
饲料脂肪和水飞蓟素对草鱼体组织脂肪酸组成的影响
2016-11-08 01:20萧培珍吉红张宝彤叶元土李森林
中国饲料 2016年14期
萧培珍,吉红,张宝彤,叶元土,李森林
(1.西北农林科技大学动物科技学院,陕西杨陵712100;2.北京市营养源研究所水产动物系统营养研究开放实验室,北京丰台100069;3.苏州大学基础医学与生命科学学院,江苏苏州215123)
饲料脂肪和水飞蓟素对草鱼体组织脂肪酸组成的影响
萧培珍1,2,吉红1*,张宝彤2,叶元土3,李森林1
(1.西北农林科技大学动物科技学院,陕西杨陵712100;2.北京市营养源研究所水产动物系统营养研究开放实验室,北京丰台100069;3.苏州大学基础医学与生命科学学院,江苏苏州215123)
为了探究饲料脂肪和添加水飞蓟素对草鱼鱼体脂肪酸组成的影响,试验采用2×3双因素设计,脂肪水平为4%和8%(LL和HL),水飞蓟素浓度为0、100、200 mg/kg(SM0,SM100,SM200),配制6组等氮等能饲料,饲喂360尾初重为(27.43±0.17)g的草鱼12周,每组4个重复。结果显示,增加饲料脂肪水平显著影响了鱼体组织脂肪酸组成(P<0.05)。与LL组相比,HL组肌肉中18∶2n-6和20∶4n-6(ARA)的比例以及腹腔脂肪组织18∶2n-6和18∶3n-3比例分别增加了60.99%、22.72%、64.40%、161.29%,而HL组腹腔脂肪组织22∶6n-3(DHA)比例显著降低了26.76%,HL组肝胰脏中的18∶2n-6(ARA),20∶5n-3(EPA)和DHA的比例分别降低了44.68%、63.59%、49.63%和74.51%(P<0.05)。水飞蓟素添加对肌肉、腹腔脂肪和肝脏的脂肪酸组成比例均无显著影响(P>0.05)。组织与饲料脂肪酸组成相关性结果显示,肌肉和腹腔脂肪组织相关性(r=0.91,0.91)均显著高于肝胰脏(r=0.59)(P<0.05);与LL组相比,HL组的腹腔脂肪组织的相关性显著提高了10.34%(r=0.96和r=0.87);而HL组肝胰脏的相关性则显著降低了44.74%(r=0.42和r=0.76);水飞蓟素添加显著影响了肌肉和腹腔脂肪组织相关性,SM100组肌肉的相关性显著高于SM200组,增加了3.33%(r=0.93和r=0.90;P<0.05);在低脂肪水平时,SM100(r=0.91)和SM200(r=0.87)组的腹腔脂肪组织相关性显著高于SM0组(r=0.84)(P<0.05)。研究表明,增加饲料脂肪水平,导致各组织与饲料脂肪酸组成的相关性存在较大差异,这可能与不同组织在应对饲料脂肪水平以及脂肪酸利用和转化等适应或调节策略不同有关。在低脂肪水平下,添加水飞蓟素有利于提高腹腔脂肪组织与饲料脂肪酸组成的相关性;水飞蓟素对不同组织与饲料脂肪酸组成的相关性的影响存在组织差异性。
草鱼;脂肪;水飞蓟素;脂肪酸
