
醋酸甲羟孕酮暴露致食蚊鱼的雄激素效应
2016-10-09 05:10金子越方展强华南师范大学生命科学学院广东省高等学校生态与环境科学重点实验室广州510631
实验动物与比较医学 2016年1期
金子越, 方展强(华南师范大学生命科学学院/广东省高等学校生态与环境科学重点实验室, 广州 510631)
醋酸甲羟孕酮暴露致食蚊鱼的雄激素效应
金子越, 方展强
(华南师范大学生命科学学院/广东省高等学校生态与环境科学重点实验室, 广州 510631)
目的 探讨人工合成孕激素(醋酸甲羟孕酮,MPA)暴露对雌性食蚊鱼(Gambusia affinis)的雄激素效应。方法 将成年雌性食蚊鱼随机分为4组, 包括对照组和40 ng/L(低密度组)、200 ng/L (中密度组)和1 000 ng/L(高密度组)等3个MPA实验暴露组,并设置平行组。持续暴露28 d后观察与测定食蚊鱼的体长(BL)、体质量(BW)、身体健康指数(CF)、臀鳍第3鳍条分节数(FJ)、第4、5、6鳍条长度(FL)变化和第14、15、16椎体脉棘的形态变化等5项指标,并在7 d、14 d、21 d、28 d测定了卵黄蛋白原基因(VTGα)、雄激素受体基因(ARα)和细胞色素P450芳香化酶基因(CYP19α)mRNA表达水平的变化。 结果 食蚊鱼经MPA暴露28 d后,与对照组相比, 200 ng/L和1 000 ng/L实验组食蚊鱼的BL、BW和CF均无明显变化(分别P>0.05)。臀鳍第3鳍条分节数和第6鳍条长度变化不明显(P>0.05),但第4、5鳍条的长度显著增加(P<0.01)。暴露在200 ng/L和1 000 ng/L MPA中、高浓度组食蚊鱼的第14椎体脉棘的总长(L)、尾部尖端到脊的高度(D)值, 第15椎体脉棘的投影长(P)、P∶D值和第16椎体脉棘的P、L∶D、P∶D值分别呈现显著性变化(P<0.05或P<0.01),椎体脉棘伸长,且与脊椎骨接近垂直,显示形态雄性化效应。与对照组相比,低浓度组(40 ng/L MPA) VTGα, CYP19α基因的表达量上升(P<0.01),而ARα表达被抑制(P<0.05); 中浓度组VTGα,CYP19α, ARα基因的表达量都显著上升(P<0.01); 高浓度组CYP19α, ARα基因的表达量显著上升(P<0.01),而VTGα的表达量则被抑制(P<0.01)。结论 MPA暴露致雌性食蚊鱼骨骼出现形态雄性化,目标基因表达水平的变化结果证明MPA对食蚊鱼具雄激素效应,表明MPA是一种具有雄激素效应的孕激素。
醋酸甲羟孕酮(MPA); 食蚊鱼; 雄性化; 细胞色素P450芳香化酶基因(CYP19α); 卵黄蛋白原基因(VTGα)、雄激素受体基因(ARα); mRNA表达; 骨骼形态学
孕激素(progestin)是由卵巢的黄体细胞分泌,以孕酮(黄体酮)为主,它通常在雌激素作用的基础上产生效用。目前, 人工合成孕激素已经大量使用,如醋酸甲羟孕酮(Medroxyprogesterone 17-acetate,MPA)中文名又称“甲羟孕酮醋酸酯”、“甲孕酮”、“安宫黄体酮”等,临床上主要用于女性如痛经、功能性闭经、功能性子宫出血等疾病的治疗,大剂量可用作长效避孕针。在鱼类体内,孕激素对刺激雌鱼卵巢的最后发育和成熟、刺激雄鱼的精子形成以及对启动雌、雄鱼减数分裂都起着很重要的作用[1]。环境中的孕激素主要来源于人体、家畜和野生动物的粪便,在临床上使用的大量孕激素类产品,以及造纸工业废水中具孕激素活性的植物激素类物质等, 它们未经处理便直接地被排放到水环境中[2]。由于孕激素是雄激素、雌激素、肾上腺皮质激素等生物合成的重要中间体[3], 因此不同程度上具有上述各类激素的作用。一些研究已经表明不同的孕激素类物质表现出不同的激素效应[1],因此,可以利用实验鱼类作为指示生物了解不同水平孕激素暴露对水生生物的毒性影响。
