
水稻Kasalath改良品系基因转化效率的初步研究
2016-10-20 04:29何行健曾崇华刘清波陈芬胡文彬王作平肖国樱
现代农业科技 2016年5期
关键词:籼稻
何行健 曾崇华 刘清波 陈芬 胡文彬 王作平 肖国樱

摘要 以亲本Kasalath和HD9802S为对照,Bar基因为筛选标记,初步评价了Kasalath改良品系9K08-3、9K18-7、9K21-3、9K28-5、9K29-9和9K30-0的基因转化效率。试验共获得15个独立转化事件,9K08-3、9K18-7、9K21-3、9K28-5、9K29-9和9K30-0的基因转化效率分别为0、0.65%、0、0.23%、0.40%和0.22%;初步结果表明改良品系9K18-7具有较高的基因转化效率。
关键词 籼稻;基因转化效率;抗除草剂
中图分类号 S511 文献标识码 A 文章编号 1007-5739(2016)05-0010-03
Abstract The transformation efficiencies of several improved lines from Kasalath(including 9K08-3,9K18-7,9K21-3,9K28-5,9K29-9,and 9K30-0)were preliminary evaluated by using parent Kasalath and HD9802S as check and herbicide resistant gene Bar as selective marker.Fifteen independent transformants were obtained in this study,and the transformation efficiencies of line 9K08-3,9K18-7,9K21-3,9K28-5,9K29-9,and 9K30-0 were 0,0.65%,0,0.23%,0.40% and 0.22%,respectively.It primary indicated that line 9K18-7 had higher transformation efficiency comparatively.
Key words Indica rice;transformation efficiency;herbicide resistance
水稻(Oryza sativa L.)是最重要的糧食作物之一,全世界近50%的人口以稻米为主食,90%的水稻产于亚洲。亚洲栽培稻一般分为籼稻和粳稻2个亚种,其中籼稻约占全球水稻产量的80%[1]。水稻在组织培养力和基因转化效率方面对基因型依赖性强,不同基因型水稻的组织织培养力和遗传转化效率存在显著差异。由于大多数籼稻不良的组织培养特性和极低的转化效率,严重限制了转基因技术在籼稻育种上的应用[2]。研究发现Kasalath是籼稻中易于组织培养和遗传转化的品种[3-4],但是Kasalath农艺性状较差,主要表现为高秆、有芒、株型松散、产量较低、生育期长等,严重制约了它在转基因育种研究上的广泛应用。水稻转基因育种研究中亟需转化效率高、农艺性状好的籼稻品系。
早籼型温敏核不育系HD9802S熟期早、农艺性状优良[5]。本课题组从HD9802S/Kasalath后代中选出了组织培养力高、矮秆、生育期短、产量性状较好的Kasalath改良品系[6],但高培养力品系的基因转化效率尚未检测。本文报道了以农杆菌介导转化法评价Kasalath改良品系基因转化效率的初步结果。
1 材料与方法