草鱼是我国主要的淡水养殖品种之一。研究报道,草鱼具有较低的利用脂肪的能力,在4%左右(Du等,2005)。另有研究报道,草鱼能够利用较高(6%~8.5%)的饲料脂肪水平(Köprücü,2012;刘玮和任根本,1995)。饲料中过高脂肪导致鱼体脂肪过度蓄积,出现如腹腔脂肪堆积、脂肪肝、肌肉风味发生改变等现象(Du等,2008);易导致脂肪酸发生脂质过氧化反应,尤其是多不饱和脂肪酸(PUFA)摄入量增加时,导致脂质过氧化发生率增加(Halve和Hardy,2002;Stéphan等,1995),进而引起组织中PUFA降低,特别是高不饱和脂肪酸,如20∶4n-6(ARA)、20∶5n-3(EPA)、22∶6n-3(DHA)(Giordano和Visioli,2014)。由于油脂价格低廉,饲料中较高比例油脂使用成为当前生产养殖中饲料配方的主要形式。在一定范围内增加饲料脂肪水平,提高鱼体生长性能,同时最大程度促进饲料脂肪酸尤其高不饱和脂肪酸在组织中沉积,对改善鱼体组织尤其是可食部分的营养价值具有重要意义。
水飞蓟素(Silymarin)是紫花水飞蓟的干燥果实经提取、精制而得到的黄酮类化合物。研究发现,水飞蓟素可抑制脂肪生成,促进脂质分解和抑制糖原异生,影响脂蛋白组成,促进胆汁酸合成,影响脂质代谢(Yao等,2013;Ka等,2009;Sobolová等,2006;Škottová等,2003)。然而,有关脂肪水平和水飞蓟素对鱼体组织脂肪酸影响鲜见报道。
本试验通过对饲料脂肪酸和草鱼不同组织脂肪酸组成的变化及相关性研究,探讨饲料脂肪水平和水飞蓟素对鱼体不同组织脂肪酸组成的影响,为今后饲料配方设计以及水飞蓟素在水产动物饲料中应用提供参考。
1 材料与方法
1.1试验饲料本试验采用2×3双因素设计,配制等氮等能半纯化饲料。设计2个脂肪水平(4%和8%,简写为LL和HL)和3个水飞蓟素浓度(0,100,200 mg/kg,简写为SM0,SM100,SM200),配制6组试验饲料。试验饲料以鱼粉、酪蛋白为蛋白源,以豆油为脂肪源,以糊精和玉米淀粉作为糖源,并调节玉米淀粉和微晶纤维素来保持等氮等能,以羧甲基纤维素钠为粘结剂。紫外法检测水飞蓟素含量为77.80%(中国药典2010版紫外法)。
各原料均粉碎过60目筛,分别按照基础配方中各原料比例进行添加、混合均匀。将水飞蓟素与复合维生素进行混合,使其达到均一状态后加入到上述基础配方中混合均匀。之后将豆油加入到基础原料中混合均匀,再加适量蒸馏水混合均匀,用小型饲料颗粒机加工成粒径为2 mm的颗粒饲料,常温风干后放置-20℃冰箱保存备用。饲料配方及营养水平见表1。
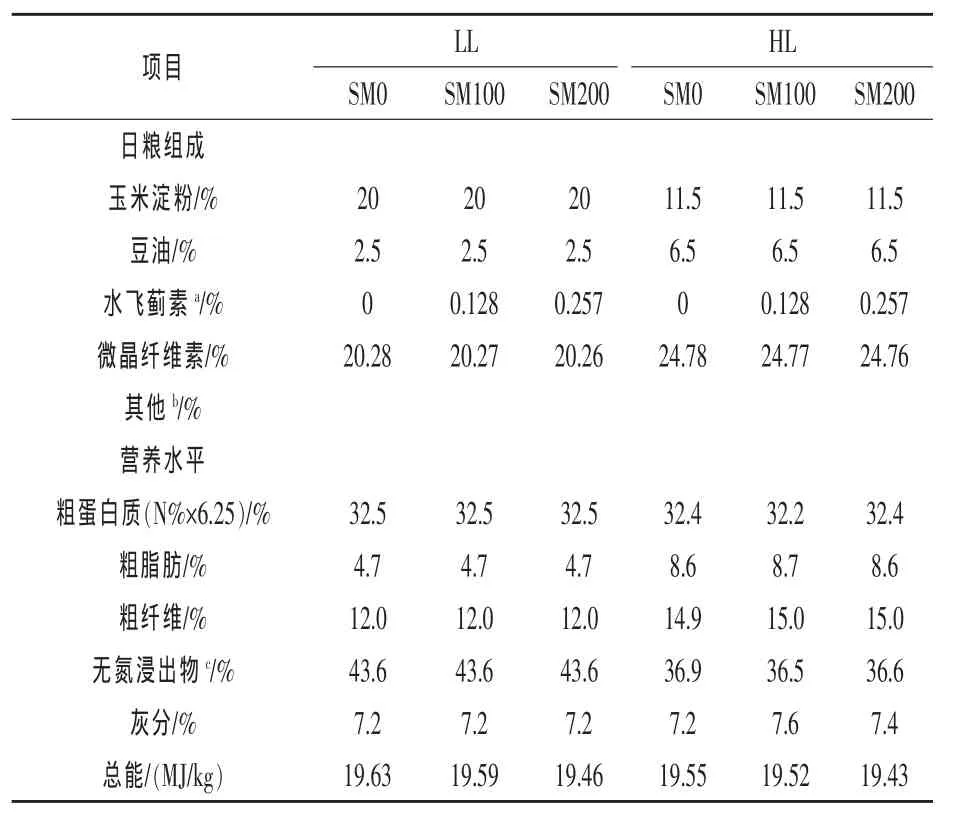
表1 饲料配方组成和营养水平
1.2饲养管理试验草鱼种购于江苏盐城华晨水产养殖场,在苏州大学基础医学与生命科学学周。试验前饲喂基础饲料(LL-SM0)进行驯化,待鱼适应循环系统后进行正式饲喂。将360尾初始体重为(27.43±0.17)g的草鱼在循环水养殖系统中进行饲喂,随机分配在24个缸中,每缸15尾,共6个处理组,每组4个重复。养殖周期为12周。每天按照鱼体重的3%~5%进行投喂,投喂2次,分别是08∶00~09∶00和16∶00~17∶00。每隔2周进行称重,并定期调整投喂率。
养殖缸水体体积为300 L,试验期间光周期为12 h光照/12 h黑暗,水温25~28.1℃,溶解氧>6.0 mg/L,总氨<0.1 mg/L,亚硝酸盐<0.1 mg/L,pH 7.0~7.4。水质符合渔业水质标准。
1.3样品采集与分析
1.3.1样品采集试验结束,停食24 h后,用MS222 100 mg/L麻醉后采集样品。每缸取6尾鱼进行解剖,取肌肉、肝胰脏和腹腔脂肪,放置-20℃冰箱保存待用。