食蚊鱼(Gambusia affinis)隶属鳉形目(Cypriondontiformes),胎鳉科(Poeciliidae),是一种原产美国东南部、墨西哥及古巴的热带性卵胎生小型鱼类,引进我国后,目前已广泛分布在华南地区不同水域。食蚊鱼具有明显的二态性,雄鱼稍细长,雌鱼腹缘圆凸,性发育成熟或者妊娠的雌鱼在臀鳍附近可以观察到孕斑。雌鱼的臀鳍和第14、15、16椎体脉棘(vertebral ribs)与雄鱼不同,性成熟的雄性个体其变长并向前弯曲,这种结构可以在雄鱼交配时为生殖足摆动提供足够的支撑[4]。另外雌鱼的臀鳍为扇形,雄鱼臀鳍部分高度分化,第3、4、5鳍条变细形成长的生殖足,其尖部具有生殖钩,雄性食蚊鱼通过生殖足给雌性食蚊鱼输送精子[5]。利用食蚊鱼的臀鳍和第14、15、16椎体脉棘的骨骼形态变化可以检测孕激素类物质暴露致鱼类的雌/雄激素效应[6]。而利用食蚊鱼目标基因mRNA表达水平变化可更敏捷地检测到水体环境激素类物质暴露致鱼类的雌/雄激素效应[7-9]。
本实验选取MPA对雌性食蚊鱼进行暴露,观察MPA对食蚊鱼骨骼发育的形态雄性化效应,同时观察食蚊鱼生殖相关的卵黄蛋白原基因(VTGα)、雄激素受体基因(ARα)和细胞色素P450芳香化酶基因(CYP19α) mRNA表达水平的变化,探讨人工合成孕激素对水生动物鱼类的雌/雄激素效应。
1 材料与方法
1.1 实验动物
食蚊鱼捕自广州市华南师范大学石牌校区池塘(该池塘已被证明未受激素类物质污染),在水族箱暂养并区分雌、雄个体。选取性成熟雌鱼进行驯养(部分雌性食蚊鱼已妊娠),持续2周使其适应生存环境,随后将实验鱼随机分配到小鱼缸。每缸保持30尾鱼,各个浓度暴露组分设2缸,每个浓度组保持60尾。实验用水在阳光照射下放置2 d,水温保持在25±2 ℃,2 d换水1次。光周期为14 h∶10 h(白天∶黑夜)。每日喂食2次,分别在上午9∶00和晚上9∶00,食用红虫购自广州花鸟鱼市场。实验鱼的使用及实验过程“参照实验动物使用的3R原则进行”[10]。
1.2 暴露药物
1.3 实验方法
1.3.1 骨骼形态学观察 设置40 ng/L(低浓度组)、200 ng/L(中浓度组)、1 000 ng/L(高浓度组) MPA 3个实验组和对照组(DMSO)。暴露实验在10 L鱼缸中进行, 每缸加入8 L处理后自来水, 采用静水更新方式的实验模式。持续暴露7 d、14 d、21 d 和28 d后,分别使用游标卡尺和电子天平测定食蚊鱼体长(BL, mm)、体质量(BW, mg)并计算健康指数[CF(%)=BW/BL×100]。同时获取不同暴露阶段食蚊鱼的骨骼组织进行染色,观察并统计数据分析骨骼的形态变化。
使用Photoshop CS3图像分析软件测量臀鳍第3鳍条分节数,臀鳍第4、5、6鳍条长度,第14/ 15/16椎体脉棘长度的L值、P值、D值即第14/ 15/16椎体脉棘总长(14L/15L/16L)、第14/15/16椎体脉棘的投影长度(14P/15P/16P)、第14/15/16椎体脉棘尾部尖端到脊柱的高度(14D/15D/16D),并根据测量的指标计算P∶D、L∶D的比值。其数据以平均值±标准差的形式表示。实验组和对照组之间的显著性差异的检验使用方差分析或者协方差分析,P<0.05为差异有统计学意义。