1.1 水稻材料
Kasalath改良品系9K08-3、9K18-7、9K21-3、9K28-5、9K29-9和9K30-0由本课题组培育[6];籼稻品种HD9802S由湖北大学提供;Kasalath由本所提供、本课题组繁殖。
1.2 组织培养和基因转化
组织培养方法参考文献[6],基因转化方法参考文献[7]。根瘤农杆菌菌株EHA105由本课题组购买、保存;质粒pCAMBIA3300由澳大利亚国际农业分子生物学应用中心惠赠,本课题组扩增、保存。植物组织培养力(%)=愈伤组织诱导率×绿苗分化率×100;基因转化效率(%)=转化事件数/试验种子数×100。
1.3 转基因植株检测
PCR和Southern检测方法参考文献[8-9]。转基因植株的除草剂抗性检测:从T1代植株中选取健康叶片以1 g/L的草铵膦溶液涂抹叶上部标记部位,以受体材料为对照,7 d后观察。
1.4 统计学分析
用Excel 2013和SPSS 21.0软件对数据进行统计分析。
2 结果与分析
2.1 转化事件的鉴定
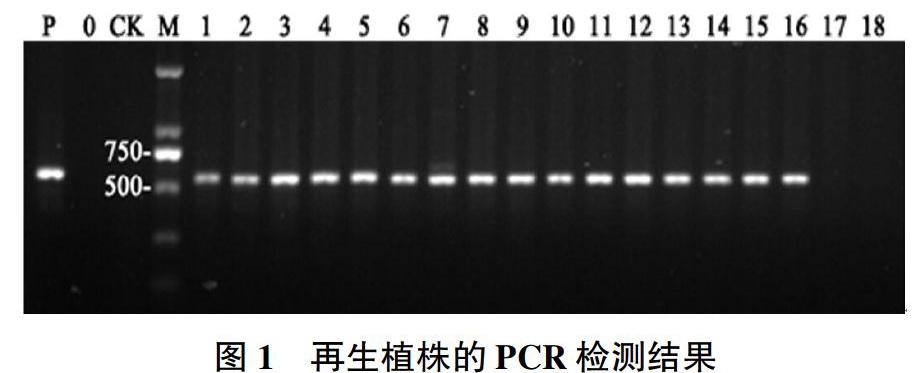
2.1.1 PCR鉴定。转化后获得的再生植株经炼苗移栽共成活18株,其中9株来源于Kasalath、2株来源于9K18-7、2株来源于9K28-5、3株来源于9K29-9、2株来源于9K30-0。对18株再生苗进行PCR检测,结果表明,除了9K28-5和9K30-0中各有1株没有检测到Bar基因外,其余16株再生苗均能检测到目的基因(图1)。
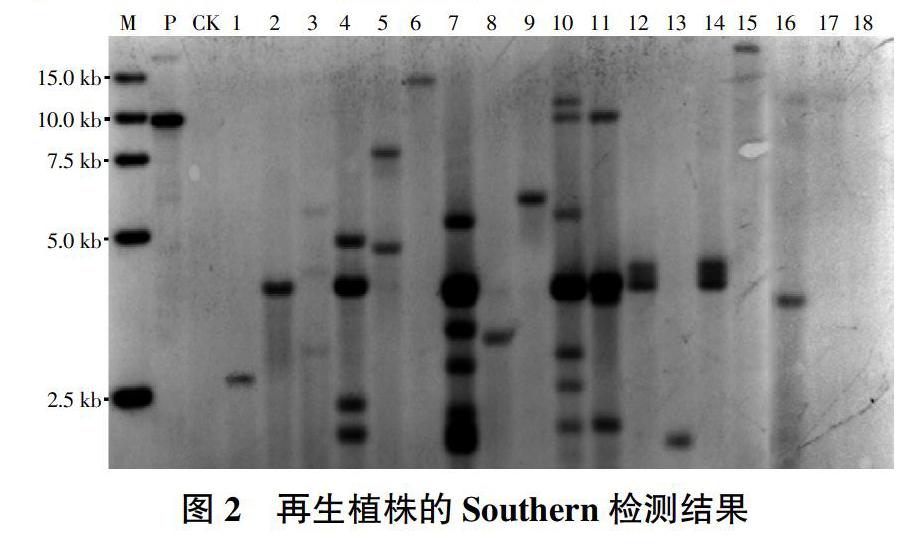
2.1.2 Southern鉴定。再生植株基因组DNA经酶切后电泳、转膜,以Bar基因片断为探针进行杂交,除PCR鉴定为假阳性的2株外,其余16株均得到了杂交信号(图2)。其中来源于Kasalath的有5株为单拷贝(图2中泳道1、2、6、8和16)、4株为多拷贝(图2中泳道3、4、5和7);来源于9K18-7的有1株为单拷贝(图2中泳道9)、1株为多拷贝(图2中泳道10);来源于9K28-5的1株为多拷贝(图2中泳道11),1株无杂交信号(图2中泳道17);来源于9K29-9的有1株为单拷贝(图2中泳道13)、2株为多拷贝且为同一转化事件(图2中泳道12、14);来源于9K30-0的1株为单拷贝(图2中泳道15),1株无杂交信号(图2中泳道18)。结果表明本试验共获得15个独立转化事件。
2.1.3 草銨膦抗性鉴定。除草剂草铵膦涂抹叶片试验发现,非转基因植株叶片明显变黄(CK),假阳性的再生植株表现与对照相似(11和12号),其余再生植株叶片伤害较弱或无明显伤害(图3)。结果表明分子检测为阳性的再生植株对草铵膦有明显的抗性。
2.2 基因转化效率的评价
研究结果表明(表1):亲本Kasalath和HD9802S的组织培养力分别为28.17%和24.36%;改良品系9K18-7和9K28-5的组织培养力分别为38.96%和46.80%,高于2个亲本;9K30-0和9K21-3分别为24.79%和25.24%,介于2个亲本之间;9K08-3和9K29-9分别为4.10%和22.97%,低于2个亲本。Kasalath和HD9802S的基因转化效率分别为4.21%和0;改良品系9K08-3、9K18-7、9K21-3、9K28-5、9K29-9和9K30-0的基因转化效率分别为0、0.65%、0、0.23%、0.40%和0.22%。改良品系9K18-7具有较高的基因转化效率,达到0.65%,基因转化效率介于高值亲本Kasalath和低值亲本HD9802S之间。
