
Expression of HIF-1α and Its Target Genes in the Nanorana parkeri Heart: Implications for High Altitude Adaptation
2016-09-28 06:55QiongZHANGXingzhiHANYinziYERobertKRAUSLiqingFANLeYANGandYiTAO
Asian Herpetological Research 2016年1期
Qiong ZHANG, Xingzhi HAN, Yinzi YE, Robert H. S. KRAUS, Liqing FAN, Le YANGand Yi TAO
1Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences,Beijing 100101, China
2College of Life Sciences, Harbin Normal University, Harbin 150025, Heilongjiang, China
3College of Life Sciences, Hangzhou Normal University, Hangzhou 311121, Zhejiang, China
4Department of Biology, University of Konstanz, Konstanz 78457, Germany
5Max Planck Institute for Ornithology, Department of Migration and Immuno-Ecology, Am Obstberg 1, Radolfzell 78315, Germany
6College of Agricultural and Animal Husbandry, Tibet University, Tibet 860000, China
7Tibet Plateau Institute of Biology, Tibet 850001, China
Expression of HIF-1α and Its Target Genes in the Nanorana parkeri Heart: Implications for High Altitude Adaptation
Qiong ZHANG1,*, Xingzhi HAN2, Yinzi YE3, Robert H. S. KRAUS4,5, Liqing FAN6, Le YANG7and Yi TAO1
1Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences,Beijing 100101, China
2College of Life Sciences, Harbin Normal University, Harbin 150025, Heilongjiang, China
3College of Life Sciences, Hangzhou Normal University, Hangzhou 311121, Zhejiang, China
4Department of Biology, University of Konstanz, Konstanz 78457, Germany
5Max Planck Institute for Ornithology, Department of Migration and Immuno-Ecology, Am Obstberg 1, Radolfzell 78315, Germany
6College of Agricultural and Animal Husbandry, Tibet University, Tibet 860000, China
7Tibet Plateau Institute of Biology, Tibet 850001, China
Hypoxia-inducible factor 1 alpha (HIF-1α) and its target genes vascular endothelial growth factor (VEGF)and transferrins (TF) play an important role in native endothermic animals' adaptation to the high altitude environments. For ectothermic animals - especially frogs - it remains undetermined whether HIF-1α and its target genes (VEGF and TF) play an important role in high altitude adaptation, too. In this study, we compared the gene sequences and expression of HIF-1α and its target genes (VEGF and TF) between three Nanorana parkeri populations from different altitudes (3008 m a.s.l., 3440 m a.s.l. and 4312 m a.s.l.). We observed that the cDNA sequences of HIF-1A exhibited high sequence similarity (99.38%) among the three altitudinally separated populations; but with increasing altitude, the expression of HIF-1A and its target genes (VEGF and TF) increased signifi cantly. These results indicate that HIF-1α plays an important role in N. parkeri adaptation to the high altitude, similar to its role in endothermic animals.
Hypoxia, cold-temperature, ectothermic animals, Nanorana parkeri, high altitude, vascular endothelial growth factor, transferrins, anura, amphibia
1. Introduction
A high mountain range's plateau environment is hostile to life due to the low atmospheric oxygen pressure (up to about 40% lower than at sea level on the Tibetan plateau,for example), cold climate and strong ultraviolet radiation. Hypoxic conditions may compromise cell and organ metabolism; especially for the heart, because the heart is an obligate aerobic organ. Under hypoxic conditions,the heart muscle not only cannot produce enough energyto maintain essential cellular processes, but also may be subjected to cardiac dysfunction, ultimately leading to death (Giordano, 2005). Organisms with long-term adaptations to high altitude environments have evolved a set of specific physiological traits to survive in this harsh environment. The study of the evolutionary basis of adaptive mechanisms to alleviate hypoxia not only has important biological, but also clinical implications. This offers the opportunity to contribute to fundamental human medical research by means of evolutionary studies (Rose,2001).