1.3.2 目标基因mRNA表达观察 分别取各组食蚊鱼性腺、肝脏、臀鳍组织提取RNA,进行反转录,荧光定量。具体操作如下。1)引物设计 登陆NCBI网站,找到西部食蚊鱼(Gambusia affinis)的VTGα cDNA、ARα cDNA的序列CYP19α中间片段序列[11],利用Primer3软件,由华大基因生物工程公司合成所需要的引物。相关基因扩增引物名称及其序列见表1。2)RNA制备 将性腺和肝脏组织匀浆,臀鳍条则研磨成粉末,分别加入1 mL 的Isoplus RNA(RNAiso Plus)提取液(TakaRa)进行总RNA提取。RNA提取后进行基因组DNA的去除,并检测RNA浓度及其完整性, 保证A260/A280的比值在1.8~2.0 (TakaRa)。3)RNA反转录成cDNA使用TakaRa Code: DRR037 PrimScriptTMRT reagents Kit (Perfect Real Tome) 产品。RT反应液配置如下:5×PrimeScript Buffer (2 μL), PrimeScript PT Enzyme Mix I (0.5 μL), Random 6 mers (100 μmol/L,0.5 μL), Oligo dT Primer (50 μmol/L, 0.5 μL), Total RNA (1 μL), Rnase Free dH2O (5.5 μL), 总共10 μL。反转录反应条件如下:37 ℃ 15 min (cDNA合成),85 ℃ 5s(酶失活)。4)实时荧光定量 使用TakaRa Code:DRR081A SYBR®Premix Ex TaqTMII (2×)10.0 μL, 正反向引物(10 μmol/L)各0.8 μL,ROX References Dye I (50×) or ROX References Dye II (50 ×) 0.4 μL, cDNA模板2.0 μL, H2O(灭菌) 6.0 μL,总共20.0 μL。按照两步法PCR扩增标准程序进行RT-PCR反应。

表 1 目标基因扩增引物名称及其序列Table 1 Name and sequence of the target gene amplified with primers
RT-PCR扩增结果数据的处理采用相对定量法,内参基因选用β-actin。通过标准曲线对对照样品、待测样品的目的基因及管家基因进行定量,然后根据计算公式求得相对值即为相对表达值。校正值=目的基因定量结果/管家基因定量结果;相对值=待测样品的校正值/对照样品的校正值。使用SPSS 19.0统计软件对所得数据进行统计学分析,采用单因素方差分析(One way-ANOVA)法对数据进行差异性分析。用Excel 2013做柱形图。设置P<0.05为差异有统计学意义。
2 结果
2.1 MPA暴露后食蚊鱼的BL、BW和CF变化
食蚊鱼暴露在MPA初期,其食欲显著下降,至7~14 d恢复到暴露前的喂食量,持续至28 d后活动行为恢复正常。暴露初期,200 ng/L组实验鱼死亡2尾、1 000 ng/L组死亡1尾,经解剖发现死亡个体均已经妊娠, 由此表明MPA暴露对妊娠食蚊鱼造成一定影响。持续暴露28 d后各实验组中均没有观察到体表出现明显变化的个体, 不同浓度组食蚊鱼的BL、BW和CF变化都不明显(P>0.05)。虽然在暴露初期食蚊鱼的食欲下降,在此期间CF也发生一些改变,但经2周后食蚊鱼的食欲恢复正常,本实验只检测28 d后的健康指数,而没有观察不同暴露时期食蚊鱼的CF。
2.2 MPA暴露后食蚊鱼骨骼的形态变化
经过28 d的MPA暴露后, 食蚊鱼臀鳍的第3鳍条的分节数(FJ)并没有出现明显变化,但是臀鳍第4和第5鳍条长度变化明显, 其中中浓度组(200 ng/L)第5臀鳍条长度发生显著性增长(P<0.01); 高浓度组(1 000 ng/L)第4、5臀鳍条长度也发生显著性增长(P<0.01)。而第6臀鳍条长度均没有发生明显变化(P>0.05)。
《卫风·伯兮》中提到,“焉得谖草,言树之背。愿言思伯,便我心痗。”说的就是树荫之下生长的忘忧草,能够消除我对你的相思之苦,但是我甘愿相思成病,只希望亲爱的爱人能快些回来。
