
229份水稻品种及重要育种材料抗稻瘟病Pik位点基因型鉴定
2016-11-01 07:16陈子强田大刚梁廷敏陈在杰胡昌泉陈松彪
福建农业学报 2016年6期
陈子强,田大刚,梁廷敏,2,陈在杰,胡昌泉,王 锋,陈松彪*
(1.福建省农业科学院生物技术研究所/福建省农业遗传工程重点实验室,福建 福州 350003;2. 福建农林大学生命科学学院,福建 福州 350002)
229份水稻品种及重要育种材料抗稻瘟病Pik位点基因型鉴定
陈子强1,田大刚1,梁廷敏1,2,陈在杰1,胡昌泉1,王锋1,陈松彪1*
(1.福建省农业科学院生物技术研究所/福建省农业遗传工程重点实验室,福建福州350003;2. 福建农林大学生命科学学院,福建福州350002)
Pik位点包含Pi-k、Pi-km、Pi-kp、Pi1、Pi-kh等抗稻瘟病等位基因,在我国水稻抗病育种中具有重要应用价值。本研究对229份水稻品种及重要育种材料的Pik位点进行基因型鉴定。利用K/N-单元型分子标记对水稻材料Pik位点进行多态性分析,鉴定了80份材料为K1K2功能性基因型。进一步通过Pi-k、Pi-kp和Pi1功能分子标记检测,结合功能多态位点序列分析,从80份K1K2型材料中,鉴定了黑稻、京虎B为Pi-k基因型;软米谷为Pi-kp基因型;恩恢99-64为Pi1基因型。研究还发现,高配合力强优恢复系闽恢3301的Pik位点为功能性K1K2基因型,其K1K2片段的序列与已克隆Pi-k、Pi-km、Pi-kp、Pi1、Pi-kh等位基因的序列存在差异,表明闽恢3301的Pik位点可能存在一个新等位基因。本研究明确了229份水稻材料Pik位点的K/N-基因型,结果可为水稻抗稻瘟病育种和品种合理布局提供参考。
水稻;稻瘟病;Pik位点;闽恢3301;基因分型
稻瘟病是水稻生产上最严重病害之一,每年可造成水稻减产10%~30%[1]。长期生产实践表明,培育和推广抗病水稻品种是防治稻瘟病最经济有效、对环境安全的措施。因此,解析水稻品种及育种材料的抗病遗传基础,对开展水稻抗稻瘟病育种和合理布局抗病品种具有重要意义。
迄今为止,已有近百个稻瘟病抗性基因被人们鉴定,其中有23个基因已经被克隆[2]。随着越来越多的稻瘟病抗性基因被克隆,基于抗病基因功能序列开发的分子标记,促使人们能够快速、准确地鉴定各种水稻种质资源抗病基因的分布,进而指导水稻抗病分子育种[3-8]。