Endothermic animals native to high altitude areas,such as the domestic yak (Bos grunniens), plateau pika(Ochotona curzoniae) and the human Tibetan population have developed traits to survive in highly hypoxic environments. Examples for such adaptations are largerlung capacity, lower pulmonary arterial pressure,and higher haemoglobin concentration (Cruz et al.,1980; Moore et al., 2000; Li et al., 2001; Wu and Kayser, 2006). Compared with endothermic animals,ectothermic animals-especially frogs-carry many special characteristics, such as the incomplete development of the respiratory and circulatory system, abundant skin secretion, and also pronounced hypoxia tolerance(Knickerbocker and Lutz, 2001; Stewart et al., 2004). Previous studies showed that they have evolved a highly efficient and well-regulated metabolism to counter the impacts of extreme environmental conditions in the fi eld. For example, Telmatobius coleus, one of the plateau anurans, harbours an increased skin surface area where the cutaneous capillaries penetrate to the outer layer of the skin, and has elevated haemoglobin concentration and haematocrit in comparison with sea-level anurans(Hutchison et al., 1976).
The molecular mechanisms underlying these phenotypic traits are modulated by several specific genes. For example, vascular endothelial growth factor(VEGF) plays an important role in adaptation to high altitude hypoxia environments for plateau pika (Li et al., 2013) and the Peruvian human population in the Andes (Espinoza et al., 2014). Transferrins (encoded by TF) play an important role in iron transportation during erythropoiesis in Ethiopians (Beall et al., 2002). Expression levels of egl nine homolog 1 (EGLN1)and peroxisome proliferator-activated receptor alpha(PPARA) were signifi cantly associated with the decreased haemoglobin phenotype in Tibetan human populations(Simonson et al., 2010). The ADAM metallopeptidase domain 17 (ADAM17), arginase 2 (ARG2) and matrix metalloproteinase-3(MMP3) genes were detected to be under positive selection in Yak (Qiu et al., 2012), and chemokine (C-C motif) ligand 2 (CCL2) and pyruvate kinase isozymes R/L (PKLR) in Tibetan antelope(Pantholops hodgsonii; Ge et al., 2013).
All genes mentioned above are parts of the hypoxiainducible factor (HIF) pathway. HIFs are crucially involved in maintaining oxygen homeostasis. They are composed of a labile hypoxia-regulated α subunit, so called HIF-1α, -2α or -3α, and a constitutive β subunit(Wenger and Gassmann, 1997). HIF-1α plays a critical role in transcriptional regulation of the amount and timing of targeted gene production during hypoxia,which mediates many genes involved in erythropoiesis,angiogenesis, autophagy, and energy metabolism (William and Peter, 2008). For example, HIF-1α regulates VEGF(Forsythe et al., 1996), which is a major mediator of vasculogenesis and angiogenesis and protects endothelial cells from undergoing apoptosis (Nor et al., 1999). TF encodes transferrins, which are other proteins modulated by HIF-1α. They mediate cellular iron uptake and deliver iron to cells requiring it (Tacchini et al., 1999). Iron is essential for oxygen delivery, as it is incorporated in the newly synthesized haemoglobin throughout erythropoiesis. Therefore, HIF-1α is a key transcription factor that regulates a variety of cellular and systemic adaptations to hypoxia; VEGF and TF are pivotal target genes of HIF-1α in angiogenesis and erythropoiesis under hypoxia. Although physiological responses to hypoxia have been extensively studied in plateau frogs (e.g. Weber et al., 2002), whether HIF-1α plays an important role in ranid adaptation to high elevation environments, like it does in endothermic animals, is poorly understood.
The Qinghai-Tibetan plateau (at greater than 4000 m a.s.l.) is the highest plateau in the world, which provides the best opportunity for us to study the adaptation of ectothermic animals to high altitude hypoxic environments in their natural habitat. Nanorana parkeri is an anuran endemic to the southern Tibetan plateau and distributes across a narrow latitudinal (28 to 31°N)but extensive altitudinal range (2850 to 5100 m a.s.l.). Therefore, N. parkeri represents the highest altitude ranid in the world (Hu, 1987). Across the species' altitudinal range, environmental conditions vary large, for example,annual mean temperature ranged from 3.0°C to 8.6°C;air oxygen content ranged from 88 to 114 mg/cm3(Zhang et al., 2012). Although N. parkeri has been a model to study morphology, life history and biological chemistry in high altitude environments (Ma et al., 2009; Ma and Lu, 2009, 2010; Lu et al., 2010; Zhang et al., 2012), the role of HIF-1α in their adaptation to high altitude remains undetermined. In this study, we compared the expression of HIF-1α and its target genes in N. parkeri in heart tissue(VEGF and TF) between populations of three different altitudes (low: 3008 m a.s.l., medium: 3440 m a.s.l., high:4312 m a.s.l.) .