暴露MPA 28 d后,中、高浓度组食蚊鱼第14椎体脉棘L值显著增长(分别P<0.01), 其他则均无显著性变化(图1A)。中浓度组第15、16椎体脉棘P值显著下降(分别P<0.01),高浓度组第16椎体脉棘P值显著下降(P<0.05),其他均无明显变化(图1B)。中、高浓度组第14椎体脉棘D值发生显著变化(分别P<0.05和P<0.01), 其他均没有发生显著新变化(图1C)。中、高浓度组第16椎体脉棘L∶D值显著性下降(P<0.01),其他均没有发生显著性变化(图1D)。中、高浓度组第15(P<0.05, P<0.01)、16椎体脉棘(分别P<0.01) P∶D值极显著性下降, 其他均没有发生显著性变化(图1E)。以上结果表明雌性食蚊鱼臀鳍和椎体脉棘出现明显的形态雄性化现象。
2.3 MPA暴露食蚊鱼目标基因的表达水平
2.3.1 ARα基因 如表2所示,低浓度组(40 ng/L)MPA暴露对食蚊鱼臀鳍组织ARα基因的表达持续21 d后影响不明显,至28 d时则显著抑制其表达(P<0.05); 中浓度组(200 ng/L)对ARα基因的表达起到促进作用,与对照组相比,暴露7~21 d均显著提高ARα mRNA的表达量(P<0.05),至28 d其影响才不明显(P>0.05); 高浓度组(1 000 ng/L)与中浓度组相类似, 暴露7 d和14 d后, MPA对ARα的影响为促进作用(P<0.05),随后持续至21 d和28 d后,ARα mRNA的表达量有所回落,与对照组相比,差异不显著(P>0.05)。结果表明,低MPA对ARα表达为抑制作用,中、高浓度MPA对ARα表达促进作用,但是中浓度组(200 ng/L)对ARα mRNA的表达量上调更高。
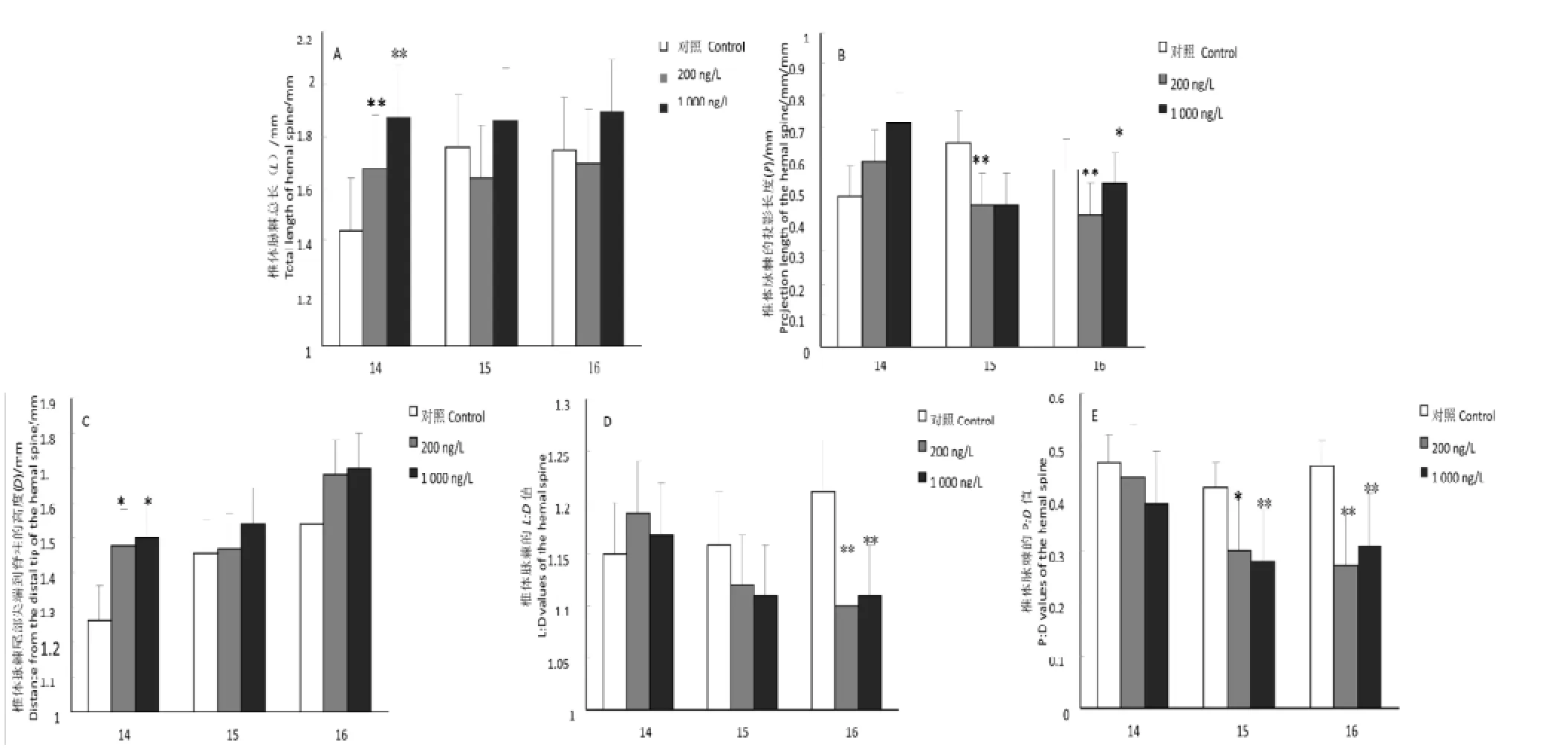
A, B, C, D, E中横标14、15、16分别代表第14、15、16椎体脉棘;与对照组比较,*P<0.05;**P<0.01图 1 MPA暴露28 d后食蚊鱼椎体脉棘L值、P值、D值、L: D值和P: D值的影响A, B, C, D, E: 14, 15 and 16 represented the 14th , 15th and 16th hemal spine. The symbol*indicated significant difference compared with control group, symbol**indicated very significant differenceFigure 1 Effects of progesterone on the L, P, D, L: D and P: D values in the female G. affinis after exposure for 28 days
2.3.2 VTGα基因 如表2所示,不同浓度MPA对食蚊鱼VTGα基因表达起到不同的作用,低浓度对VTGα mRNA的表达起到了促进作用, 与对照组比较其VTGα mRNA表达量最高达到15.9倍(P<0.01);中浓度组VTGα mRNA表达量不断提升并在14 d开始趋于平缓;而在高浓度组中,VTGα mRNA的表达量却被显著抑制,mRNA的表达量仅为对照组的0.1倍左右(P<0.01)。结果显示,随时间的持续mRNA表达量也随着提升的关系。
2.3.3 CYP19α基因 如表2所示,MPA对食蚊鱼CYP19α基因起正调控作用,低、中浓度组分别随着时间的增加表达量逐渐提升,在中浓度组中CYP19α mRNA的表达量最高(P<0.01); 而在高浓度组中,CYP19α mRNA的表达量在7 d的时候达到最高值,随后CYP19α mRNA的表达量逐渐降低,并在28 d,CYP19α mRNA的表达量和对照组无明显差异(P>0.05),整体呈趋于平缓的趋势。
