
猪瘟病毒C株特异的TCR Vα5和Vβ6真核表达载体的构建及共转染研究
2016-10-20 02:20王春燕陈国华何小兵贾怀杰房永祥李守杰景志忠
动物医学进展 2016年10期
王春燕,陈国华,何小兵,曾 爽,贾怀杰,房永祥,冯 园,李守杰,景志忠
(中国农业科学院兰州兽医研究所,家畜疫病病原生物学国家重点实验室,农业部兽医公共卫生重点实验室,甘肃兰州 730046)
猪瘟病毒C株特异的TCR Vα5和Vβ6真核表达载体的构建及共转染研究
王春燕,陈国华,何小兵,曾爽,贾怀杰,房永祥,冯园,李守杰,景志忠*
(中国农业科学院兰州兽医研究所,家畜疫病病原生物学国家重点实验室,农业部兽医公共卫生重点实验室,甘肃兰州 730046)
本实验室前期研究发现猪瘟病毒C株特异的TCR Vα5和TCR Vβ6基因家族,为进一步研究其体外表达情况,应用RT-PCR从猪外周血单个核细胞中扩增其全长基因序列,并构建TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP重组质粒,将其转染293T细胞。结果显示,TCR Vα5和TCR Vβ6全长基因分别为816 bp和945 bp,双酶切和核酸序列测定证实TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP重组质粒构建正确;转染293T细胞24 h后荧光显微镜下可以看到70%以上的细胞表达EGFP蛋白。此外,实时荧光定量PCR可检测到TCR Vα5和TCR Vβ6基因的表达,单独转染组和共同转染组Western blot均可检测到EGFP蛋白的表达,且蛋白杂交带强度相似。研究结果表明,CSFV-C株特异性的TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP真核表达质粒可同时转染到293T细胞中,实现体外共表达。
猪瘟病毒C株;TCR Vα/Vβ基因;脂质体转染;增强绿色荧光蛋白
猪瘟(Classical swine fever,CSF)是由猪瘟病毒(Classical swine fever virus,CSFV)引起的对养猪业危害严重的一种高度接触性传染病[1],以猪的高热稽留、白细胞减少、出血梗死和高病死率为特征,被世界动物卫生组织(OIE)列为必须报告的动物疫病,其大范围的暴发给我国和世界养猪业造成了巨大的经济损失[2-4]。
猪瘟病毒兔化弱毒疫苗(CSFV-C株弱毒苗,简称C株苗)在包括中国在内的全球猪瘟免疫预防中发挥了关键作用,其免疫效果得到了世界的认可,但其产生有效保护力的免疫学机制还不完全清楚。多个研究发现,C株弱毒苗免疫后既可产生中和抗体,又可诱导T细胞应答。免疫接种后7 d甚至更早便可以抵抗强毒攻击[5-7],但2周~3周才可检测到中和抗体[8],免疫接种后9 d能检测到诱导IFN-γ产生的病毒特异的T细胞,且这些特异的T细胞在体外具有直接的裂解靶细胞的效应[9]。这说明猪瘟病毒C株弱毒苗在中和抗体出现之前即可产生免疫保护力,推测细胞免疫应答在猪瘟病毒的早期免疫保护中可能扮演着重要角色[8,10]。因此,探究C株弱毒苗产生快速保护力的免疫学效应机制将有助于新一代高效疫苗的研发。
T细胞免疫是机体细胞免疫应答的重要组成部分,其中αβ T细胞是获得性免疫最关键的细胞,其表面的标志性异源二聚体,即αβ TCR是与抗原递呈细胞(APC)上的Ag-MHC分子识别、结合以及被激活的重要信号转导分子,决定着T细胞免疫识别、免疫应答的多样性和特异性,其中TCR α、β链的互补决定区3(CDR3)共同决定了T细胞识别抗原的特异性[11]。在机体正常情况下,αβ TCR CDR3的长度、序列组成呈高度多态性和多样性,具有识别和结合各种各样抗原表位肽的潜能,以应对复杂的环境。然而,在病毒感染时,只有某种TCR发生反应性重排,分化形成识别该病毒抗原特异性TCR的T细胞,并大量增殖,这种αβ TCR具有相同的或高度相似的CDR3序列和长度,具有特异性识别同类抗原的能力[12]。因此,通过分析病毒抗原刺激T细胞的TCR基因的CDR3长度和序列,就可以确定抗原特异的TCR,通过转导病毒抗原特异的TCR基因至正常T细胞中,使其强制表达这种独特型TCR,改变T细胞内源性TCR识别抗原的原有模式,经大量扩增后,具备分泌细胞因子的能力或特异性杀伤靶细胞的功能,完成机体T细胞对抗原的免疫应答。目前在医学上已有对肿瘤、细菌和病毒抗原特异性TCR基因的研究,以期应用于免疫治疗研究中[13-14]。然而,在兽医学中,这方面的研究较少,尚未有公开的报道。