2. Materials and Methods
2.1 Sample preparation Healthy adult Nanorana parkeri were captured at various altitudes (3008 m a.s.l.,3440 m a.s.l. and 4312 m a.s.l.) in the Sejila Mountains,in Nyingtri county, Tibet in June 2014 (Table 1). Five individuals for each altitude were used for HIF-1α quantification. Animals were killed by doublepithing technique adopted from Costanzo et al. (1991)immediately upon capture to harvest heart tissue. Half ofthe tissue preserved in RNA holder (TransGen Biotech Co., Ltd., Beijing, China, stored at room temperature),was brought to our laboratory in Beijing and used for RNA extraction. The remaining tissue was frozen at -80°C and transferred to our laboratory for protein extraction. All procedures involved in the handling and care of animals were in accordance with the China Practice for the Care and Use of Laboratory Animals and were approved by China Zoological Society.
2.2 RNA extraction and primer preparation Total RNA was extracted and purified from N. parkeri heart using TRIZOL reagent (Invitrogen). The concentrations of RNA samples were quantifi ed with a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific Inc., DE)for further analyses.
We designed HIF-1A, VEGF and TF primers according to the whole-genome sequence of N. parkeri (Sun et al., 2014) and homologous sequences of Human (Homo sapiens), yak (Bos grunniens), common frog (Rana temporaria), rainbow trout (Oncorhynchus mykiss),African clawed frog (Xenopus laevis) and tropicalis frog (Xenopus tropicalis) in GenBank (Table 2). All of the primers were produced by Shanghai Biotechnology Corporation (Shanghai, China).
2.3 RT-PCR Reverse-transcription polymerase chain reaction (RT-PCR) was performed with the Access RTPCR System (Promega) according to the manual. The total of 0.6 μg RNA isolated from N. parkeri heart for each altitude from each of five individuals were pooled into a total aliquot of three μg and reverse transcribed for 60 min at 42°C and for 10 min at 75°C with M-MLV reverse transcriptase. RT-PCRs were performed by using SYBR green PCR Master Mix (Applied Biosystems) in a 10 μl total volume, including 5 μl premix, 2 μl 1 μM each primer and 1 μl cDNA template to quantify the expression of HIF-1A, VEGF and TF mRNA. The amplification was performed for 40 cycles at the following cycle conditions: 95°C for 10 s (denaturation), 56°C for 10 s(annealing) and 72°C for 20 s (extension). Each reaction was performed in triplicate. To compare among groups,mRNA levels of target genes were measured as relative expression using 2-△△CTvalues and normalized to β-Actin generated from the same sample (Livak and Schmittgen 2001).
2.4 Sequence alignment The PCR products of HIF-1A of the three altitude groups were sequenced with an automated sequencer by the BGI Tech Solutions Corporation (Shenzhen, China). For each altitude,the PCR products from the cDNA of the pool of fiveindividuals were sequenced together. Multiple sequence alignment was carried out using DNAMAN software package (Lynnon Biosoft).
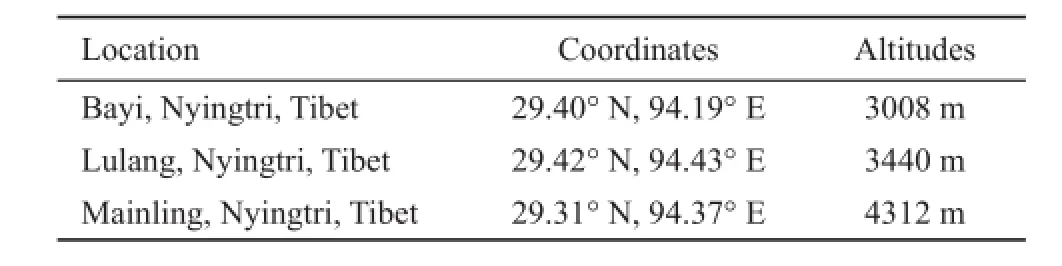
Table 1 Samples information.