3 讨论
3.1 MPA暴露对食蚊鱼生长发育的影响
CF的变化能够反映摄食、脂质沉积和蛋白质平衡[12]。用孕酮处理食蚊鱼时显示, 低浓度组和对照组之间BL和CF都没有表现出明显差异, 而高浓度组的BW和对照组相比时则显著下降(P<0.05)[13]。雌性食蚊鱼经过不同浓度的MPA暴露28 d后, BW与对照组相比没有明显的变化; 其BW与CF虽略低于对照组, 但差异不明显, 这表明中、高浓度MPA并未使食蚊鱼健康状况受到影响。但在实验中观察到妊娠食蚊鱼死亡可能与肝脏损伤有关, 推测MPA暴露影响了食蚊鱼肝脏脂类代谢, 由于雌性食蚊鱼妊娠,幼体对母体营养需求对食蚊鱼产生负荷, 从而影响肝脏激素代谢和解毒, 导致妊娠雌性食蚊鱼的死亡。
3.2 MPA暴露对雌性食蚊鱼骨骼形态发育的影响
食蚊鱼臀鳍共有10根鳍条,每根鳍条又由许多的分节来构成,已有研究表明[14],暴露在含有雄激素类物质的水环境中致食蚊鱼第3臀鳍条分节数显著增加,第4、5、6臀鳍条长度也显著增长;高浓度孕酮暴露致食蚊鱼臀鳍第3鳍条分节数显著增加[13],这表明食蚊鱼骨骼出现明显的形态雄性化特征。本实验结果显示,雌性食蚊鱼第4、5臀鳍条的长度显著增加,这也表明MPA暴露后致食蚊鱼臀鳍发生形态雄性化的变化,MPA暴露对食蚊鱼具有雄激素效应。将雌、雄性食蚊鱼暴露在雄激素或者雌激素中都使其第14、15、16椎体脉棘发生形态变化[14]。雄性食蚊鱼的椎体脉棘发育具有先后性,研究表明第16椎体的脉棘首先发育,紧随其后的是第14和15椎体[15]。但范俊杰等[13]的暴露实验结果显示,第14椎体发育对雄激素活性物质比对第15、16的更敏感。在本实验中,雌性食蚊鱼暴露于MPA 28 d之后,仅第14椎体发生显著性增长,但第15、16椎体骨骼增长则不明显;MPA暴露对第14椎体脉棘的D值有显著性增长,其结果与范俊杰等的结果相一致。本实验中浓度MPA暴露致第15、16椎体脉棘P值显著降低,高浓度组第16椎体脉棘P值也显著性降低。P、D 和L的两两比值是研究内分泌干扰物(EDCs)效应的一项非常有价值的测量指标[15]。在本实验中,第16椎体脉棘的L∶D值显著性降低,第15、16椎体脉棘的P∶D值显著性降低。可见,经过28 d MPA暴露以后,食蚊鱼的第14、15、16椎体脉棘的发育都发生不同程度的变化,L变长以及P值的减小显示骨骼变长并向中线移动,向前移动的椎体脉棘的长度是受雄激素类物质控制,这与雄激素、孕酮暴露食蚊鱼产生的效应相似,因而表明MPA暴露出现雄激素效应。
虽然MPA暴露致使食蚊鱼出现不同程度的形态雄性化效应,但是与强雄激素类比较,孕酮对食蚊鱼的生长发育影响并不十分显著,推测可能是暴露时间不长以及浓度设置较低对性成熟的雌鱼暴露产生的效应结果。MPA为人工合成的孕激素,在进入体内以后可能会参与其他反应或被进一步修饰,从而显示为弱雄激素效应。MPA暴露对食蚊鱼骨骼发育产生影响,使雌性食蚊鱼形态雄性化。
3.3 MPA暴露对食蚊鱼目标基因表达的影响
MPA暴露诱导VTGα基因表达, 这可能与雌激素有关,孕激素在体内往往和雌激素一起发挥作用以改变体内激素水平。本实验表明,随着MPA暴露浓度的提高, VTGα mRNA的表达量没有上升反而趋于平缓, 在高浓度组(1 000 ng/L)中VTGα mRNA的表达量则受到显著抑制,这与Huang 等[16]的实验结果相一致,他们将食蚊鱼暴露在1 000 ng/L孕激素中,肝脏中的VTGα、VTGβ、VTGγ mRNA表达量都被显著抑制,表明VTG mRNA的表达量与暴露剂量相关,推断这可能与抗雌激素效应有关。AR为雄激素受体,其与雄激素结合才能发挥其作用。本实验中,低浓度组MPA暴露,ARα mRNA的表达受到抑制,而在中、高浓度组ARα mRNA的表达量则被上调, 以200 ng/L组表达量最高,mRNA的表达量不呈剂量依赖关系。推测ARα表达被抑制可能和雌激素的抗雄激素效应有关,而暴露浓度的提升对ARα为正调控,可能是体内生成雄激素促进了基因表达,因为孕激素可以是雄激素的前体,当孕激素反应生成雄激素的时候,就可以促进ARα基因的表达。