本实验室前期研究发现,猪瘟病毒C株疫苗病毒感染能引起猪的某些TCR Vα和Vβ亚家族发生单克隆或寡克隆扩增[15]。本研究将在此基础上克隆其特异的Vα5和Vβ6亚家族的全长αβ TCR,构建CSFV-C株特异的TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP真核表达载体。由于293T细胞转染效率高,常用作外源基因转染的理想细胞,且其不表达TCR,不会产生错配现象,因此我们将CSFV-C株特异的TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP真核表达载体转染至293T细胞中,从mRNA和蛋白水平检测其转录与表达情况,为进一步在猪源T细胞上研究其免疫学功能奠定基础。
1 材料与方法
1.1材料
Trizol,美国Invitrogen公司产品;PrimescriptTM 1st strand cDNA Synthesis kit、LATaq酶、T4 DNA连接酶,宝生物工程(大连)有限公司产品;DNA片段凝胶回收试剂盒、质粒提取试剂盒,Axygen公司产品;pGEM-T easy载体,Promega公司产品;感受态大肠埃希菌DH5α,北京全式金生物技术有限公司产品;XholⅠ和SalⅠ限制性内切酶、SYBRHT5SS〗○RPremix ExTaqTM Ⅱ试剂盒,宝生物工程(大连)有限公司产品;小鼠抗人GAPDH单抗、鼠抗人EGFP单抗、兔抗小鼠的IgG二抗,Sigma公司产品;LipofectamineHT5SS〗○R3000转染试剂,美国Invitrogen公司产品;293T细胞、pIRES2-EGFP质粒,中国农业科学院兰州兽医研究所家畜疫病病原生物学国家重点实验室保存。
1.2方法
1.2.1CSFV-C株相关抗原特异TCR基因的全长克隆在前期研究中,利用RT-PCR和基因扫描-谱型分析技术分析猪瘟病毒C株免疫后猪外周血单个核细胞19个Vα和20个Vβ亚家族的表达情况和T细胞的克隆性,发现了Vα5和Vβ6亚家族呈单/寡克隆性增殖趋势,根据TCR Vα5和TCR Vβ6家族的部分已知序列,利用Primer5.0软件设计用于扩增TCR Vα5亚家族基因全长的引物,上游:5′-ATGAAGACATCCATTGGACCTT-3′,下游:5′-TCAGCTGGACCACAGCCG-3′;用于扩增TCR Vβ6亚家族基因全长的引物,上游:5′-ATGATCTCTGGTTTTCTCTGCTGG-3′,下游:5′-TCAGGCACCCTTTCTCTTGACCGT-3′ 。PCR总反应体积为25 μL:灭菌去离子水17.45 μL,10×PCR buffer 2.5 μL,2.5 mmol/L dNTP Mixture 2 μL,cDNA 2μL,上、下游引物(25 μmol/L)各0.4 μL,rTaq酶0.25 μL。PCR扩增条件:95 ℃ 5 min;94 ℃ 50 s,65 ℃ 15 s,72 ℃ 40 s,35个循环;72 ℃ 5 min。利用凝胶DNA回收试剂盒回收TCR Vα5和TCR Vβ6扩增的目的产物,与pGEM-T easy载体于16 ℃水浴连接过夜。将连接产物转化感受态大肠埃希菌DH5α,氨苄青霉素抗性筛选阳性菌落,经PCR、酶切鉴定后,将阳性克隆送至北京金唯智生物科技有限公司测序。
1.2.2全长基因的序列测定及分析 利用DNA Star、Signal 4.0 Sever-prediction 软件对测序正确的TCR Vα5和V β6全长基因序列进行结构预测分析。