1.3.2样品测定105℃烘干法测定饲料、肌肉、腹腔脂肪和肝胰脏水分含量。干燥后的样品用于营养成分和脂肪酸分析。粗蛋白质(N×6.25)含量采用凯氏定氮法进行测定;粗脂肪采用索氏抽提法测定;灰分550℃灰化12 h后测定;饲料中的粗纤维含量采用纤维分析仪测定;饲料总能采用XRY-1C型氧弹式热量计测定。
饲料和组织的脂肪酸组成采用气相色谱仪进行测定(Tian等,2015);结果采用面积归一法计算不同脂肪酸含量,以总脂肪酸的百分比的形式表示。饲料脂肪酸百分比组成见表2。
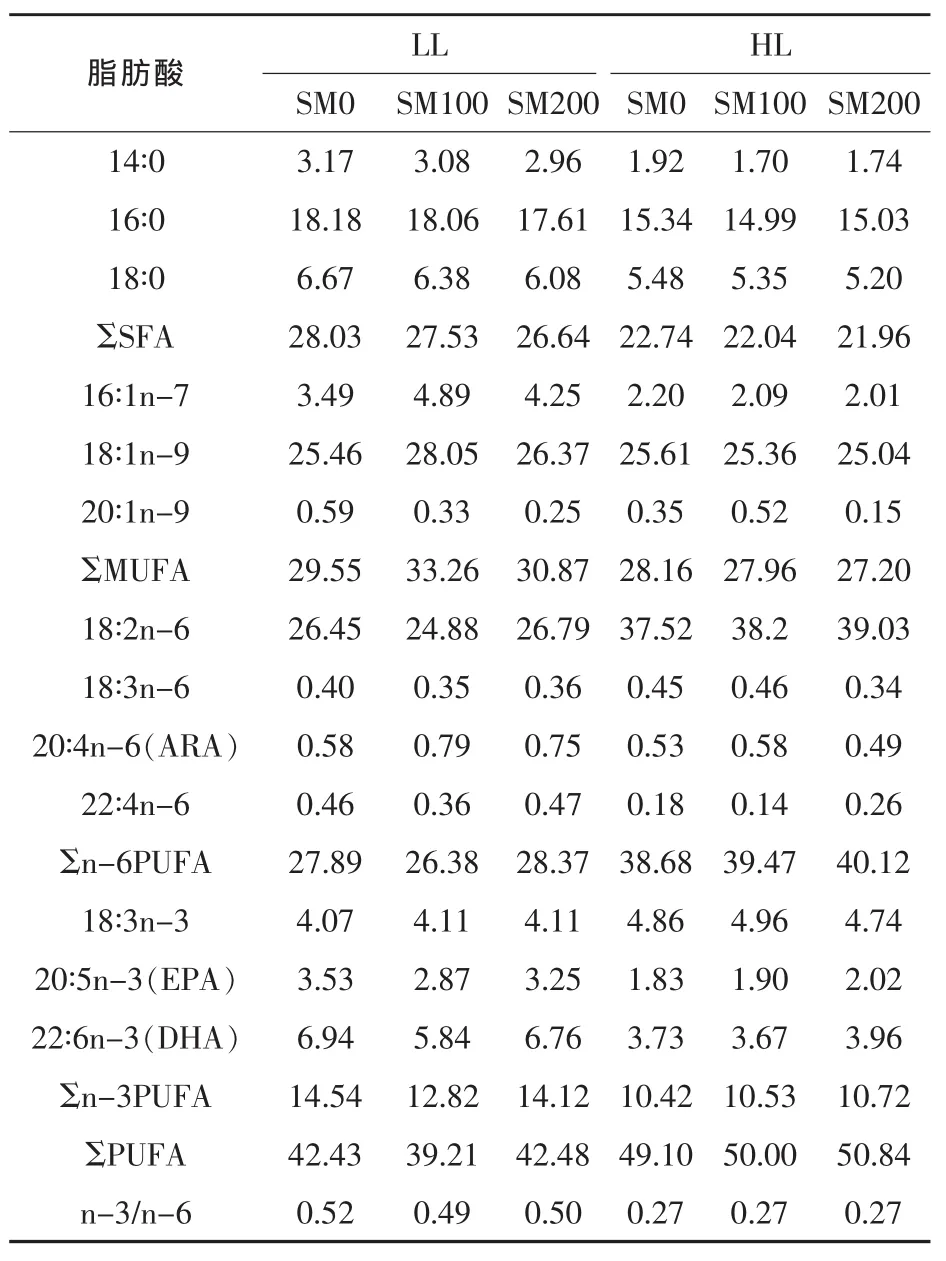
表2 试验饲料的脂肪酸组成(%总脂肪酸)
1.4统计方法试验结果以平均值表示。利用PASW 18.0软件对数据进行处理,采用双因素方差对脂肪水平、水飞蓟素、脂肪和水飞蓟素交互作用进行分析。对处理组间的组织脂肪酸组成、组织和饲料脂肪酸组成的相关性进行了单因素方差分析,存在显著性差异时进一步采用Duncan’s法进行多重比较分析。显著水平为P<0.05。
2 结果
2.1对鱼体脂肪酸组成的影响
2.1.1饲料脂肪和水飞蓟素对肌肉脂肪酸组成的影响饲料脂肪水平和水飞蓟素对草鱼肌肉脂肪酸组成的影响见表3。结果显示,饱和脂肪酸结果中,与LL组比较,HL组的14∶0比例显著降低了12.57%(1.86%,1.62%),而ΣMUFA、16∶1n-7和18∶1n-9的比例显著减少,分别降低了19.11%、44.26%和13.52%(P<0.05)。14∶0和16∶0比例受脂肪和水飞蓟素的交互影响(P<0.05)。在高脂肪水平时,14∶0比例在SM100组显著低于SM200组(分别为1.79%和1.67%),而16∶0比例则是SM100组显著高于SM200组(分别为21.85%和23.16%;P<0.05)。16∶0、18∶0、ΣSFA和20∶1n-9比例不受脂肪水平影响(P>0.05)。18∶0、ΣSFA、ΣMUFA、16∶1n-7、18∶1n-9和20∶1n-9的比例均不受水飞蓟素、脂肪与水飞蓟素交互作用的影响(P>0.05)。
多不饱和脂肪酸中,HL组的18∶2n-6、ARA、Σn-6PUFA和ΣPUFA的比例显著高于LL组,分别增加了60.99%、22.72%、45.41%和21.51%,而HL组n-3/n-6比值则比LL组显著降低了35.32%(P<0.05)。18∶3n-6、22∶4n-6、18∶3n-3、EPA、DHA、Σn-3PUFA比例不受脂肪水平的影响(P>0.05)。Σn-6PUF、Σn-3PUFA、ΣPUFA、n-3/n-6比值以及其余多不饱和脂肪比例均不受水飞蓟素、水飞蓟素和脂肪的交互作用的影响(P>0.05)。
2.1.2饲料脂肪和水飞蓟素对腹腔脂肪组织脂肪酸组成的影响饲料脂肪水平和水飞蓟素对草鱼腹腔脂肪的脂肪酸组成影响见表4。结果显示,与LL组相比,HL组的14∶0、ΣSFA、ΣMUFA、16∶1n-7和18∶1n-9的比例分别降低了18.14%、13.02%,28.59%、39.46%和15.80%(P<0.05),而16∶0、18∶0和20∶1n-9的比例均不受脂肪水平的影响(P>0.05)。多不饱和脂肪酸结果显示,HL组的18∶2n-6、18∶3n-6、ARA、18∶3n-3、Σn-6PUFA、Σn-3PUFA和ΣPUFA显著高于LL组,分别提高了64.40%、28.46%、62.21%、161.29%、75.37%、43.88%和69.66%;而HL组DHA和n-3/n-6比值则显著降低,分别为1.73%和0.19%(P<0.05)。腹腔脂肪组织的所有脂肪酸比例均不受水飞蓟素、脂肪和水飞蓟素的交互作用的影响(P>0.05)。