本实验食蚊鱼暴露在不同浓度的MPA中,低、中浓度组CYP19α mRNA的表达量随着时间的推移表达量逐渐上升;而高浓度组(1 000 ng/L)在7 d达到最高值随后表达量逐渐降低,在28 d时CYP19α mRNA的表达量和对照组相近。高浓度组CYP19α mRNA表达量的降低可能是CYP19α蛋白催化睾酮(T)(或者T衍生物)生成雌二醇(E2)(或者E2衍生物), 由于E2浓度过高而产生抑制作用。而低、中浓度组CYP19α mRNA的表达量并没有被抑制, 这可能与暴露试剂浓度有关, 也有可能与其他机制相关。
MPA暴露对食蚊鱼体内目标基因的影响是内在联系的,通过对实验结果的比较观察到,不同浓度MPA暴露对食蚊鱼目标基因的表达产生不同的影响。低浓度(40 ng/L)MPA和雌激素共同作用促使VTGα基因表达显著上升,MPA可能在体内通过一系列的反应生成雄烯二酮(AED)和T及其衍生物,正向调节了CYP19α mRNA的表达,生成雌醇及E2同时促进VTGα基因表达,而雌激素水平的提高则对ARα产生抗雄激素作用因而抑制了其表达。中浓度(200 ng/L) MPA持续刺激CYP19α mRNA的表达,从而产生更多E2,使得体内的E2水平大大提高, 在反应效率相同的情况下,暴露浓度的提升使体内的T水平也提高,T通过血液循环进入臀鳍, 促进了臀鳍ARα基因的表达; 同时E2水平的提高则对VTGα基因的表达产生负反馈, 降低了VTGα mRNA的表达量, 但与对照组相比较依然是显著性提高。而在高浓度(1 000 ng/L)组中,MPA进一步促进CYP19α的表达,产生大量E2, 不仅显著性地抑制VTGα mRNA的表达, 也引起本身的负反馈机制。闫月明等[17]的研究表明, 高浓度17α-甲基睾酮会抑制CYP19α表达, 所以体内MPA水平的提高使得T、AED水平提高的同时也会抑制CYP19α表达。以上结果表明随着暴露时间的增加CYP19α mRNA水平降低,而ARα的表达量也有所回落,可能是由于T或者其衍生物在体内参加其他途径反应造成ARα的表达量下降。这些结果表明,高浓度MPA暴露将造成所有目标基因都处于负调控当中,继续提升MPA浓度将对机体产生的毒性效应如何将有待作进一步研究。
[1] Paulos P, Runnalls TJ, Nallani G, et al. Reproductive responses in fathead minnow and Japanese medaka following exposure to a synthetic progestin, norethindrone [J]. Aquat Toxicol,2010, 99(2):256-262.
[2] Arcend-Hoy LD, Nimrod AC, Benson WH. Endocrinemodulating substances in the environment: estrogenic effects of pharmaceutical products [J]. Toxicol, 1998, 17:139.
[3] Conneely OM, Mulac-Jericevic B, DeMayo F, et al. Reproductive functions of progesterone receptors [J]. Recent Prog Hormone Res, 2002, 57(1):339-355.