1.2.3真核表达载体引物设计与PCR扩增根据测序正确的TCR Vα5和TCR Vβ6亚家族的全长基因序列,设计包括信号肽的表达引物并在其上、下游各加入酶切位点,TCR Vα5的引物,上游:5′-GC-CTCGAGGCCACCATGAAGACATCCATTGGA
CCTT-3′(下划线为XholⅠ位点),下游:5′-CCGTCGACTCAGCTGGACCACAGCCG-3′(下划线为SalⅠ位点);TCR Vβ6的引物,上游:5′-GC-CTCGAGGCCACCATGATCTCTGGTTTTCTCT
GCTGG-3′(下划线为XholⅠ位点),下游:5′ -CCGTCGACTCAGGCACCCTTTCTCTTGACCGT-3′(下划线为SalⅠ位点)。PCR总反应体积为25 μL:灭菌去离子水18.45 μL,10×PCR buffer 2.5 μL,2.5 mmol/L dNTP Mixture 2 μL,TCR Vα5-pGEM-T easy和TCR Vβ6-pGEM-T easy载体质粒1 μL为模板,带有酶切位点的上、下游引物(25 μmol/L)各0.4 μL,rTaq酶0.25 μL。PCR扩增条件:95 ℃ 5 min;94 ℃ 50 s,65 ℃ 15 s,72 ℃ 40 s,35个循环;72 ℃5 min。
1.2.4真核表达载体的构建与鉴定将纯化后的TCR Vα5和TCR Vβ6的PCR产物与pIRES2-EGFP载体质粒分别用XholⅠ和SalⅠ双酶切,纯化回收,用T4 DNA连接酶16 ℃连接过夜,转化DH5α感受态细胞。挑选菌落培养扩增并提取质粒,经PCR和双酶切鉴定后,将阳性克隆送北京金唯智生物技术有限公司测序,将测序正确的重组质粒命名为TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP。
1.2.5脂质体转染方法
1.2.5.1细胞培养将293T细胞以5×105个细胞接种于50 mL 25 cm2的培养瓶,于37 ℃、体积分数为5%的CO2培养箱内培养至细胞达80%瓶壁面积时收集细胞,以4×105个细胞/孔的密度接种于6孔板,置于37 ℃、体积分数为5%的CO2培养箱内培养至贴壁细胞达70% ~ 80%。
1.2.5.2转染提取阳性质粒和对照质粒,紫外分光光度计测定质粒浓度。试验共分5组:转染TCR Vα5-pIRES2-EGFP 质粒组(A组),转染TCR Vβ6-pIRES2-EGFP质粒组(B组),TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP 质粒共转染组(C组),转染空载体组(D组),不加任何DNA的未转染组(E组),转染方法参照Lipofectamine 3000 试剂说明书,在6孔板中进行转染。
1.2.6TCR Vα5和TCR Vβ6重组质粒转染293T细胞后的表达与鉴定
1.2.6.1荧光显微镜观察 利用荧光显微镜于转染后24 h,观察转染细胞的绿色荧光产生情况。
1.2.6.2RT-PCR和real-time PCR检测TCR Vα5和TCR Vβ6基因在mRNA水平的表达转染后48 h收集每个转染组细胞各2×106个,Trizol法提取总RNA,反转录成cDNA,用相应的TCR Vα5和TCR Vβ6亚家族引物进行PCR扩增, PCR扩增体系和扩增条件同1.2.1。应用SYBR Premix ExTaqⅡ试剂盒,SYBR Green荧光染料法检测TCR Vα5和TCR Vβ6基因的表达情况,并以同比例稀释的RNA作为内对照。总反应体积为20 μL,包括10 μL SYBR Premix ExTaqⅡ,6.0 μL 灭菌水,2.0 μL模板,上、下游引物(10 μmol/L)各0.8 μL,0.4 μL ROX Rerference Dye Ⅱ。扩增参数为:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 34 s,40个循环;95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。使用MX3005P荧光定量PCR仪进行扩增。