进一步分析了Kasalath改良品系中基因转化效率与组织培养特性之间的相关性。结果表明(表2):基因转化效率与愈伤组织诱导率、绿苗分化率和成熟胚培养力之间的Pearson相关系数分别为0.692、0.375和0.547,相关性均不显著;成熟胚培养力与愈伤组织诱导率和绿苗分化率之间的Pearson相关系数分别为0.871和0.966,相关性分别达到显著和极显著水平;绿苗分化率和愈伤组织诱导率之间的Pearson相关系数为0.754,相关性不显著。
3 结论与讨论
转基因技术是一种快速、定向改良水稻的有效手段。自从1994年Hiei等[10]建立粳稻的农杆菌介导法高效转化体系以来,粳稻的遗传转化工作迅速展开,取得了很多进展[11-16]。由于基因型不同,籼稻的基因转化效率不高,研究进展较慢[17],且主要集中于转化体系的优化上。Rashid等[18]通过调整乙酰丁香酮用量、愈伤组织类型和共培养时间等因素,改变激素配比,建立了一种高效转化中籼型水稻品种印度香米385的体系;Hiei等[19]研究了不同凝胶剂和共培养基对籼稻转化效率的影响;叶松青等[20]主要针对籼稻转化过程中涉及的愈伤组织的诱导、筛选、分化等进行了研究,还研究了脯氨酸、二甲基亚砜对分化的影响。但是,籼稻转基因受体品种的改良方面研究较少。马炳田[21]从优良三系杂交稻恢复系(SH527和SH163)、保持系(D62B、D297B和D702B)、两系恢复系W2008、两系不育系612S等材料中筛选,并建立了一套转基因中间受体系统。籼稻转基因优良受体品种的缺乏,在一定程度上制约了籼稻转基因研究的发展。
本课题组从受体品种改良方面入手,利用培养力较好且易于转化的籼稻品种Kasalath与农艺性状较好、培养力高的不育系HD9802S杂交,结合分子标记方法,得到了291个优良品系,并筛选出培养力极显著高于亲本HD9802S和Kasalath的家系9K18和9K19,为水稻转基因育种研究提供了较好的高培养力受体材料[6]。
水稻的组织培养和转化是一个极为复杂的过程,在此过程中,组织培养特性被认为与转化效率密切相关。李素娟等[22]通过16个籼稻品种和5个粳稻品种的遗传转化试验,认为较好的组织培养力是获得转基因植株的前提。樊秀霞[23]通过对21个水稻品种的成熟胚组织培养特性以及转基因效率的考察,发现这些水稻品种间组织培养特性和转基因效率均存在显著差异,认为水稻品种间组织培养特性和转基因效率有关。Zhao等[2]认为一个水稻品种的遗传转化效率在很大程度上取决于它的组织培养力。但他们均没有对组织培养力和转化效率之间的关系进行系统研究,这可能与籼稻遗传转化的低成功率和难重复性有关。
本试验对6个Kasalath改良品系进行遗传转化,获得了6个独立转化体,相关分析表明Kasalath改良品系的基因转化效率和组织培养特性之间的相关性并不显著。Kasalath和HD9802S的组织培养力并没有显著差异[6],但本研究中HD9802S的转化效率为0,Kasalath的基因转化效率4.21%。虽然刘风珍等[24]在花生的遗传转化研究上也获得了类似结果,但由于本试验获得的转化事件数少,还不具有普遍意义。因此,在后续筛选试验中,一方面要加大试验工作量,探索基因转化效率和组织培养特性之间的内在关系,另一方面也要以基因转化效率为最终衡量标准,筛选组织培养力和基因转化效率都高的改良品系,为籼稻的转基因育种研究提供理想的受体材料。
4 参考文献
[1] J L MACLEAN.水稻知识大全[M].杨仁崔,汤圣祥,译.福州:福建科学技术出版社,2003.