[4] Rosen DE, Gordon M. Functional anatomy and evolution of male genitalia in Poeciliid fishes [J]. Zoologica, 1953, 38(1): 1-52.
[5] Rosa-Molinar E, Hendricks SE, Rodriguez-Sierra JF, et al. Development of the anal fin appendicular support in the western mosquitofish, Gambusia affinis affinis (Baird and Girard 1854): A reinvestigation and reinterpretation [J]. Acta Anat (Basel), 1994, 151(1):20-35.
[6] 范俊杰, 黎紫兰, 徐少群, 等. 孕激素暴露致食蚊鱼形态雄性化的生物学效应[J]. 中国实验动物学报, 2014, 22(4): 31-36.
[7] 谢勇平, 方展强. 利用食蚊鱼目标基因转录水平评价东莞寒溪河雌/雄激素物质污染现状[J]. 水生生物学报, 2013,37(4):691-697.
[8] 谢勇平, 方展强. 城市废水暴露致食蚊鱼形态雌/雄性化的生物学效应[J]. 生物灾害科学, 2015, 38(2):106-113.
[9] 侯丽萍, 舒琥, 李强, 等. 利用食蚊鱼目标基因转录水平评价四会市邓村河雌/雄激素物质污染现状[J]. 安徽农业大学学报, 2015, 42(1):152-158.
[10] 贺争鸣, 李冠民, 邢瑞昌. 3R理论的形成、发展及在生命科学研究中的应用. 实验动物科学与管理, 2000, 17(3): 43-47.
[11] 甘为. 食蚊鱼CYP19α基因的克隆及三丁基锡暴露对其表达的影响[D]. 广州: 华南师范大学, 2015.
[12] Goede RW, Barton BA. Organisnic indices and an autopsybased assessment as indicators of health and condition in fish [C]. In: Adam SM (ed) Biological indicators of stress in fish. American Fisheries Society, Bethesda, 1990, pp93-108.
[13] 范俊杰, 徐少群, 方展强, 等. 17α-甲基睾酮对食蚊鱼形态雄性化及目标基因表达的影响[J]. 水产学报, 2013, 37(1): 9-15.
[14] Xie Y, Fang Z, Hou L, et al. Altered development andreproduction in western mosquitofish (Gambusia affinis)found in the Hanxi river, Southern China [J]. Environ Toxicol Chem, 2010, 29(11):2607-2615.
[15] Rawson CA, Lim RP, Warne MSJ. Skeletal morphology and maturation of male Gambusia holbrooki exposed to sewage treatment plant effluent [J]. Ecotoxicol Environ Saf, 2008, 70 (3):453-461.