1.2.6.3Western blot检测重组质粒转染细胞后EGFP蛋白的表达情况转染后48 h裂解细胞提取总蛋白,同时以转染pIRES2-EGFP空载体组的细胞蛋白作为阳性对照,以未转染组的细胞蛋白作为阴性对照,以GAPDH蛋白为内参检测EGFP蛋白的表达。取15 μL蛋白样品和5 μL 4×SDS-PAGE Loading buffer进行120 g/L的SDS-PAGE电泳,电泳完成后转NC膜, 用小鼠抗人的GAPDH单抗和兔抗小鼠的IgG二抗进行孵育,底物显色检测GAPDH蛋白(内参照)的表达,以明确所提取蛋白的质量;同时,用鼠抗人的EGFP单抗和兔抗鼠的IgG抗体进行孵育,底物显色检测EGFP蛋白的表达,从而推测目的分子是否表达。
2 结果
2.1CSFV-C株抗原特异性猪αβ TCR全长基因的克隆
利用设计的特异性引物,成功扩增了CSFV-C株抗原特异性的TCR Vα5和TCR Vβ6亚家族全长基因序列,长度分别为800 bp和900 bp左右(图1)。
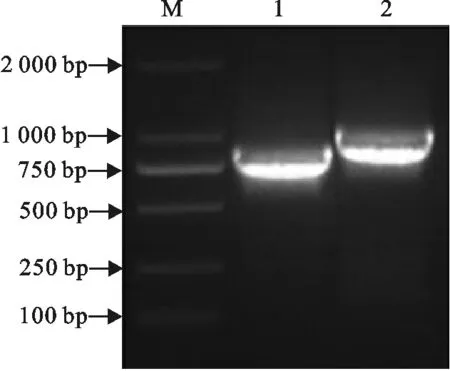
M.DNA 标准DL 2 000;1.TCR Vα5基因PCR产物;2.TCR Vβ6基因PCR产物
M.DNA Marker DL 2 000;1.PCR product of TCR Vα5 gene;2.PCR products of TCR Vβ6 gene
图1猪TCR Vα5和TCR Vβ6全长基因的PCR扩增
Fig.1PCR amplification of full-length TCR Vα5 and TCR Vβ6 genes
2.2猪αβ TCR全长基因的序列分析与结构预测
测序结果表明,克隆得到的猪TCR Vα5和Vβ6全长基因分别为816 bp和945 bp,均含有完整的开放阅读框(ORF)。采用ExPASy Proteomics Sever(http://expasy.org)中的Signal 4.0 Sever-prediction 软件分析表明,其各自含21个和24个氨基酸组成的信号肽(图2)。
2.3猪αβ TCR真核表达质粒的构建与鉴定
将构建的TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP重组表达质粒,利用XholⅠ和SalⅠ进行双酶切,可分别得到800 bp、900 bp的目的片段和5 000 bp的载体片段,表明成功地构建了猪αβ TCR真核表达质粒(图3)。
2.4荧光显微镜观察转染效率
转染24 h后,应用荧光显微镜观察293T细胞的绿色荧光产生情况,结果表明转染空载体组(D组)和转染重组质粒组(A、B、C组)均出现绿色荧光,转染效率约达70%。此外,单独转染组(A组和B组)与共同转染组(C组)绿色荧光产生情况相似,表明二者的转染效率相似;阴性对照组(E组)未见明显的绿色荧光(图4)。
2.5猪αβ TCR真核表达质粒转染细胞后mRNA水平的表达
RT-PCR 结果显示,单独转染组(A和B组)分别可以扩增到TCR Vα5和Vβ6的目的片段,共转染组(C组)可同时扩增到TCR Vα5和Vβ6基因片段,转染空载体组和阴性对照组未检测到TCR Vα5和Vβ6基因的表达,因此表明TCR Vα5和Vβ6在mRNA水平上能够单独或者共表达(图5)。

图2 猪TCR Vα5和Vβ6全长基因编码蛋白的信号肽预测
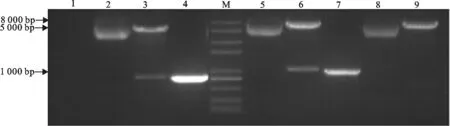
M.DNA 标准DL 8 000;1.阴性对照;2.TCR Vα5-pIRES2-EGFP质粒;3.TCR Vα5-pIRES2-EGFP双酶切产物;4.TCR Vα5 PCR产物;5.TCR Vβ6-pIRES2-EGFP质粒;6.TCR Vβ6-pIRES2-EGFP双酶切产物;7.TCR Vβ6 PCR 产物;8.pIRES2-EGFP质粒;9.pIRES2-EGFP质粒酶切