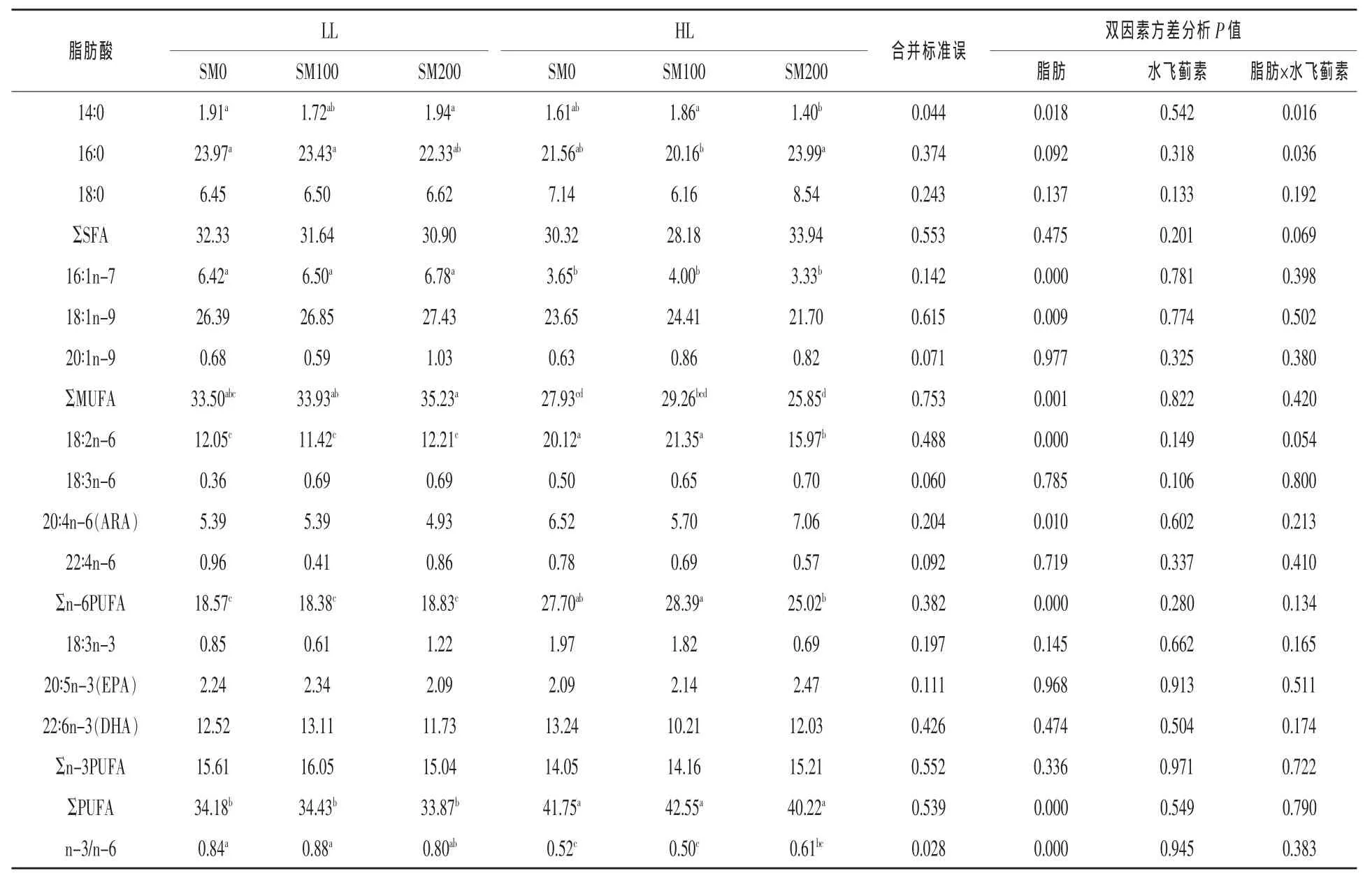
表3 饲料脂肪和水飞蓟素对草鱼肌肉脂肪酸组成的影响(%总脂肪酸)
2.1.3饲料脂肪和水飞蓟素对肝胰脏脂肪酸组成的影响饲料脂肪水平和水飞蓟素对草鱼肝胰脏脂肪酸组成的影响见表5。结果显示,HL组的16∶0、18∶0和ΣSFA的比例均显著高于LL组(P<0.05);而HL组的16∶1n-7、20∶1n-9和ΣMUFA的比例显著低于LL组,分别降低了36.34%、43.31%和6.42%(P<0.05)。14∶0和18∶1n-9的比例不受脂肪水平的影响(P>0.05)。
多不饱和脂肪酸中,HL组的18∶2n-6、ARA、EPA、DHA以及Σn-3 PUFA比例显著低于LL组,分别降低了44.68%、63.59%、49.63%、74.51%和 51.68%;而HL组18∶3n-6和22∶4n-6比例显著高于LL组(P<0.05)。ΣPUFA和n-3/n-6比值均不受脂肪水平的影响(P>0.05)。肝胰脏所有脂肪酸比例均不受水飞蓟素水平、脂肪和水飞蓟素二者交互作用的影响(P>0.05)。
2.2组织与饲料脂肪酸组成的相关性分析比较了饲料处理和不同组织对组织与饲料脂肪酸组成相关性的影响见表6。结果显示,不同的饲料处理组和3种组织的相关性均显著影响组织与饲料脂肪酸组成的相关性(P<0.05);3种组织中,肌肉和腹腔脂肪组织与饲料脂肪酸的相关性(r= 0.91,0.91)均显著高于肝胰脏(r=0.59)(P<0.05)。其次,对组织与饲料脂肪酸组成相关性进行双因素分析,结果显示肌肉与饲料脂肪酸组成的相关性受水飞蓟素水平的影响,但不受脂肪水平的影响(P<0.05)。肌肉与饲料脂肪酸的相关系数在SM100组比SM200组增加了3.33%(P<0.05),但均与SM0组(r=0.91)无差异。腹腔脂肪组织与饲料脂肪组成的相关性受脂肪、水飞蓟素以及二者的交互影响(P<0.05),其相关性在HL组比LL组增加了10.34%;在LL组中,SM100和SM200的相关系数显著高于SM0(P<0.05)。肝胰脏与饲料脂肪酸组成的相关性在HL组显著低于LL组,降低了44.74%(P<0.05),但不受水飞蓟素以及脂肪和水飞蓟素的交互作用的影响(P>0.05)。
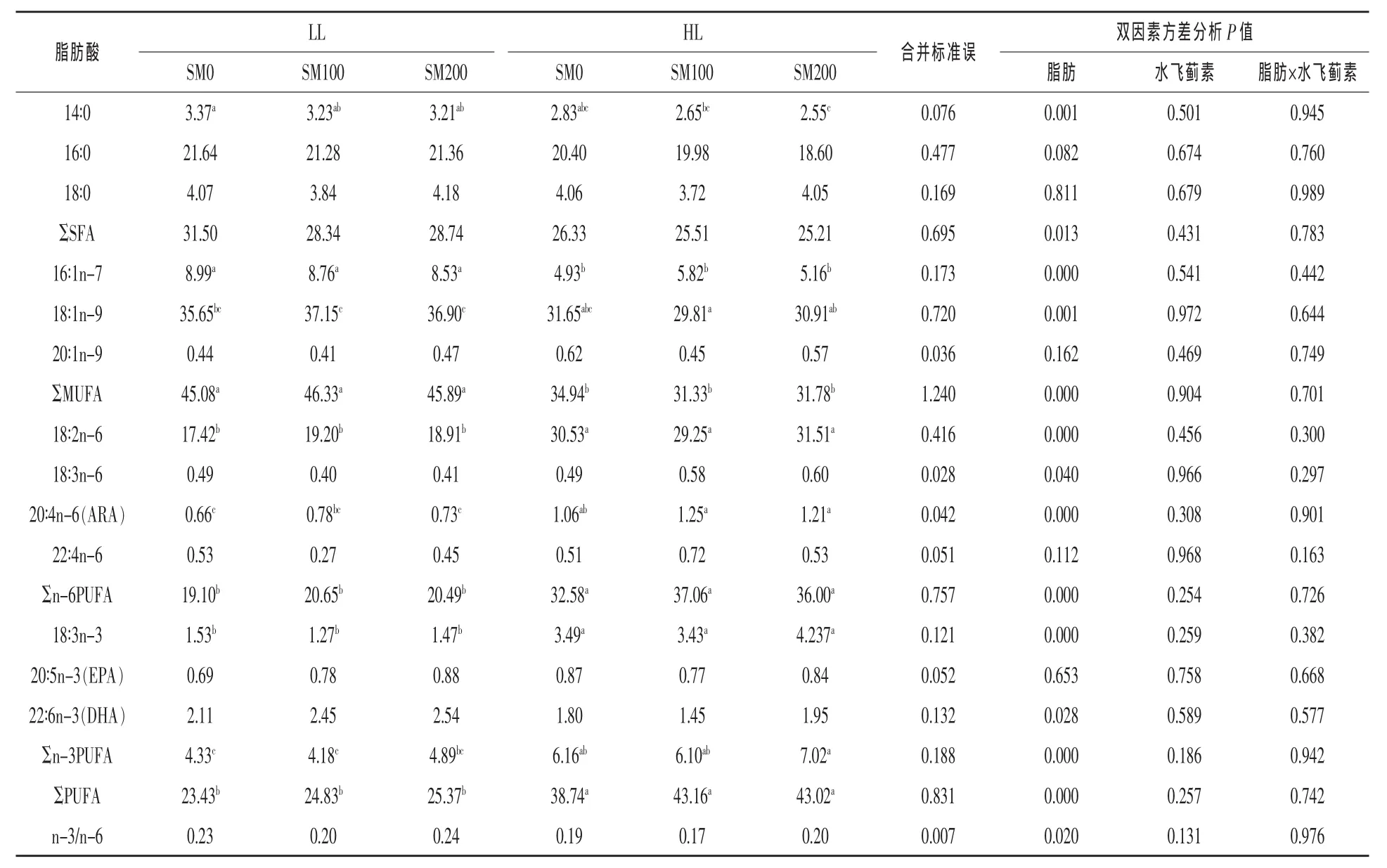
表4 饲料脂肪和水飞蓟素对草鱼腹腔脂肪组织脂肪酸组成的影响(%总脂肪酸)
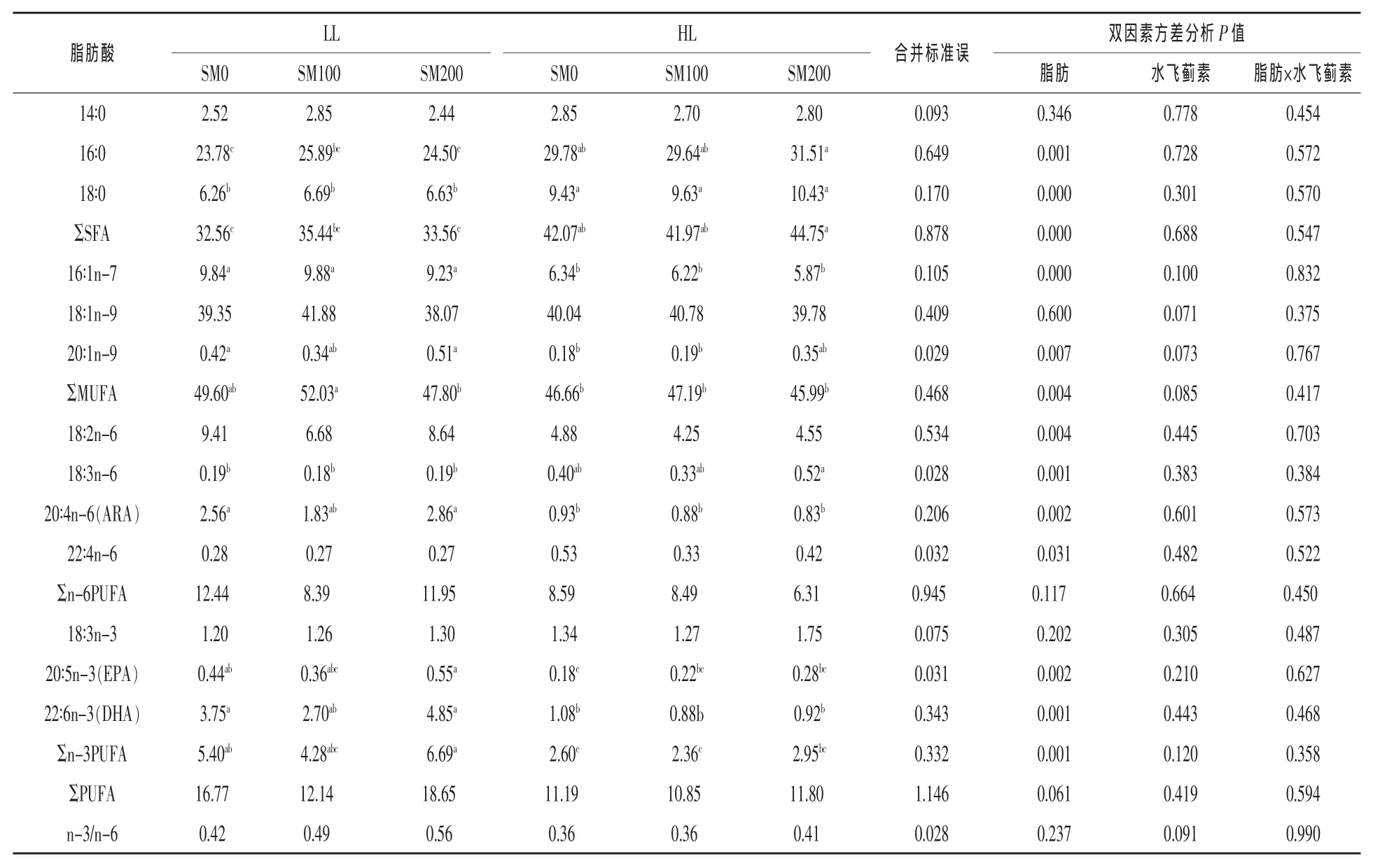
表5 饲料脂肪和水飞蓟素对草鱼肝胰脏脂肪酸组成的影响(%总脂肪酸)
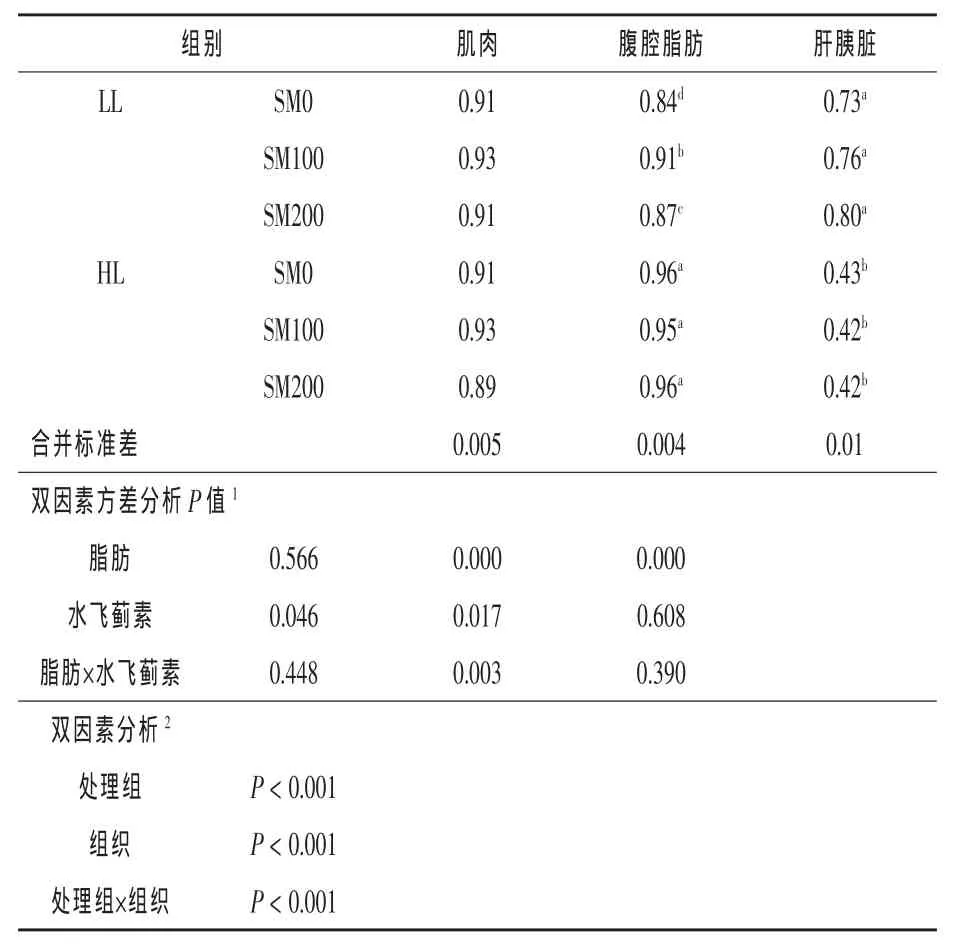
表6 不同组织脂肪酸与饲料脂肪酸组成的相关性分析
3 讨论