Table 2 Primer details for RT-PCR.
2.5 Western blot Hearts of three samples (together 100 mg) from each altitude were homogenized in 1 ml lysis buffer (1 mM PMSF, 3 mM EDTA, 40 mM Tris (PH 7.5), 5 mM DTT). The tissue was crushed on ice, and centrifuged at 10 000 rpm (Sigma 1-15K,Germany) for 15 min at 4°C. Then the upper layer was transferred into a new 1.5 ml Eppendorf PE tube. Protein concentration was measured directly with a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific Inc.,DE). An aliquot containing 30 μg of protein was diluted in loading buffer (loading buffer:sample = 5:1, v/v,heated to 97°C for 15 min) and was separated by 10% sodium dodecyl sulfate polyacrylamide gel (SDS-PAGE)electrophoresis until the blue dye front was at the end of the gel but not diffused off the gel. Then, the protein was transferred onto a 0.45-μm-pore nitrocellulose fi lter membrane (NC, Immuno-Blot, BioRad, USA) at 9V for 1 h at 4°C. The membranes were blocked at room temperature for 1 h with 3 % fat-free milk in TBS (2M Tris, NaCl, PH 7.5). The membrane was then incubated in 1:500 diluted HIF-1α antibody (Abcam, Cambridge, UK)at 4°C overnight. After washing twice with TBS-T (1 L TBS + 200 μl Tween), and twice with TBS-every washing lasted for 10 min - the membrane was incubated with HIF-1α-ChIP grade antibody (diluted 1:10 000; AB2185,Abcam, Cambridge, MA) for 3h at room temperature. After additional washing, twice with TBS-T and twice with TBS, proteins were visualized by exposing the blot to an X-ray fi lm, and photographed with an ImageQuantLAS4000 (GE Healthcare UK Ltd, Little Chalfont, UK). The net intensities of individual bands were measured using Quantity One (version 4.6.2, Bio-Rad company,USA). Each altitude group was measured three times.
2.6 Statistical analysis of data Results were presented as mean ± S.E. per altitude group. Group means were compared by one-way analysis of variance, with a post hoc Scheffe's test. A value of P < 0.05 was considered statistically signifi cant (SPSS ver. 17.0).
3. Results
3.1 The sequence alignment of HIF-1A Chromatograms of the pooled sequences indicated no mixed signals from nucleotide variation that might have been present in the pooled individuals. We therefore infer no signs of intraaltitude genetic variation. The length of the N. parkeri HIF-1A cDNA was 2358 bp. The identity of HIF-1A cDNA sequences between altitudes was 99.38% across the three altitudes (3008 m a.s.l., 3440 m a.s.l. and 4312 m a.s.l.; Figure 1), and there were a total number of 28 variable sites. Among them, 23 were substitutions, and five of them were indels (insertions or deletions). Eight of these substitution sites lead to amino acid differences(Table 3).
3.2 The expression of HIF-1α protein The protein concentration of HIF-1α increased significantly with increasing altitude, as measured by the net intensities ± SE of individuals bands: 8.20 ± 0.8418 (low altitude),24.81 ± 1.6079 (medium altitude), and 68.63 ± 1.0281(high altitude) (Table 4 Line A, Figure 2A).
3.3 The expression of HIF-1A, VEGF and TF mRNA The expression of HIF-1A, VEGF or TF mRNA increased with altitude, too (Figure 2B, C, D). For HIF-1A and VEGF, the largest source of variance was between groups; for example, the expression of high altitude was significantly higher than the medium altitude and low altitude. For TF, the largest source of variance derived from within groups, so no signifi cant differences among the three altitudes was observed (Table 4 Lines B, C, D).
4. Discussion
Although sequence similarity of HIF-1A among samples collected from the three altitudes was high, some substitutions have led to amino acid changes (Table 3). Furthermore, there seem to be more genetic differences between the high altitude group and the two other groups. For example, six of the eight amino acid changes werebetween high altitude and the two other altitude groups;therefore, the high altitude environment seems to have resulted in the largest change in the genetic background. Variation in the amino acid sequence may induce important functional changes of HIF-1α, and could therefore be responsible for differences between altitude groups. A functional analysis of the changed amino acid residues in further proteomic experiments might shed light on the important questions of the function of this protein.