M.DNA Marker DL 8 000;1.Negative control;2.TCR Vα5-pIRES2-EGFP plasmid;3.Products of TCR Vα5-pIRES2-EGFP digested withXhol Ⅰ andSalⅠ;4.PCR product of TCR Vα5 gene;5.TCR Vβ6-pIRES2-EGFP plasmid;6.Products of TCR Vβ6-pIRES2-EGFP digested withXholⅠ andSalⅠ;7.PCR product TCR Vβ6 gene;8.pIRES2-EGFP plasmid;9.Products of pIRES2-EGFP digested withXholⅠ andSalⅠ
图3TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP真核表达质粒的双酶切及PCR鉴定
Fig.3Identification of TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP plasmids by double enzyme digestion and PCR
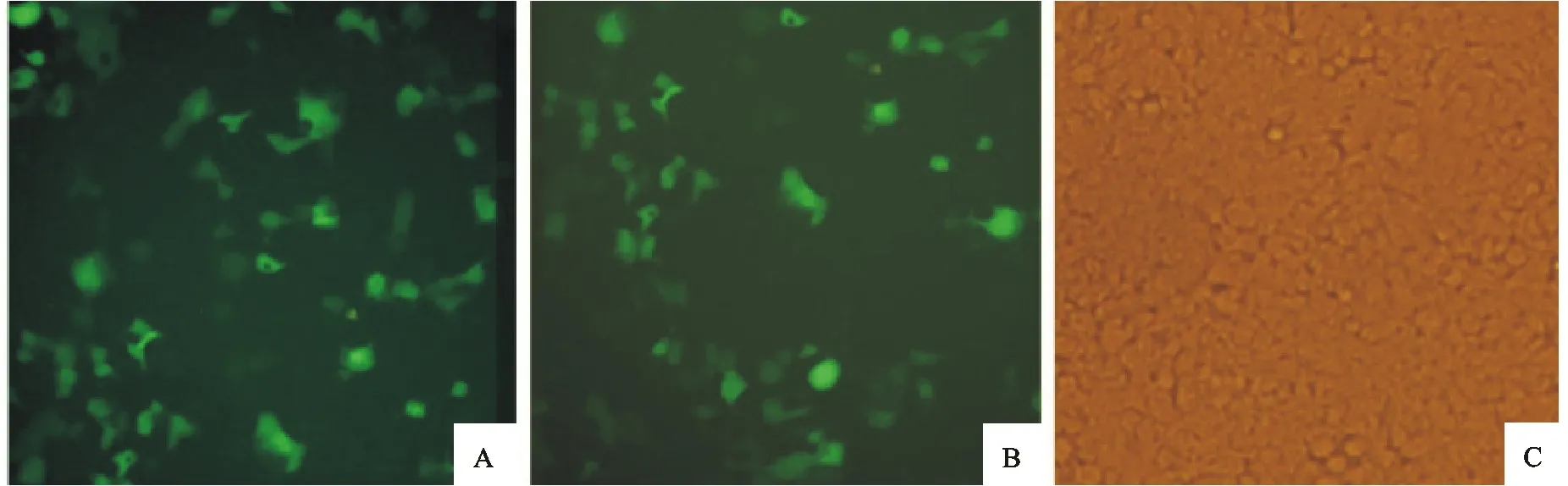
A.阳性对照;B.重组质粒;C.阴性对照
A.Positive control;B.Recombinant plasmid;C.Negative control
图4荧光显微镜下观察转染重组质粒的293 T细胞的绿色荧光产生情况(200×)
Fig.4Green fluorescence detection of 293T cells transfected with recombined plasmid(200×)

M.DNA 标准DL 2 000;1,4.转染空载体组;2.转染TCR Vα5-pIRES2-EGFP PCR 产物;3.TCR Vα5-IRES2- EGFP和TCR Vβ6-IRES2-EGFP共同转染PCR产物;5.转染TCR Vβ6-pIRES2-EGFP PCR产物;6.TCR Vα5-IRES2- EGFP和TCR Vβ6-IRES2-EGFP共同转染PCR产物
M.DNA Marker DL 2 000;1,4.Transfection with pIRES2-EGFP plasmid;2.PCR products of TCR Vα5 gene transfection with TCR Vα5-pIRES2-EGFP;3.PCR products of TCR Vα5 gene cotransfection with TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP plasmids;5.PCR products of TCR Vβ6 gene transfection with TCR Vβ6-pIRES2-EGFP;6.PCR products of TCR Vβ6 gene cotransfection with TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP plasmids
图5RT-PCR分析重组质粒转染293 T细胞后的转录表达情况
Fig.5Analysis of the transcription in 293 T cells transfected with recombinant plasmids by RT-PCR
2.6实时荧光定量PCR分析重组质粒转染293T细胞后的目的分子的转录情况
转染重组质粒48 h后,通过real-time PCR对转染重组质粒的293T细胞进行外源TCR Vα5和TCR Vβ6基因mRNA水平表达的检测。同时,我们以同比例稀释的RNA为模板进行real-time PCR检测,以排除cDNA被质粒DNA污染而影响结果判定,检测结果为阳性,提示cDNA中的确存在质粒DNA污染,但是以cDNA为模板的转染重组质粒的293T细胞外源TCR Vα5和TCR Vβ6扩增的Ct值较同比例稀释的RNA为模板扩增的Ct值提前2个~3个且远高于转染空载体组,表明转染重组质粒的细胞中外源TCR基因在mRNA水平上进行了表达(图6)。