Pik位点位于水稻第11染色体长臂近端粒处,是水稻一个主效抗稻瘟病位点。该位点具有Pi-k、Pi-km、Pi-kp、Pi-kh、Pi1及Pi-ks等多个抗稻瘟病等位基因。这些基因对我国不同稻区的稻瘟菌生理小种均具有较广谱及较持久、高效抗性,因此,在我国水稻抗病育种中具有重要的应用价值[9-14]。Pi-km[9]、Pi-kp[11]、Pi-k[12]、Pi1[13]和Pi-kh[14]等基因已被克隆。研究表明,Pik位点等位基因介导的抗性由2个紧密连锁、具有独立功能的NBS-LRR基因协同作用[9-14]。对比抗病品种Kusabue(Pi-k)、Tsuyuake(Pi-km)、K60(Pi-kp)、C101LAC(Pi1)等Pik位点2个功能性NBS-LRR基因序列与感病品种日本晴非功能DNA序列发现,该位点可分为K-单元型(K1K2功能型)和N-单元型(N1N2非功能型)[12]。进一步研究发现,Pi-km、Pi-k、Pi-kp和Pi1基因型取决K1、K2区间的6个SNP,其中K1基因组C1-685A /G能区分Pi-km与其他等位基因,T1-2944G能区分Pi-k与其他等位基因,K1基因组T1-786A/G和K2基因组A2-1879G是Pi-kp特异的SNP位点,K1基因组T1-687G和K2基因组T2-2261A是Pi1特异的SNP位点[12-13]。基于这些多态开发的分子标记能够检测Pik位点的N/K-基因型并区分不同等位基因型。
本研究对229份中国水稻品种及重要育种材料的抗稻瘟病Pik位点进行K/N-单元型多态性分析及等位基因分型,明确了这些水稻材料Pik位点的K/N-基因型。研究结果将为水稻抗稻瘟病育种和品种合理布局提供参考。
1 材料与方法
1.1水稻材料
水稻Pik位点单基因系材料IRBLKM-TS(Pi-km)、IRBLk-Ka(Pi-k)、IRBLkp-K60(Pi-kp)、IRBL1-F5(Pi1)、IRBLks-S(Pi-ks)及IRBLkh-K3(Pi-kh)和Pib位点单基因系材料IRBLb-B(Pi-b)由国际水稻研究所Zhou Bo博士惠赠。229份水稻品种及重要育种材料以及参考基因组水稻品种日本晴由福建省农业科学院遗传工程重点实验室保存。
1.2水稻基因组DNA 的提取
取3~4叶期水稻植株的新鲜叶片组织100 mg,装入 2 mL离心管,用液氮研磨成粉末。采用CTAB法提取基因组DNA[15]。
