相近。但本研究中,随着树龄的增加,表型变异系数越来越大,表明随着树木的生长和发育,无性系间树高和胸径差异越来越大,即在相同入选率条件下,遗传增益越来越高。树高和胸径重复力均超过0.79,这与Kien[26]对桉树(Eucalyptus)的研究结果相近,属于高重复力,表明选择的无性系树高和胸径表现稳定,高变异和高遗传力为杨树无性系评价选择提供基础。
姜立春等[27]以80株落叶松(Larixolgensis)解析木为材料,采用Richards生长模型作为基础模型,利用S-PLUS软件汇总的NIME过程,拟合了非线性树高和胸径生长模型,采用最小信息量准则(AIC)、贝叶斯信息准则(BIC)、对数似然值和似然比检验等模型评价统计指标对不同模型的精度进行比较分析,结果表明混合模型在应用上不但能反映树高和胸径的平均预测趋势,还能用方差、协方差结构和误差相关性结构校正随机参数来反映个体间的差异。本研究主要对不同地点30个白杨杂种无性系的树高和胸径的生长量数据进行模型构建与分析,采用AIC和BIC来评估模型拟合度,统计检验表明,生长轨迹可以很好地适应指数模型(r2=0.82和0.87)。宁阳各无性系树高和胸径的生长量比其他三个地点无性系树高和胸径的生长量低,这可能与宁阳土壤是沙壤土的条件有关。利用模型对未测定数据进行估算,无论树高还是胸径,冠县无性系的最初生长量最高,峰峰树高生长速率最快,威县胸径生长速率最快。
林木生长规律预测系统可以提高林业经营水平[28]。在对苗期生长进行模拟的过程中,很多学者选择Logistic模型或者Richards模型,因为这两个模型对“S”曲线拟合效果较理想,特别是Richards生长方程,当其参数m在数轴上滑动取值时,不仅包含了Mitscherlich、Bertalanffy、Gompertz、Logistic等生长方程,而且包括了他们中间过渡类型和更为广义的形状[29],但这两个方程均需要植株生长过程达到一个周期(速生前期、速生期、速生后期)。本研究中数据是苗期数据,还未到达整个生长周期,因此没有采用上述两种模型而选择指数模型,模型拟合度高,且该模型不但可以对已知生长过程进行动态模拟,还可以根据几年的生长性状对将来生长趋势进行预测,为无性系早期选择和评价提供了一种新的方法与思路。
1.Seyed M S.Estimation of genetic parameters related to morphological traits in poplar clones[J].Americian Journal of Scientific Research,2001,27:105-110.
2.Fang S Z,Xu X Z,Lu S X,et al.Growth dynamics and biomass production in short-rotation poplar plantations:6-year results for three clones at four spacings[J].Biomass and Bioenergy,1999,17:415-425.
3.Marron N,Ceulemans R.Genetic variation of leaf traits related to productivity in aPopulusdeltoides×Populusnigrafamily[J].Can J Forest Res,2006,36:390-400.
4.Nicolas M,Sophie Y,Reinhart C.Evaluation of leaf traits for indirect selection of high yielding poplar hybrids[J].Environmental and Experimental Botany,2007,61(2):103-116.
5.赵廷松,方文亮,曾清贤.5个核桃早实杂交新品种鲁甸县区域试验[J].西北林学院学报,2007,22(5):83-85.
6.黄鹏,路生林.国外李品种区域化栽培试验[J].河北林果研究,2006,21(1):59-62.
7.李春迤,毛立仁,秦德智.日本栗引种试验初报[J].北方果树,2008(6):7-10.
8.李丕军,林思祖,李宏,等.银×新无性系二次选优及无性系的推广[J].东北林业大学学报,2009,37(2):94-95.
9.姜岳忠,李善文,秦光华,等.黑杨派无性系区域化试验初报[J].林业科学,2006,42(12):143-147.
10.Zhao X Y,Tong C F,Pang X M,et al.Functional mapping of ontogeny in flowering plants[J].Briefings in Bioinformatics,2011,13(3):317-328.
11.Zhang L J.Cross-validation of non-linear growth functions for modeling tree height-diameter relationships[J].Annals of Botany,1997,79:251-257.
12.Huang S,Titus S J,Wiens D P.Comparison of nonlinear height-diameter functions for major Alberta tree species[J].Canadian Journal of Forest Research,1992,22:1297-1304.
13.Nothdurft A,KuPin E,Lappi J.A non-linear hierarchical mixed model to describe tree height growth[J].Eur J Forest Res,2006,125:281-289.
14.Lappi J.A longitudinal analyses of height/diameter curves[J].For Sci,1997,43(4):555-569.
15.时立文.SPSS 19.0统计分析从入门到精通[M].北京:清华大学出版社,2012.
16.薛毅,陈立萍.R统计建模与R软件[M].北京:清华大学出版社,2007.