2.7猪αβ TCR真核表达质粒转染细胞后目的蛋白的表达水平
转染48 h后,提取转染空载体、重组质粒和阴性对照组细胞的总蛋白,经SDS-PAGE电泳后,转NC膜,用小鼠抗人GAPDH单抗和兔抗鼠的IgG二抗杂交孵育后底物显色表明,所有试验组均出现与GAPDH亚基分子质量(36 ku)一致的杂交带,这说明所提取的目的蛋白量良好(图7);同时,用鼠抗人的EGFP单抗和兔抗鼠的IgG二抗杂交孵育,底物显色表明转染空载体组、转染重组质粒组均出现与EGFP蛋白分子质量(27 ku)一致的条带,说明转染空载体组和转染重组质粒组均表达了EGFP蛋白(图7)。
3 讨论
本实验室前期利用RT-PCR基因扫描分析技术,发现感染猪瘟病毒C株的外周血单个核细胞(PBMCs)中具有TCR Vα5和Vβ6亚家族基因的单/寡趋势的T细胞,该试验在此基础上扩增出其全长基因序列,其CDR3区各含保守的氨基酸基序“DDY”和“PAV”[15],这种保守的基序可能与猪瘟病毒C株的抗原肽表位的识别和结合有关。
αβ TCR是一种异源二聚体,只有共同表达、定位在T细胞膜上,才能发挥T细胞抗原受体的作用。本试验构建了TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP真核表达质粒,将其转染293T细胞观察体外表达、定位情况,判断其是否具有免疫学功能。研究结果发现,转染重组质粒和转染空载体的细胞均可看到绿色荧光,表明所构建的重组质粒转染细胞后EGFP能够有效表达,且单独转染组和共同转染组的荧光情况产生情况相似,转染效率无明显差异,初步认为TCR Vα5和Vβ6基因在转染细胞中的表达水平基本相同,但是在共同转染组中仍不能区分两种重组质粒的表达比例以及二者是否均得到了表达。因此,在后续的试验中可以采用不同的标记基因来构建真核表达载体,对两种基因共表达时的水平进行比较。其次,通过mRNA水平的比较发现,TCR Vα5和Vβ6基因均可以表达。由于目前没有商品化的猪TCR单抗,在蛋白质水平上我们只能通过Western blot检测EGFP的表达来间接反映目的基因的表达,我们发现除阴性对照组外,其他试验组均显示与EGFP分子质量大小一致的杂交条带,这表明转染质粒的细胞表达了EGFP。尹青松等[16]将弥散性大B细胞淋巴瘤(DLBCL)相关抗原特异的TCR Vα6和Vβ13分别连接入pIRES2-EGFP中,为解决αβ两种基因转入同一个真核表达载体后下游基因表达量低的问题,同时提高TCR Vβ基因的表达水平,其在mRNA和蛋白质水平上对外源基因的表达进行了检测。但是与α、β两种TCR链基因插入同一个真核表达载体相比,前者只有在α、β两种TCR链重组质粒转染入同一个T细胞后才可能形成功能性的αβ TCR异二聚体。
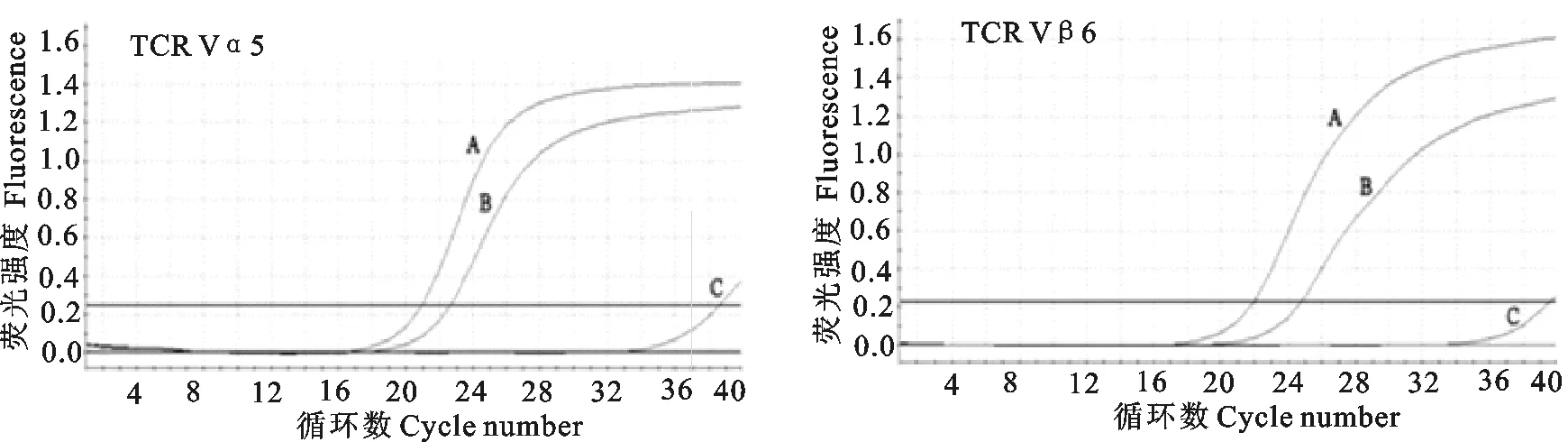
A.转染重组质粒的293T细胞的cDNA为模板;B.转染重组质粒的293T细胞的RNA为模板;C.转染空载体的293T细胞的cDNA为模板
A.cDNA template of 293T cells transfected with recombinant plasmid;B.RNA template of 293T cells transfected with recombinant plasmid;C.cDNA template of 293T cells transfected with empty vector
图6实时荧光定量PCR分析重组质粒转染293 T细胞后的转录
Fig.6Analysis of the transcription in 293 T cells transfected with recombinant plasmids by real-time PCR

1.阳性对照;2.转染TCR Vα5-pIRES2-EGFP重组质粒的293T细胞;3.转染TCR Vβ6-pIRES2-EGFP重组质粒的293T细胞;4.共转染TCR Vα5-pIRES2-EGFP和TCR Vβ6-pIRES2-EGFP重组质粒的293T细胞;5.阴性对照