1.3PCR扩增、酶切检测及SNP位点测序
PCR体系总体积为25 μL,其中2.5 μL的10×buffer,2.0 μL的2.5 mmol dNTP, 1.0 μL上游引物和1.0 μL下游引物, 1.0 μL的基因组DNA,0.5 μL的Taq聚合酶,ddH2O补至25 μL。PCR 反应程序为:94℃预变性3 min;94℃变性30 s,56℃退火30 s,72℃延伸1 kb·min-1,35个循环;72℃延伸5 min。用1.5%的琼脂糖凝胶电泳分离PCR产物。
Pi-k、Pikp-k2、Pi1-k1基因型检测采用dCAPS标记策略,分别以dCAPS-2953[12]、dCAPS-1879[12]和Pi1-5C[13]等3个标记的引物进行扩增。PCR产物在37℃条件下,分别用限制性内切酶KpnI、NdeI、ClaI进行酶切处理4 h。酶切产物用3%的琼脂糖凝胶电泳检测。
Pikp-k1和Pi1-k2基因型通过测序进行检测。PCR产物经试剂盒(TIANGEN公司,北京)回收纯化,由英骏生物技术公司(上海)进行测序。
2 结果与分析
2.1229份水稻材料抗稻瘟病Pi-k位点K/N-单元型多态分析
利用4对特异性引物Ku1-GT[12]、Ku2-GT[12]、NP5-GT[12]、NP6-GT[12](表1)对日本晴及携带Pi-b、Pi-k、Pi-km、Pi-kp、Pi-kh、Pi1和Pi-ks等位基因的单基因系进行单元型检测。利用N-型标记NP5-GT和NP6-GT可以从日本晴和IRBLb-B单基因系基因组分别检测637 bp和676 bp的片段,而6个单基因系则无扩增产物;相反,利用K-型标记Ku1-GT和Ku2-GT可以从6个单基因系基因组分别检测到1 600 bp和4 879 bp的片段,而日本晴和IRBLb-B单基因系则无扩增产物(图1,表2),验证了Pik位点存在K-单元型(K1K2功能型)和N-单元型(N1N2非功能型)的现象[12]。
进一步利用上述4对引物对229份水稻品种及重要育种材料(表2)进行检测,结果表明,117、02428、IR046、R187粳、安农晚粳B和黑竹粳等80份材料为K1K2基因型;D297B、R527、ZR21、白芒粳、光壳香糯和黑粳糯2号等53份材料为N1N2基因型;其余96份材料则为无规则基因型(图1,表2)。
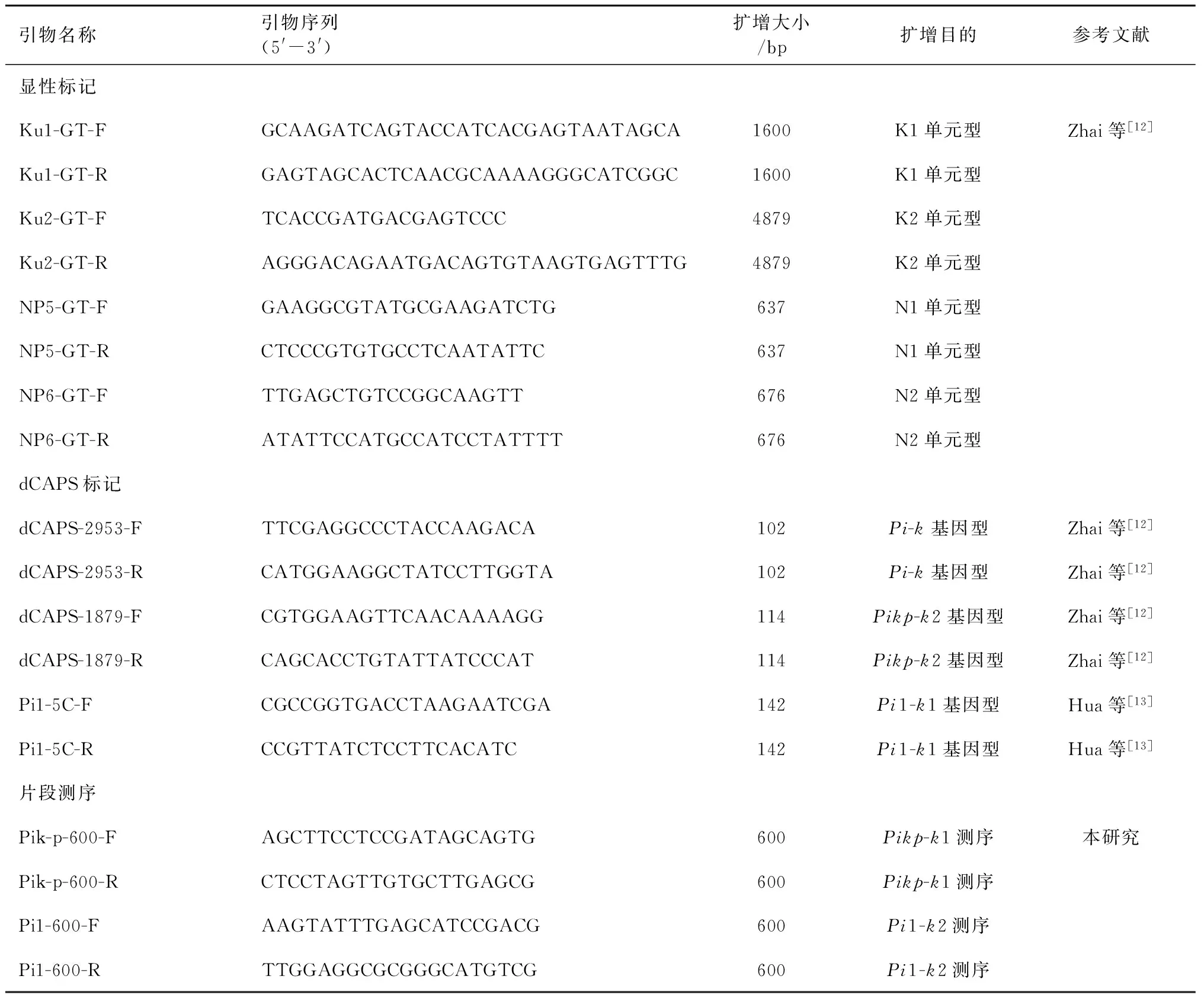