鱼类必需脂肪酸是指鱼体自身不能合成,必需由饲料提供的脂肪酸。淡水鱼类具有将18碳的PUFA如亚油酸(18∶2n-6)和α-亚麻酸(18∶3n-3)转化为长链多不饱和脂肪酸(≥20碳,且含有3个或以上双键的PUFA,如ARA,EPA和DHA)的能力(Tocher,2003)。必需脂肪酸不仅用于满足鱼体生长,还具有许多生理功能,如构成细胞膜的重要组成物质、可影响细胞上的蛋白功能(如受体、载体和酶),脂肪酸缺乏则导致生长异常、饲料系数降低、抗病力降低,甚至出现死亡。先前研究报道草鱼(平均初重3.9~4.2 g)对亚油酸(18∶2n-6)和亚麻酸(18∶3n-3)的需求量分别为饲料干重的1.0%和0.5%~1%(Takeuchi等,1991)。研究证实高能量饲料促进鱼体生长性能提高(NRC,2011),增加必需脂肪酸需求量将会满足最适生长和健康的需要(Tocher,2015、2003)。本研究中,随着脂肪水平即豆油添加比例的增加,饲料中的18∶2n-6(亚油酸)和18∶3n-3(亚麻酸)比例增加,证实了这一结果。
鱼体组织脂肪酸组成受饲料脂肪酸组成的影响,即同一组织中不同脂肪酸组成比例受饲料中脂肪酸组成比例的影响(Kissinger等,2016; Peng等,2014)。此外,鱼体组织脂肪酸组成受脂肪酸的消化、转运、吸收、延长和去饱和加工、β-氧化、沉积等影响(Tocher等,2010)。相关系数可用于评估组织与饲料脂肪酸组成的变化情况。本试验中,肌肉和腹腔脂肪组织的相关性显著高于肝胰脏(表6)。肌肉与饲料脂肪酸组成的相关性高(r≥0.89),这与大西洋鲑分别摄食含菜油和棕榈油的饲料后,肌肉脂肪酸组成与饲料呈显著的正相关性的结果一致(Bell等,2002,2001)。腹腔脂肪组织作为鱼体脂肪沉积主要部位之一,是鱼体能量蓄积的主要场所(Tocher,2003),饲料脂肪一部分被鱼体用于分解提供能量,大部分则被重新合成甘油三酯或者膜磷脂的形式储存在脂肪组织中,这可能是导致其与饲料脂肪酸组成相关性高的原因(r为0.84~0.96)。肝胰脏作为脂肪代谢的主要器官,参与脂肪酸合成、分解、转运等,具有自我适应性调节机制来维持自身脂肪酸组成,或改变肝胰脏脂蛋白分泌来影响脂肪酸组成,或在摄入高脂或低脂饲料后抑制或促进脂肪酸合成。本研究发现,肝胰脏与饲料脂肪酸组成的相关性在LL组高于HL组,推测可能原因是肝胰脏在应对不同脂肪水平饲料时采取了不同的适应策略来适应饲料的变化,导致肝脏脂肪酸组成改变。
n-6系列PUFA,HL组肌肉中18∶2n-6和ARA的比例以及腹腔脂肪组织18∶2n-6肝胰脏中的18∶3n-6和22∶4n-6比例显著增加,而肝胰脏中的18∶2n-6和ARA下降,这可能因本研究选择豆油为主要油脂源,豆油中含有较高比例的18∶2n-6,导致HL组饲料中18∶2n-6的比例增加,影响组织中n-6系列PUFA的比例,表明随着饲料18∶2n-6比例增加可影响草鱼组织中的n-6系列的脂肪酸的比例,而ARA和22∶4n-6比例增加证明了草鱼能够通过一系列的去饱和酶和碳链延长酶作用将18∶2n-6转化为其他高不饱和脂肪酸如ARA,22∶4n-6等,这与先前草鱼(曹俊明等,1997)、团头鲂(Li等,2016)和松浦镜鲤(Tian等,2015)上报道一致。本研究中,肝脏组织中18∶2n-6比例下降,这可能与肝脏通过去饱和酶和/或延长酶作用将更多的18∶2n-6合成其他的高不饱和脂肪酸如22∶4n-6有关。