Table 3 Eight substitution sites cause amino acid differences.
Simultaneously, differential gene expression patterns among different altitude groups were observed. The HIF-1α expression of N. parkeri is increasing with increased habitat altitude. The same pattern of expression is also observed in plateau pika (Ochotona curzoniae; Li et al.,2009). These results indicate that HIF-1A is a hypoxiainducible gene in N. parkeri, just like in endothermic animals. In lower vertebrates, the role of HIF-1α in hypoxia tolerance was first reported for rainbow trout(Soitamo et al., 2001). Furthermore, the role of HIF-1α in hypoxia tolerance has also been proven indirectly by a set of target genes of HIF-1α in euryoxic fi sh (Gillichthys mirabilis; Gracey et al., 2001). Rissanen et al. (2006)found that except for hypoxia (Cao et al. 2008), cold temperature also induces the expression of HIF-1α in crucian carp (Rissanen et al., 2006). In our study,low temperature and high altitude habitats covary, and temperature could thus play an additional role in altituderelated HIF-1α regulation. The possible interaction of altitude and temperature will need to be addressed in future experiments. However, whatever up-regulated the HIF-1A expression, hypoxia or the cold temperature,HIF-1α plays an important role in the local adaptation of N. parkeri to its high-altitude environment.

Figure 1 Multiple sequence alignment of Nanorana parkeri HIF-1α cDNA at three altitudes (high altitude: HIF-1AH, medium altitude: HIF-1AM and low altitude: HIF-1AL). Asterisks indicate identical sites and the gap indicated the variable sites among three altitude sequences.

Table 4 The ANOVA results of HIF-1α protein and HIF-1A, VEGF and TF mRNA expression.

Figure 2 Expression of HIF-1α protein (A) and HIF-1A (B) , VEGF (C) and TF (D) mRNA of Nanorana parkeri at different altitudes (low,3008 m; medium, 3440 m; high, 4312 m). For mRNA, expression levels were normalized to β-actin mRNA levels. Representative results from three independent experiments in triplicate on the same protein or mRNA of different individuals are presented as means ± standard error.
Our findings indicate that VEGF mRNA levels are increased in the N. parkeri that inhabit higher altitudes. The trend is similar to the changes in HIF-1α mRNA expression with altitude. It is well known that hypoxiainduced expression of VEGF is under the control of HIF-1A in other species (Damert et al., 1997), therefore we assume that the higher expression level of VEGF mRNA may be supported by the higher expression of HIF-1α protein in N. parkeri inhabiting higher altitudes. In addition, low temperature is reported to be involved inangiogenesis through up-regulating VEGF expression by HIF in mouse adipose tissue (Xue et al., 2009). Thus,cold temperatures could also a play an important role in VEGF up-regulation, like for HIF-1A. Therefore, hypoxia and cold temperature, the two prime ecological factors of high-altitude habitat, may play an important role in the adaption of N. parkeri to high altitude environments through HIF-1A and VEGF.
Chytridiomycosis is a potentially lethal disease of amphibians caused by the amphibian chytrid fungus(Batrachochytrium dendrobatidis) that has been associated with population declines in several amphibian species throughout the world (Daszak et al., 1999; Carey, 2000;Green et al., 2002; Lips et al., 2006). Research suggests that B. dendrobatidis is more abundant in medium and high altitudes than low altitude because medium and high altitudes provide ideal temperatures for B. dendrobatidis(Daszak et al., 2003; Berger et al., 2004; Woodhams and Alford, 2005; Drew et al., 2006). The downstream gene of HIF-1A, TF, is up-regulated under hypoxic conditions in endothermic animals (e.g. Ethiopians, Beall et al.,2002; plateau pika, Ochotona curzoniae, Li et al., 2013). In our study, we indeed found a trend of increasing TF mRNA expression with increasing altitude in N. parkeri. TF is also associated with the innate immune system(Breitman et al., 1980; Evans et al., 1989; Stafford and Belosevic, 2003) as an acute phase protein in response to infection or stress conditions and limits the amount of iron, leading to the inhibition of bacterial growth (Sahoo et al., 2009). Based on the fact that orthologs of TF were identifi ed in amphibians (Moskaitis et al., 1990; Morabito and Moczydlowski, 1994; Mohd-Padil H et al., 2012)and that the amphibian's skin can excrete antimicrobial peptides (Bevins and Zasloff, 1990), we hypothesize that high expression of TF mRNA could be related to defense mechanisms against pathogenic microorganisms in high altitude.