表1 供试引物
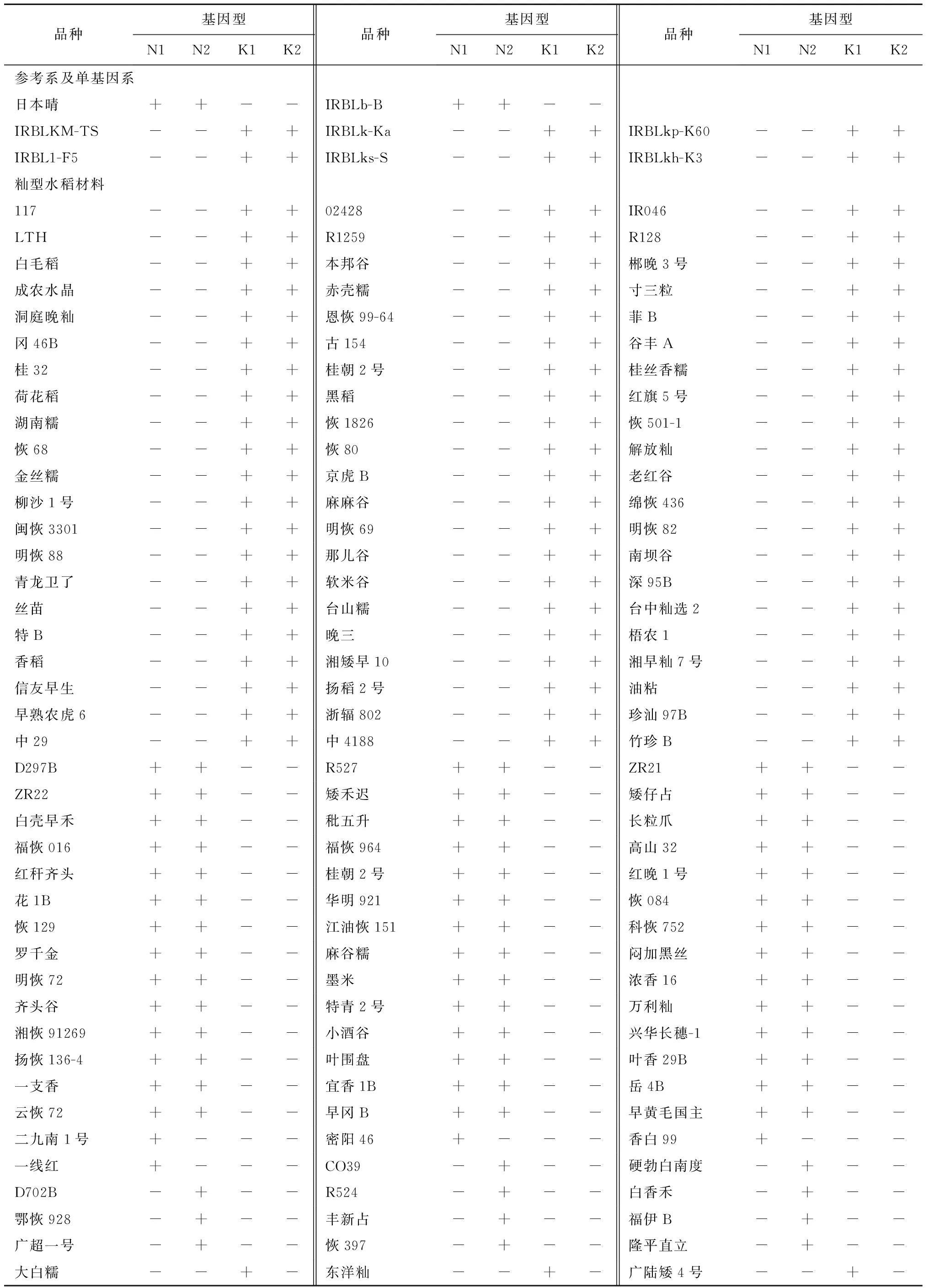
表2 229份水稻品种及重要育种材料Pik位点N/K-基因型检测
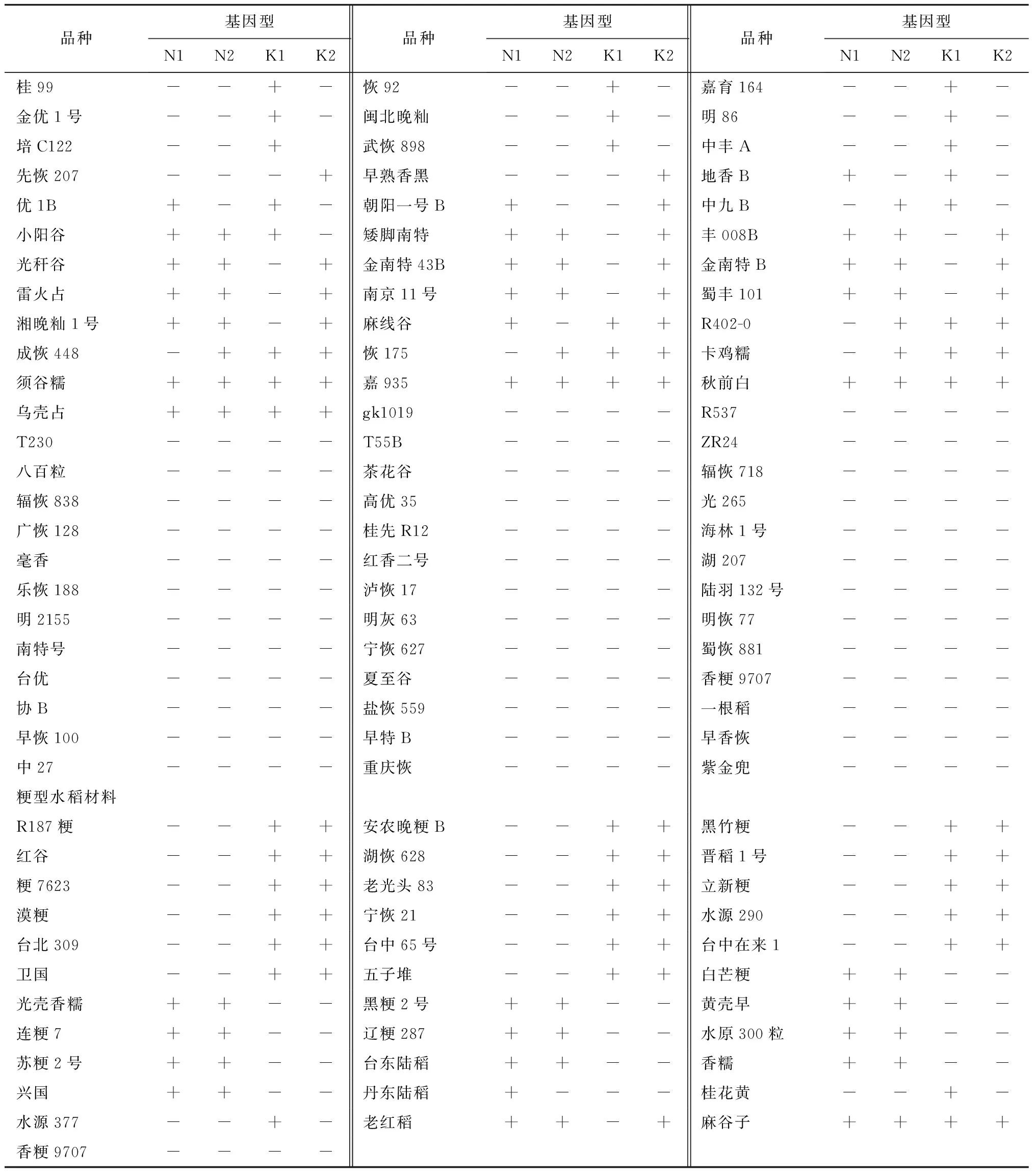
(续上表)
2.2Pik位点等位基因型鉴定
Zhai等[12]和Hua等[13]设计了针对Pi-km、Pi-k、Pi-kp、Pi1等位基因的dCAPS标记。本研究参考这些标记进行水稻材料检测,发现dCAPS-2953标记(对应Pi-k基因型)、dCAPS-1879标记(对应Pikp-k2基因型和Pikh-k2基因型)和Pi1-5C标记(对应Pi1-k1基因型)扩增、酶切效果稳定。以dCAPS-2953标记对80份K1K2型水稻材料进行检测,结果发现黑稻、京虎B等2份材料的基因型与IRBLk-Ka(Pi-k)一致;以dCAPS-1879标记进行分析,检测到软米谷在Pikp-k2位点上与IRBLkp-K60(Pi-kp)、IRBLkh-K3(Pi-kh)基因型一致;以Pi1-5C标记进行分析,检测到洞庭晚籼、彬晚3号、恩恢99-64等3份材料在Pi1-k1位点上与IRBL1-F5(Pi1)基因型一致(图2)。
为进一步分析软米谷在Pikp-k1位点上,洞庭晚籼、彬晚3号、恩恢99-64在Pi1-k2位点上的基因型,分别对软米谷和洞庭晚籼、彬晚3号、恩恢99-64的相应位点进行PCR扩增和序列测序。结果发现,软米谷在Pikp-k1位点上的基因型与IRBLkp-K60(Pi-kp)一致;恩恢99-64在Pi1-k2位点上的基因型与IRBL1-F5(Pi1)一致,洞庭晚籼、彬晚3号的Pi1-k2位点基因型则与IRBL1-F5(Pi1)存在变异(图3)。
2.3闽恢3301基因型分析
闽恢3301是福建省农业科学院生物技术研究所近年来选育的高配合力强优恢复系[16],以其为父本配制并审定了十多个超高产组合,并大面积应用于生产中。K/N-单元型多态检测的结果表明,闽恢3301的Pik位点为K1K2基因型(图1,表2)。进一步以dCAPS-2953、dCAPS-1879、Pi1-5C等三个标记对闽恢3301的Pik位点进行检测,发现其基因型不同于Pi-k、Pi-kp、Pi1。为明确闽恢3301的基因型,对闽恢3301Pik位点的K1和K2片段进行测序并与5个已克隆等位基因Pi-k、Pi-km、Pi-kp、Pi1和Pi-kh的序列进行比对,结果发现,闽恢3301的K1与Pikp-k1功能位点相同,但其K2不同于Pikp-k2或Pi1-k2功能位点。综合K1和K2位点,闽恢3301序列与5个等位基因的序列存在多态(图3),说明闽恢3301的Pik位点可能存在新的等位基因。