1.Positive control;2.293T cells transfected with TCR Vα5-pIRES2-EGFP recombinant plasmid;3.293T cells transfected with TCR Vβ6-pIRES2-EGFP recombinant plasmid;4.293 T cells transfected with TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP recombinant plasmids;5.Negative control
图7Western blot检测转染重组质粒的293T细胞中EGFP的表达
Fig.7EGFP expression in 293T cells transfected with recombinant plasmid detected by Western blot
总之,本研究构建了猪瘟病毒C株特异的TCR Vα5和Vβ6基因的真核表达载体TCR Vα5-p IRES2-EGFP和TCR Vβ6-pIRES2-EGFP,将其单独转染或共转染不表达TCR的293T细胞中,在mRNA水平和蛋白质水平上证明了其体外表达。但是这种将TCR Vα5和Vβ6基因分别构建在同一个真核表达载体中,共同转染后是否会影响异源二聚体TCR的形成,并有效在T细胞中表达,以及转染后两种质粒能否稳定存在[16],是否对猪瘟病毒C株具有特异性的杀伤作用仍有待于进一步研究证实。
[1]Durand S V M,Hulst M M,De Wit A A C,et al.Activation and modulation of antiviral and apoptotic genes in pigs infected with classical swine fever viruses of high,moderate or low virulence[J].Arch Virol,2009,154(9):1417-1431.
[2]Graham S P,Everett H E,Haines F J,et al.Challenge of pigs with classical swine fever viruses after C-strain vaccination reveals remarkably rapid protection and insights into early immunity[J].PLoS One,2012,7(1):e29310.
[3]何小兵,贾怀杰,陈国华,等.猪瘟病毒 GSLZ 株的分离与鉴定[J].中国兽医科学,2011,41(2):131-137.
[4]Moennig V.The control of classical swine fever in wild boar[J]. Front Microbiol,2015,6:1211.
[5]Kaden V,Lange B.Oral immunisation against classical swine fever (CSF):onset and duration of immunity[J].Vet Microbiol,2001,82(4):301-310.
[6]Qiu H J,Shen R X,Tong G Z.The lapinized Chinese strain vaccine against classical swine fever virus:A retrospective review spanning half A century[J].Agr Sci China,2006,1(6):1-14.
[7]Kaden V,Glaner M.Onset and persistence of immunity to swine fever following aerogenic immunization[J].Arch Exp Vet Med,1982,36(4):593-599.