n-3系列的PUFA,腹腔脂肪组织18∶3n-3比例增加,DHA比例降低,肝胰脏EPA和DHA比例均降低,可能是受组织中n-3和n-6PUFA竞争性抑制作用的影响(Sargent等,2002),即鱼体在分别以18∶2n-6和18∶3n-3为底物,均需要延长酶(延长酶2和延长酶5)和去饱和酶(△5和△6去饱和酶)(Morais等2009;Li等,2016),因饲料中豆油添加比例增加而导致饲料中18∶2n-6比例高于18∶3n-3比例,结果使较多的去饱和酶和延长酶用于满足较多18∶2n-6底物的需求以致合成更多的n-6系列PUFA,而n-3系列PUFA合成比例较少,组织中n-3/n-6比值降低也证明这一结果。此外,本试验各处理组添加相同比例的鱼粉,鱼粉中含有较高的EPA和DHA,随着饲料脂肪水平增加,EPA和DHA占脂肪酸的比例降低,这可能也是导致鱼体组织中n-3系列PUFA尤其EPA和DHA比例降低的原因。
随着脂肪水平增加,肌肉和腹腔脂肪组织中的ΣSFA和ΣMUFA比例降低,而ΣPUFA比例增加。PUFA作为结构物质用于机体结构组成,可影响肌肉的氧化稳定性以及肉质的风味和色泽(Wood等,2008),肌肉PUFA比例增加有利于满足人类营养需求。SFA和MUFA是鱼体生长过程中主要的能量来源(Tocher等,2003)。腹腔脂肪组织中ΣSFA和ΣMUFA降低可能是被消耗提供能量,PUFA也是细胞膜结构和功能的主要组成部分,ΣPUFA比例增加,PUFA则被优先沉积或保留,对维持细胞膜的流动具有重要作用,这将保障腹腔脂肪在低温期不凝固,有利于鱼体顺利越冬。SFA是机体脂肪酸从头合成的主要来源(Sargent等,2002),肝脏作为脂质代谢的主要场所,参与脂质的合成、水解和氧化供能,这可能是影响肝脏中SFA和MUFA变化的主要原因。
水飞蓟素是从紫花水飞蓟的果实中提取,属于黄酮类化合物。研究报道水飞蓟素能够促进胆汁酸合成,而胆汁酸能够提高脂肪酶的活力,促进脂肪的消化吸收(Romański等,2007;Crocenzi等,2003)。另研究报道,水飞蓟素通过促进脂质分解进而降低内脏脂肪蓄积(Yao等,2013)。腹腔脂肪组织是脂肪沉积的主要部位,结果显示在低脂水平下,水飞蓟素添加显著提高了组织与饲料脂肪酸的相关性,表明水飞蓟素添加有利于提高腹腔脂肪组织与饲料脂肪酸组成的相关性。然而,水飞蓟素对肝胰脏与饲料脂肪酸组成的相关性无影响,表明水飞蓟素对不同组织与饲料脂肪酸组成相关性的影响可能存在组织差异性。此外,本研究中发现SM100组的肌肉与饲料脂肪酸组成的相关性显著高于SM200组,显示了不同水飞蓟素浓度对提高组织相关性存在差异,可能存在剂量依赖关系,由于本研究未进行单因素梯度试验研究,故有待进一步研究。
4 小结
综上所述,增加饲料脂肪水平,显著影响了草鱼组织(肌肉、腹腔脂肪组织和肝胰脏)的脂肪酸组成,而添加水飞蓟素对其无显著影响。不同组织在应对饲料脂肪水平改变时的适应或调节策略不同,造成不同组织与饲料脂肪酸组成的相关性存在组织特异性。水飞蓟素可能通过促进脂肪的消化吸收提高草鱼腹腔脂肪组织与低脂肪水平饲料的相关性。该研究结果为饲料脂肪水平设置及水飞蓟素作为一种潜在的添加剂在养殖鱼类中的应用提供参考。
[1]曹俊明,刘永坚,劳彩玲,等.饲料中不同脂肪酸对草鱼组织脂质含量和脂肪酸构成的影响[J].动物营养学报,1997,9(3):36~44.
[2]刘玮,任本根.饲料中不同脂肪含量对草鱼稚鱼生长的影响[J].江西科学,1995,4:219~223.