In conclusion, comparison of HIF-1α protein and mRNA expression across various altitudes indicates the important role of HIF-1α in adaptation to a high altitude environment. Our study made the first step for the understanding of ranids' adaptation to such high altitude environments. In future, creating whole transcriptomes(Wolf, 2013) will become affordable also for ecologically oriented working groups and might allow for a fresh look without being biased towards knowledge from other systems. Candidate genes for adaptive processes have been mined with genome-wide technology before in similar experimental or empirical set-ups (Bonin et al., 2006; Kane and Rieseberg, 2007) and true RNA sequencing may help us to identify so far unknown genes and pathways.
Acknowledgements We thank the anonymous villagers in Tibet for assisting with our collecting samples. Our research was supported by National Natural Science Foundation of China (No. 31471994). The samples were analyzed at Key laboratory of Animal Ecology and Conservation Biology, Chinese Academy of Sciences.
References
Beall C. M., Decker M. J., Brittenham G. M., Kushner I.,Gebremedhin A., Strohl K. P. 2002. An Ethiopian pattern of human adaptation to high-altitude hypoxia. Proc Natl Acad Sci USA, 99: 17215-17218
Bevins C. L., Zasloff M. 1990. Peptides from frog skin. Annu Rev Biochem, 59: 395-414
Berger L., Spear R., Hines H. B. Marantelli G., Hyatt A. D.,McDonald K. R. 2004. Effect of season and temperature on mortality in amphibians due to chytridiomycosis. Aust Vet J, 82:31-36
Bonin A., Taberlet P., Miaud C., Pompanon F. 2006. Explorative genome scan to detect candidate loci for adaptation along a gradient of altitude in the common frog (Rana temporaria). Mol Biol Evol, 23: 773-783
Breitman T. R., Collins S. J., Keene B. R. 1980. Replacement of serum by insulin and transferring supports growth and differentiation of the human promyelocytic cell line HL-60. Exp Cell Res, 126, 494-498
Cao Y. B., Chen X. Q., Wang S., Wang Y. X., Du J. Z. 2008. Evolution and regulation of the downstream gene of hypoxiainducible factor-1 α in naked carp (Gymnocypris przewalskii)from lake Qinghai, China. J Mol Evol, 67: 570-580
Carey C. 2000. Infectious disease and worldwide declines of amphibian populations, with comments on emerging diseases in coral reef organisms and in humans. Environ Health Persp, 108:143-150
Costanzo J. P., Lee R. E., Wright M. F. 1991. Effect of cooling rate on the survival of frozen wood frogs, Rana sylvatica. J Comp Physiol B, 161: 225-229
Cruz J. C., Reeves J. T., Russell B. E., Alexander A. F., Will D. H. 1980. Embryo transplanted calves: the pulmonary hypertensive trait is genetically transmitted. Proc Soc Exp Biol Med, 164: 142-145
Damert A., Ikeda E., Risau W. 1997. Activator-protein-1 binding potentiates the hypoxia-inducible factor-1-mediated hypoxiainduced transcriptional activation of vascular-endothelial growth factor expression in c6 glioma cells. Biochem J, 327: 419-423
Daszak P., Berger L., Cuningham A. A., Hyatt A., Green D. E.,Spear R. 1999. Emerging infectious diseases and amphibian population declines. Emerg infect dis, 5: 735-748
Daszak P., Cunningham A. A., Hyatt A. D. 2003. Infectious disease and amphibian populations declines. Divers Distrib, 9:141-150
Drew A., Allen E. J., Allen L. J. S. 2006. Analysis of climatic andgeographic factors affecting the presence of chytridiomycosis in Australia. Dis Aquat Organ, 68: 245-250