17.Hansen J K,Roulund H.Genetic parameters for spiral grain,stem form,Pilodyn and growth in 13 year old clones of Sitka Spruce(Piceasitchensis(Bong.) Carr.) [J].Silvae Genet,1996,46:107-113.
18.续九如.林木数量遗传学[M].北京:中国林业出版社,2006.
19.解孝满,李景涛,赵合娥.柳树无性系苗期遗传测定与选择[J].江苏林业科技,2008,35(3):6-14.
20.Abbott R D,Carroll RJ.Interpreting multiple logistic regression coefficients in prospective observational studies[J].American Journal of Epidemiology,1984,119(5):830-836.
21.Calinski T,Czajka S,Kaczmarek Z,et al.A mixed model analysis of variance for multi-environment variety trials[J].Stat Papers,2009,50(4):735-759.
22.王军辉,顾万春,李斌,等.桤木优良种源/ 家系的选择研究:生长的适应性和遗传稳定性分析[J].林业科学,2000,36(3):59-66.
23.Pliura A,Zhang S Y,Mackay J,et al.Genotypic variation in wood density and growth traits of poplar hybrids at four clonal trials[J].For Ecol Manag.,2007,238:92-106.
24.Safavi S A,Pourdad S A,Mohmmad T,et al.Assessment of genetic variation among safflower(CarthamustinctoriusL.) accessions using agro-morphological traits and molecular markers[J].J Food Agric Environ,2010,8:616-625.
25.Liu M R,Yin S P,Si D J,et al.Variation and genetic stability analyses of transgenic TaLEA poplar clones from four different sites in China[J].Euphytica,2015,DOI:10.1007/s10681-015-1471-7.
26.Kien ND Jansson,Harwood C,et al.Genetic variation in wood basic density and Pilodyn penetration and their relationships with growth,stem straightness and branch size forEucalyptusurophyllaS.T.Blake in Northern Vietnam[J].NZ J For Sci,2008,38:160-175.
27.姜立春,杜书立.基于非线性混合模型的东北兴安落叶松树高和直径生长模型[J].林业科学研究,2012,25(1):11-16.
28.刘洋,亢新刚,郭艳荣,等.异龄林生长动态研究进展[J].西北林学院学报,2012,27(6):146-151.
29.邢黎峰,孙明高,王元军.生物生长的Richards模型[J].生物数学学报.1998,3(3):348-353.
VarianceAnalysisandGrowthModelBuildingofHandDBHofPoplarClonesatDifferentSites
SI Dong-Jing1ZHANG Xin-Xin1HAN Dong-Hui1JIN Yun-Zhe2DONG Yuan-Hai2ZHAO Guang-Hao3LENG Wei-Wei3ZHAO Xi-Yang1*
(1.State Key Laboratory of Tree Genetics and Breeding,Northeast Forestry University,Harbin 150040;2.Forestry Bureau of Longjing,Longjing 133400;3.Kaishantun Forestry Farm of Longjing,Longjing 133400)
Thirty poplar clones in different sites as materials and tree height(H), diameter at breath height(DBH) of different clones were investigated to calculate the variance parameters and build growth models. The result showed that there exit significant difference(P<0.01) among sites, clones and the interaction of site with clone in traits H and DBH. The coefficient variation of phenotype(PCV) of H and DBH ranged from 15.34% to 23.80% and 19.42% to 31.88%, respectively. The repeatability(R) of all the investigated traits ranged from 0.797 7 to 0.985 9, which indicated a high repeatability values. The degree of fitting of H and DBH models were 0.82 and 0.87, respectively, which showed a perfect imitative effect. With the help of model to estimate the data which were not measured, Guanxian showed the highest initial H and DBH values but Fengfeng and Weixian suggested the highest growth rates for H and DBH, respectively. There was obvious interaction between genotype and environment and whatever the different clones in same site or the same clone in different sites there were all exist larger difference in H and DBH, which indicated that the environment factors should be considered when excellent clones were evaluated and selected.
poplar;tree height;diameter at breath height;growth model;variance
中央高校青年教师自主创新基金项目(2572014BA13)
司冬晶(1990—),女,硕士研究生,主要从事杨树遗传改良工作。
* 通信作者:E-mail:zhaoxyphd@163.com
2015-12-03
S792.11
A
10.7525/j.issn.1673-5102.2016.04.015
猜你喜欢
山东绿化(2021年5期)2021-11-26
小学生必读(低年级版)(2021年4期)2021-07-28
山东绿化(2020年3期)2020-07-30
公民与法治(2020年13期)2020-07-27
金桥(2019年11期)2020-01-19
红豆(2018年4期)2018-04-20
红豆(2018年4期)2018-04-20
百科探秘·航空航天(2016年6期)2016-12-01
水能经济(2016年9期)2016-10-19
机构与行政(2016年7期)2016-07-29