3 讨 论
目前已鉴定的水稻抗稻瘟病基因分布在除第3染色体外的其余11条染色体上,许多基因以基因簇形式分布在不同的染色体区域。其中,第6染色体的Pi2/9位点、第11染色体的Pik位点以及第12染色体的Pi-ta位点,均存在着数目众多的抗病等位基因[2]。Pik位点的多个等位基因均具有对中国不同稻区稻瘟菌生理小种较广谱和持久的抗性,如Wang等[10]鉴定发现,Pi-km、Pi-k、Pi-kp等基因对广东、湖南、四川、江苏等稻瘟菌株的抗性频率达到90%以上。另一方面,不同抗病基因对不同地区稻瘟菌株的抗性表现差异明显,如上述3个基因对东北稻区稻瘟菌株的抗性频率低于50%[10]。因此,了解水稻材料抗病位点基因型,根据不同稻区稻瘟病发病情况选择、选育携带不同抗病基因的抗病品种,对不同稻区稻瘟病防控具有重要指导意义。
基于K/N-单元型分子标记检测,本研究从229份水稻材料中,鉴定了80份材料为K1K2基因型,即这些材料可能在Pik位点含有抗稻瘟病功能基因。进一步以对应Pik功能位点的分子标记dCAPS-2953、对应Pikp-k2功能位点的分子标记dCAPS-1879以及对应Pi1-k1功能位点的分子标记Pi1-5C,结合Pik功能位点、Pikp-k1功能位点以及Pi1-k2功能位点序列分析,鉴定了黑稻、京虎B为Pi-k基因型,软米谷为Pi-kp基因型,恩恢99-64为Pi1基因型。Pik位点上已被克隆的抗病基因包括Pi-km、Pi-kp、Pi-k、Pi1和Pi-kh等基因。鉴于标记扩增稳定性问题,本研究未针对Pi-km和Pi-kh基因型进行分析,未来仍有待鉴定哪些K1K2型材料为Pi-km或Pi-kh基因型。
K/N-单元型分子标记分析结果表明,闽恢3301为K1K2基因型。进一步对闽恢3301 K1K2片段的序列分析结果表明,闽恢3301基因型不同于已克隆的Pi-k、Pi-km、Pi-kp、Pi1和Pi-kh等抗性基因(图3),说明闽恢3301的Pik位点可能存在一个新的等位基因,但其是否具有抗稻瘟病功能,需要进一步的遗传、抗病性实验予以证明。