[3]Bell J G,McEvoy J,Tocher D R,et al.Replacement of fish oil with rapeseed oil in diets of Atlantic salmon(Salmo salar)affects tissue lipid compositions and hepatocyte fatty acid metabolism[J].The Journal of Nutrition,2001,131(5):1535~1543.
[4]Bell J G,Henderson R J,Tocher D R,et al.Substituting fish oil with crude palm oil in the diet of Atlantic salmon(Salmo salar)affects muscle fatty acid composition and hepatic fatty acid metabolism[J].The Journal of Nutrition,2002,132(2):222~230.
[5]Crocenzi F A,Pozzi E J S,Pellegrino J M,et al.Preventive effect of silymarin against taurolithocholate-induced cholestasis in the rat[J].Biochemical pharmacology.2003,66,355~364.
[6]Du Z Y,Liu Y J,Tian L X,et al.Effect of dietary lipid level on growth,feed utilization and body composition by juvenile grass carp(Ctenopharyngodon idella)[J].Aquaculture Nutrition,2005 11,139~146.
[7]Du Z Y,Clouet P,Degrace P,et al.Hypolipidaemic effects of fenofibrate and fasting in the herbivorous grass carp(Ctenopharyngodon idella)fed a highfat diet[J].British journal of nutrition.2008,100,1200~1212.
[8]Giordano E,Visioli F.Long-chain omega 3 fatty acids:molecular bases of potential antioxidant actions[J].Prostaglandins,Leukotrienes and Essential Fatty Acids,2014,90(1):1~4.
[9]Halver J E,Hardy R W.Fish nutrition[A].Academic press[C].2002
[10]Ka S O,Kim K A,Kwon K B,et al.Silibinin attenuates adipogenesis in 3T3-L1 preadipocytes through a potential upregulation of the insig pathway[J].International journal of molecular medicine,2009,23(5):633~637.