[8]Terpstra C,Woortmeyer R,Barteling S J.Development and properties of a cell culture produced vaccine for hog cholera based on the Chinese strain[J]. Deut Tierarztl Woch,1990,97(2):77-79.
[9]Graham S P,Haines F J,Johns H L,et al.Characterisation of vaccine-induced,broadly cross-reactive IFN-γ secreting T cell responses that correlate with rapid protection against classical swine fever virus[J].Vaccine,2012,30(17):2742-2748.
[10]Suradhat S,Intrakamhaeng M,Damrongwatanapokin S.The correlation of virus-specific interferon-gamma production and protection against classical swine fever virus infection[J].Vet Immunol Immunopathol,2001,83(3/4):177-189.
[11]Stubbington M J,Lnnberg T, Proserpio V,et al.T cell fate and clonality inference from single-cell transcriptomes[J].Nat Methods,2016,13(4):329-332. doi:10.1038/nmeth.3800.
[12]Höhn H,Neukirch C,Freitag K,et al.Longitudinal analysis of the T-cell receptor (TCR)-VA and-VB repertoire in CD8+T cells from individuals immunized with recombinant hepatitis B surface antigen[J].Clin Exp Immunol,2002,129(2):309-317.
[13]李扬秋.T细胞受体的研究和应用[M].北京:人民卫生出版社,2009:152.
[14]June C H,Riddell S R, Schumacher T N.Adoptive cellular therapy:A race to the finish line[J]. Sci Transl Med,2015,7(280):280ps7.doi:10.1126/scitranslmed.aaa3643.
[15]姚文娟.猪瘟病毒C株感染前后猪αβTCR变化动态与分子结构组成特征研究[D].北京:中国农业科学院,2014.
[16]尹青松,李扬秋,陈少华,等.抗原特异TCR Vα6-pIRES2-EGFP和TCRV β13-pIRES2-EGFP真核表达载体的构建及共转染研究[J].免疫学杂志,2009,25(1):1-5.
Construction and Cotransfection of Eukaryotic Expression Plasmids TCR Vα5 and Vβ6 Specific for CSFV-C Strain
WANG Chun-yan,CHEN Guo-hua,HE Xiao-bing,ZENG Shuang,JIA Huai-jie,FANG Yong-xiang,FENG Yuan,LI Shou-jie,JING Zhi-zhong
(StateKeyLaboratoryofVeterinaryEtiologicalBiology/KeyLaboratoryofVeterinaryPublicHealthofMinistryofAgriculture/LanzhouVeterinaryResearchInstitute,ChineseAcademyofAgriculturalSciences,Lanzhou,Gansu,730046,China)
Previously,we detected TCR Vα5 and TCR Vβ6 gene families specific for CSFV-C strain.In this study,to further observe the expression levelinvitro,RT-PCR technique was used to amplify the full-length sequences of TCR Vα5 and TCR Vβ6,and recombinant plasmids of TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP were also constructed and co-transfected into 293T cells to observe the expression of CSFV-C specific αβTCR gene.In result,the TCR Vα5 and TCR Vβ6 sequences were 816 bp and 945 bp,respectively;the sequences of TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIERS2-EGFP recombinant plasmids were confirmed to be correct by restriction enzyme digestion analysis and sequencing.Green fluorescence was observed with at least 70% transfected cells after 24 h post-transfection.Besides,the expression of TCR Vα5 and TCR Vβ6 were detected through real-time PCR and the EGFP protein was also verified by Western blot and the EGFP expression level was the same between two groups.These results indicated that the eukaryotic expression vector of TCR Vα5-pIRES2-EGFP and TCR Vβ6-pIRES2-EGFP specific for CSFV-C strain were constructed successfully and can be co-transfected into the 293T cells,realizing co-expression of TCR Vα5 and TCR Vβ6 geneinvitro.
Classical swine fever-C strain;TCR Vα/Vβ gene;lipofectin transfection;EGFP
2016-03-15
国家自然科学基金项目(31372423,31302072)
王春燕(1990 -),女,山东烟台人,硕士研究生,主要从事动物疫苗与分子免疫学研究。*通讯作者
S852.651;Q789
A
1007-5038(2016)10-0034-07
猜你喜欢
现代畜牧科技(2021年8期)2021-10-13
昆明医科大学学报(2021年5期)2021-07-22
江西农业学报(2021年4期)2021-04-20
幸福(2019年12期)2019-05-16
猪业科学(2018年8期)2018-09-28
猪业科学(2018年8期)2018-09-28
猪业科学(2018年4期)2018-05-19
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