总体上,本研究鉴定了229份中国水稻品种及重要育种材料抗稻瘟病Pik位点的K/N-基因型,研究结果对开展水稻抗稻瘟病分子育种以及合理利用抗病品种具有指导意义。
[1]DEAN R A,TALBOT N J,EBBOLE D J,et al. The genome sequence of the rice blast fungusMagnaporthegrisea[J]. Nature,2005,434(7036):980-986.
[2]LIU W,LIU J,Triplett L,et al. Novel insights into rice innate immunity against bacterial and fungal pathogens [J]. Annual Review Phytopathology,2014,52:213-241.
[3]张学堂,廖新华,朱振华,等.Pi-z、Pi-ta2等基因在云南粳稻稻瘟病抗性育种中的应用 [J]. 中国农业科技导报,2010,12,(1):100-105.
[4]张羽,冯志峰,崔明珠,等. 稻瘟病抗性基因Pi-km在陕西省稻种资源中的分布状况 [J]. 分子植物育种,2013,(3):311-316.
[5]冯慧,杨成明,吴孝波,等. 四川省杂交稻亲本及32份抗源抗稻瘟病基因的检测分析 [J]. 西南农业学报,2013,(3):987-993.
[6]王军,杨杰,朱金燕,等. 稻瘟病抗病基因Pi-kh功能标记的开发及江苏粳稻品种中Pi-kh的变异 [J]. 中国水稻科学,2014,(2):141-147.
[7]何重,陈涛,张亚东,等. 江苏部分粳稻品种和品系中稻瘟病抗性基因Pi-ta和Pi-b的基因型分析 [J]. 江苏农业学报,2014,(5):921-927.
[8]TIAN D,CHEN Z,CHEN Z,et al. Allele-specific marker-based assessment revealed that the rice blast resistance genesPi2 andPi9 have not been widely deployed in Chineseindicarice cultivars[J]. Rice,2016,9(1):19.