[16] Huang J, Shan J, Kim D, et al. Basolateral chloride loading by the anion exchanger type 2: role in fluid secretion by the human airway epithelial cell line Calu-3[J]. J Physiol, 2012, 590 (21):5299-5316.
[17] 闫月明. 多氯联苯(Aroclor 1248)暴露对食蚊鱼CYP19α mRNA表达的影响及毒性效应研究[D]. 广州: 华南师范大学, 2013.
Androgenic Effects of Synthetic Progesterone (Medroxyprogesterone acetate) on Gambusia affinis
JIN Zi-yue, FANG Zhan-qiang
(Key Laboratory of Ecology and Environmental Sciences of Guangdong Higher Education,College of Life Science, South China Normal University, Guangzhou 510631, China)
Objective To investigate the androgen effects of synthetic progesterone (Medroxyprogesterone acetate, MPA) exposure on the female mosquitofish (Gambusia affinis). Methods The mature female mosquito fish were randomly divided into four groups, with one control group and three experimental groups exposed with 40 ng/L, 200 ng/L and 1 000 ng/L MPA respectively. Parallel experimental groups were also established. After 28-day-long exposure, five indexes including the body length (BL),body weight (BW), health index (CF), the section number (FJ) of the 3rd anal fin, the length (FL) change of the 4rd, 5rd, 6rd anal fin, and the morphological changes in the 14, 15 and 16 vertebral ribs in mosquito fish were observed and measured. The mRNA expression levels of vitellogenin gene (VTGα), cytochrome P450 gene (CYP19α) in livers, and androgen receptor gene (ARα) in anal were also determined after 7, 14, 21 and 28 days exposure. Results The BL, BW and CF of experimental groups exposed in concentrations of 200 ng/L and 1000 ng/L MPA for 28 d were not changed significantly (respectively,P>0.05) when compared with those of the control group. The 3rd anal fin of FJ and the 6rd anal fin of FL were not changed significantly (P>0.05), however, the 4rd, 5rd of FL in experimental groups had been increased in varying degrees (P<0.01). When exposed in 200 ng/L and 1000 ng/L MPA, the L, D values of the 14th vertebral ribs, the P, P∶D value of the 15th vertebral ribs, and the P, L∶D, P∶D value of the 16th vertebral ribs in female G. affinis were appeared very significantly different (P<0.05 or P<0.01), respectively. Centrum haemal spine elongation and with the spine nearly vertical were displayed significant changes in morphological masculinization. Compared with the control group, low concentration groups (40 ng/L MPA) VTGα, CYP19α genes expression were increased significantly (P<0.01), while ARα expression was inhibited (P<0.05); medium concentration groups (200 ng/L MPA) VTGα, CYP19α,ARα genes expression were increased significantly (P<0.01); high concentration groups (1 000 ng/L MPA) CYP19α, ARα genes expression were increased significantly (P<0.01), however the expression of VTGα was inhibition (P<0.05). Conclusion MPA exposure induced skeletal development morphological masculinizing effects in female mosquitofish, and the results of target gene expression level changes showed that the effects of androgen induced by MPA in mosquitofish, indicating that the MPA was a kind of androgen effect of progesterone.
Medroxyprogesterone(MPA); Gambusia affinis; Masculinization; Cytochrome P450 gene (CYP19α); Vitellogenin gene (VTGα); Androgen receptor gene (ARα); mRNA expression;Skeleton morphology
Q95-33
A
1674-5817(2016)01-0006-07
10.3969/j.issn.1674-5817.2016.01.002
2015-08-24
广东高校城市水环境生态治理与修复工程技术研究中心建设项目(2012gezxA004)
[作者介绍] 金子越(1989-), 男, 硕士, 研究方向: 水生生物学。E-mail: komsjk1990@163.com
方展强(1953-), 男, 教授, 博士生导师, 研究方向: 水生生物学。E-mail: fangzhq@scnu.edu.cn
猜你喜欢
现代仪器与医疗(2021年6期)2022-01-18
有色金属(矿山部分)(2021年4期)2021-08-30
世界科学技术-中医药现代化(2021年12期)2021-04-19
中国生殖健康(2019年4期)2019-12-14
农药科学与管理(2019年10期)2019-04-20
中成药(2017年8期)2017-11-22
现代冶金(2016年6期)2016-02-28
上海大学学报(自然科学版)(2016年6期)2016-02-28
中国继续医学教育(2015年4期)2016-01-07
川北医学院学报(2015年5期)2015-12-05