[11]Kissinger K R,García-Ortega A,Trushenski J T.Partial fish meal replacement by soy protein concentrate,squid and algal meals in low fish-oil diets containing Schizochytrium limacinum for longfin yellowtail Seriola rivoliana[J]. Aquaculture,2016,452:37~44.
[12]Köprücü K.Effects of dietary protein and lipid levels on growth,feed utilization and body composition of juvenile grass carp(Ctenopharyngodon idella)[J].Journal of Fisheries Sciences,2012,6(3),243~251.
[13]Li Y,Liang X,Zhang Y,Zhang Y,et al.Effects of different dietary soybean oil levels on growth,lipid deposition,tissues fatty acid composition and hepatic lipid metabolism related gene expressions in blunt snout bream(Megalobrama amblycephala)juvenile[J].Aquaculture,2016,451:16~23.
[14]Morais S,Monroig O,Zheng X,et al.Highly unsaturated fatty acid synthesis in Atlantic salmon:characterization of Elovl5-and Elovl2-like elongases[J].Marine Biotechnology,2009,11(5):627~639.
[15]National Research Council(NRC)[M],Nutrient Requirements of Fish and Shrimp.The National Academies Press,Washington DC.2011.
[16]Peng M,Xu W,Mai K,et al.Growth performance,lipid deposition and hepatic lipid metabolism related gene expression in juvenile turbot(Scophthalmus maximus L.)fed diets with various fish oil substitution levels by soybean oil[J].Aquaculture,2014,433:442~449.
[17]Romański KW.The role and mechanism of action of bile acids within the digestive system-bile acids in the liver and bile[J].Adv Clin Exp Med,2007,16:793~799.
[18]Sargent J R,Tocher D R,Bell J G.The lipids[J].Fish nutrition,2002,3:181~257.
[19]Stéphan G,Guillaume J,Lamour F.Lipid peroxidation in turbot(Scophthalmus maximus)tissue:effect of dietary vitamin E and dietary n-6 or n-3 polyunsaturated fatty acids[J].Aquaculture,1995,130,251~268.
[20]Škottová N,R,Urbánek K,et al.Effects of polyphenolic fraction of silymarin on lipoprotein profile in rats fed cholesterol-rich diets[J].Pharmacological research,2003,47,17~26.
[21]Sobolová L,Škottová N,R,et al.Effect of silymarin and its polyphenolic fraction on cholesterol absorption in rats[J].Pharmacological research,2006,53,104~112.
[22]Tian J J,Lei C X,Ji H.Influence of dietary linoleic acid(18:2n-6)and α -linolenic acid(18:3n-3)ratio on fatty acid composition of different tissues in freshwater fish Songpu mirror carp,Cyprinus Carpio[J].Aquaculture Research,2015.1~15.
[23]Takeuchi T,Watanabe K,Yong W Y,et al.Essential Fatty Acids of Grass Carp Ctenopharyngodon idella[J].Nippon Suisan Gakkaishi,1991,57(3):467~473.
[24]Tocher D R.Metabolism and functions of lipids and fatty acids in teleosts fish[J].Rev.Fish.Sci.,2003,11,107~184.
[25]Tocher D R.Fatty acid requirements in ontogeny of marine and freshwater fish.Aquaculture Research,2010,41(5),717~732.
[26]Tocher D R.Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective[J].Aquaculture,2015,449:94~107.
[27]Wood J D,Enser M,Fisher A V,et al.Fat deposition,fatty acid composition and meat quality:A review[J].Meat science,2008,78(4):343~358.
[28]Yao J,Zhi M,Gao X,et al.Effect and the probable mechanisms of silibinin in regulating insulin resistance in the liver of rats with non-alcoholic fatty liver[J].Brazilian Journal of Medical and Biological Research,2013,46(3):270~277.■
This study was carried out to evaluate whether the silymarin supplementation could influent fatty acid composition in juvenile grass carp with dietary lipid levels increasing.A total of 360 fish with(27.43±0.17)g body weighet were fed six isonitrogenous and isocaloric diets in a factorial design containing 0,100 or 200 mg/kg silymarin(SM0,SM100,SM200)associated with either 4%or 8%lipid levels(Low lipid,LL;high lipid,HL;respectively)for 12 weeks in an indoor recircling rearing system.Each treatment was replicated four groups.Results showed that tissue fatty acid composition were significantly affected by dietary lipid levels(P<0.05),but weren’t influenced by silymarin supplementation(P>0.05).Compared with LL treatments,percent of 18∶2n-6 and 20∶4n-6(ARA)in muscle,18∶2n-6 and 18∶3n-3 in intraperitoneal fat(IPF)were significantly increased by 60.99%,22.72%,64.40%and 161.29%in HL treatments,while percent of DHA in IPF,18∶2n-6,ARA,EPA and DHA in hepatopancrease were decreased by 26.76%,44.68%,63.59%,49.63%and 74.51%,respectively(P<0.05).Both muscle and IPF(r=0.91,0.91)showed markedly higher correlation than hepatopancreas(r=0.59)(P<0.05).SM100 groups had 3.33%higher correlation in muscle than SM200 groups(r=0.93,r=0.90)(P< 0.05).Correlation in IPF was 10.34%higher in HL treatments than LL treatments(r=0.96,r=0.87),and for LL treatments,both SM100 and SM200 had significantly higher correlation in IPF than SM0(r=0.91,0.87,0.84)(P<0.05).Correlation in hepatopancreas in HL treatments was 44.74%lower than LL treatments(r=0.42,r=0.76)(P<0.05).Increasing dietary lipid levels resulted in different correlation of fatty acid profile between tissue and diet,which might be associated with the adapting or regulating mode of dietary lipid levels,and the utilization and transformation of fatty acid.Silymarin supplementation could enhance the correlation of fatty acid profile between IPF and lower lipid level diets.There were tissue specific correlation of fatty acid profile in tissue and diets for silymarin supplementation.
grass carp;dietary lipid levels;silymarin;fatty acid
10.15906/j.cnki.cn11-2975/s.20161407
S816.7
A
1004-3314(2016)14-0026-08
北京市科学技术研究院青年骨干计划(201420);陕西省水产健康生态养殖技术集成与示范(2014-TS-36)
猜你喜欢
食品与生物技术学报(2022年11期)2023-01-22
昆明医科大学学报(2022年1期)2022-02-28
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
化工设计通讯(2020年4期)2020-01-15
化工设计通讯(2020年3期)2020-01-14
意林·全彩Color(2019年9期)2019-10-17
科学与财富(2018年8期)2018-05-09
中成药(2018年1期)2018-02-02
科学与财富(2016年32期)2017-03-04