[9]Ashikawa I,Hayashi N,Yamane H,et al. Two adjacent nucleotide-binding site-leucine-rich repeat class genes are required to conferPikm-specific rice blast resistance [J]. Genetics,2008,180(4):2267-2276.
[10]WANG L,XU X,LIN F,et al. Characterization of rice blast resistance genes in thePikcluster and fine mapping of thePik-plocus[J]. Phytopathology,2009,99:900-905.
[11]YUAN B,ZHAI C,WANG W,et al. ThePik-p resistance toMagnaportheoryzaein rice is mediated by a pair of closely linked CC-NBS-LRR genes [J]. Theoretical and Applied Genetics,2011,122(5):1017-1028.
[12]ZHAI C,LIN F,DONG Z,et al. The isolation and characterization ofPik,a rice blast resistance gene which emerged after rice domestication [J]. New Phytologist,2011,189(1):321-334.
[13]HUA L,WU J,CHEN C,et al. The isolation ofPi1, an allele at thePiklocus which confers broad spectrum resistance to rice blast [J]. Theoretical and Applied Genetics,2012,125(5):1047-1055.
[14]ZHAI C,ZHANG Y,YAO N,et al. Function and interaction of the coupled genes responsible forPik-hencoded rice blast resistance [J]. PLoS One,2014,9(6):e98067.