[2] ZHAO L,ZHOU H,LU L,et al.Identification of quantitative trait loci controlling rice mature seed culturability using chromosomal segment substitution lines [J].Plant Cell Reports,2009,28(2):247-256.
[3] YUKOH H,TOSHIHIKO K.Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed[J].Nature Protocols,2008,3(3):824-834.
[4] SAIKA H,TOKI S.Mature seed-derived callus of the model indica rice variety Kasalath is highly competent in Agrobacterium-mediated transfo-rmation[J].Plant Cell Reports,2010,29(12):1351-1364.
[5] 周勇,居超明,徐国成,等.优质早籼型水稻温敏核不育系HD9802S的选育与应用[J].杂交水稻,2008(2):7-10.
[6] 曾崇华.HD9802S/Kasalath选系中高培养力和抗稻瘟病株系的筛选[D].长沙:中国科学院,2015.
[7] SEIICHI T,NAHO H,KAZUKO O,et al.Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice[J].The Plant Journal,2006,47(6):969-976.
[8] 邓力华,邓晓湘,魏岁军,等.抗虫抗除草剂转基因水稻B1C893的获得与鉴定[J].杂交水稻,2014,29(1):67-71.
[9] 肖国樱,唐俐,袁定阳,等.转Bar基因抗除草剂两系杂交早稻恢复系Bar 68-1的培育研究[J].杂交水稻,2007,22(6):57-61.
[10] HIEI Y,OHTA S,KOMARI T,et al.Efficient transformation of rice(Oryza sativa L.)mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA [J].The Plant Journal,1994,6(2):271-282.
[11] 铁韦韦.农杆菌介导的籼粳稻转化差异[D].武汉:华中农业大学,2012.
[12] 杨友才,周清明.转基因水稻研究进展[J].湖南农业大学学报(自然科学版),2003,29(1):85-88.
[13] 朱祯.转基因水稻研发进展[J].中国农业科技导报,2010,8(2):9-16.
[14] 陈文岳.转Bt基因水稻“克螟稻”的遗传改良研究[D].杭州:浙江大学,2005.
[15] 沈圣泉,吴殿星,崔海瑞,等.Bt转基因抗虫粳稻品种选育中若干问题研究[J].浙江大学学报(农业与生命科学版),2003,29(5):31-35.
[16] 王利润.抗螟虫粳稻新材料的创建和抗螟虫两系杂交粳稻新组合的选育[D].武汉:华中农业大学,2012.
[17] KARABI D,KUMAR D S.Indica rice(Oryza sativa,BR29 and IR64)[J].Methods in Molecular Biology,2006,343(343):201-212.
[18] RASHID H,YOKOI S,TORIYAMA K,HINATA K.Transgenic plant pro-duction mediated by Agrobacterium in Indica rice[J].Plant Cell Reports,1996,15(10):727-730.
[19] HIEI Y,KOMARI T.Improved protocols for transformation of indica rice mediated by Agrobacterium tumefaciens[J].Plant Cell,Tissue and Organ Culture,2006,85(3):271-283.
[20] 叶松青,储成才,曹守云,等.提高水稻转化效率几个主要因素的研究[J].遗传学报,2001,28(10):933-938.
[21] 马炳田.基因工程改良杂交籼稻的抗虫性[D].雅安:四川农业大学,2002.
[22] 李素娟,樊秀霞,王华,等.水稻不同品种组培再生和转基因频率研究[J].核农学报,2013,27(12):1817-1827.
[23] 樊秀霞.水稻不同品種组培再生和转基因频率研究[D].杭州:杭州师范大学,2013.
[24] 刘风珍,万勇善,潘昱名.花生遗传转化高效基因型的筛选与研究[J].花生学报,2009,38(4):21-25.
猜你喜欢
安徽农业科学(2022年12期)2022-07-06
安徽农学通报(2022年8期)2022-05-06
意林·少年版(2018年11期)2018-07-04
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
原生态民族文化学刊(2015年1期)2015-05-05
绿色科技(2014年12期)2015-01-27
江苏农业科学(2014年11期)2015-01-15
世界热带农业信息(2014年3期)2014-08-12
原生态民族文化学刊(2014年1期)2014-08-08