[15]MURRAY M G,THOMPSON W F.Rapid isolation of high molecular weight plant DNA [J]. Nucleic Acids Research, 1980,8(19):4321-4325.
[16]吴明基,向海涛,刘华清,等. 利用形态学性状及SSR分子标记鉴定水稻优良恢复系闽恢3301的亲代来源 [J]. 福建农业学报,2012,27(7):673-678.
(责任编辑:柯文辉)
Characterization of the Genotypes at the Rice Blast ResistancePikLocus in 229 Rice Cultivars and Important Breeding Materials
CHEN Zi-qiang1, TIAN Da-gang1, LIANG Ting-min1,2, CHEN Zai-jie1, HU Chang-quan1,WANG Feng1, CHEN Song-biao1*
(1.BiotechnologyResearchInstitute,FujianKeyLaboratoryofGeneticEngineeringforAgriculture,FujianAcademyofAgriculturalSciences,Fuzhou,Fujian350003,China; 2.CollegeofLifeScience,FujianAgricultureandForestryUniversity,Fuzhou,Fujian350002,China)
The ricePiklocus that contains multiple blast resistance genes, such asPi-k,Pi-km,Pi-kp,Pi1 andPi-kh, has great utilization value in rice breeding for blast resistance in China. In the present study, we investigated the genotypes at thePiklocus in 229 rice cultivars and important breeding materials. Based on PCR amplification using K- and N-type specific primer sets, 80 accessions were identified harboring the K1K2 haplotypes. The 80 K1K2 accessions were further accessed for the presence ofPi-k,Pi-kpandPi1 using allele-specific markers in combination with sequencing identification of allele-specific SNPs. Heidao and Jinghu B were identified harboring thePi-kresistance allele, and Ruanmigu and Enhui 99-64 were identified harboringPi-kpandPi1, respectively. In addition, an eliteindicarestorer line Minhui 3301 was identified harboring the K1K2 haplotypes. Allele-specific marker analysis and sequencing data revealed that the sequence of Minhui 3301 atPiklocus differs from the sequences ofPi-k,Pi-km,Pi-kp,Pi1 andPi-kh, suggesting that Minhui 3301 might possess a new allelic gene at thePiklocus. In summary, we validated the distribution of the K1K2 haplotypes at thePikloucs in 229 rice accessions. Our results provide a wealthy of information for developing and deploying resistant cultivars to control rice blast disease.
rice; rice blast;Piklocus; Minhui 3301; genotyping
CHEN Z-Q,TIAN D-G,LIANG T-M,et al.Characterization of the Genotypes at the Rice Blast ResistancePikLocus in 229 Rice Cultivars and Important Breeding Materials[J].FujianJournalofAgriculturalSciences,2016,31(6):553-559.
2016-03-12初稿;2016-05-14修改稿
陈子强(1988-),男,硕士,研究实习员,主要从事植物分子生物学与分子免疫研究
陈松彪(1973-),男,博士,研究员,主要从事植物分子免疫及植物功能基因研究(E-mail: songbiao_chen@hotmail.com)
福建省自然科学基金杰出青年基金滚动项目(2014J07004);国家自然科学基金委员会-福建省人民政府促进海峡两岸科技合作联合基金重点支持项目(U1405212);福建省科技计划项目——省属公益类科研院所基本科研专项(2014R1019-10)
Q 78
A
1008-0384(2016)06-553-07
陈子强,田大刚,梁廷敏,等.229份水稻品种及重要育种材料抗稻瘟病Pik位点基因型鉴定[J].福建农业学报,2016,31(6):.553-559
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
今日农业(2022年4期)2022-06-01
作物学报(2022年6期)2022-04-08
智慧健康(2021年17期)2021-07-30
农民致富之友(2020年15期)2020-05-25
农民致富之友(2019年24期)2019-08-20
发明与创新·大科技(2019年5期)2019-07-31
新课程·下旬(2018年9期)2018-11-14
青少年科技博览(中学版)(2015年10期)2015-01-11
中学生物学(2008年6期)2008-08-29