Espinoza J. R., Alvarez G., LeÓn-Velarde F., Preciado H. F. J., Macarlupu J., Rivera-Ch M., Rodriguez J., Favier J.,Gimenez-Roqueplo A., Richalet J. 2014. Vascular Endothelial Growth Factor-A is associated with chronic mountain sickness in the Andean population. High Alt Med Biol, 15:146-154
Evans W. H., Wilson S. M., Bednarek J. M., Peterson E. A.,Knight R. D., Mage M. G., McHugh L. 1989. Evidence for a factor in normal human serum that induces human neutrophilic granulocyte end-stage maturation in vitro. Leuk Res, 13: 673-682
Forsythe J. A., Jiang B. H., Iyer N. V., Agani F., Leung S. W.,Koos R. D., Semenza G. L. 1996. Activation of vascular endothelial growth factor gene transcription by hypoxiainducible factor 1. Mol Cell Biol, 16: 4604-4613
Ge R. L., Cai Q., Shen Y. Y., San A., Ma L., Zhang Y., Yi X.,Chen Y., Yang L. F., Huang Y. 2013. Draft genome sequence of the Tibetan antelope. Nat Commun 4, 1858 | DOI: 10.1038/ ncomms2860
Giordano F. J. 2005. Oxygen, oxidative stress, hypoxia, and heart failure. J Clin Invest, 115: 500-508
Gracey A. Y., Troll J. V. Somero G. N. 2001. Hypoxia-induced gene expression profiling in the euryoxic fish Gillichthys mirabilis. Proc Natl Acad Sci USA, 98:1993-1998
Green D. E., Converse K. A., Schrader A. K. 2002 Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996-2001. Ann NY Acad Sci, 969: 323-339
Hu S. Q. 1987. Amphibia-reptilia in Tibet. Beijing: Science Press
Hutchison V. H., Haines H. B., Engbretson G. 1976. Aquatic life at high altitude: respiratory adaptations in the lake Titicaca frog,Telmatobius coleus. Respir Physiol, 27: 115-129
Kane N. C., Rieseberg L. H. 2007. Selective sweeps reveal candidate genes for adaptation to drought and salt tolerance in common sunflower, Helianthus annuus. Genetics, 175: 1823-1834
Knickerbocker D. L., Lutz P. L. 2001. Slow ATP loss and the defense of ion homeostasis in the anoxic frog brain. J Exp Biol,204: 3547-3551
Li Q. F., Sun R. Y., Huang C. X., Wang Z. K., Liu X. T., Hou J. J., Liu J. S., Cai L. Q., Li N., Zhang S. Z., Wang Y. 2001. Cold adaptive thermogenesis in small mammals from different geographical zones of China. Comp Biochem Physiol A, 129:949-961
Li H., Ren Y., Guo S., Cheng L., Wang D., Yang J., Chang Z.,Zhao X. 2009. The protein level of hypoxia-inducible factor-1α is increased in the plateau pika (Ochotona curzoniae) inhabiting high altitudes. J Exp Zool, 311A: 134-141
Li H., Guo S., Ren Y., Wang D., Yu H., Li W., Zhao X., Chang Z. 2013. VEGF189expression is highly related to adaptation of the plateau pika (Ochotona curzoniae) inhabiting high altitudes. High Alt Med Biol, 14: 395-404
Lip K., Brem F., Brenes R., Reeve J. D., Alford R. A. Voyles J.,Carey C., Livo L., Pessier A. P., Collins J. P. 2006. Emerging infectious disease and the loss of biodiversity in a neotropical amphibian community. Proc Natl Acad Sci USA, 103: 3165-3170.
Livak K. J., Schmittgen T. D. 2001. Analysis of relative gene expression data using realtime quantitative PCR and theMethod. Methods, 25: 402-408
Lu Z. K., Zhai L., Wang H., Che Q., Wang D., Feng F., Zhao Z., Yu H. 2010. Novel families of antimicrobial peptides with multiple functions from skin of Xizang plateau frog, Nanorana parkeri. Biochimie, 92: 475-481
Ma X. Y., Lu X., Merilä J. 2009. Altitudinal decline of body size in a Tibetan frog. J Zool, 279: 364-371
Ma X. Y., Lu X. 2009. Sexual size dimorphism in relation to age and growth based on skeletochronological analysis in a Tibetan frog. Amphib Reptil, 30: 351-359
Ma X. Y., Lu X. 2010. Annual cycle of reproductive organs in a Tibetan frog, Nanorana parkeri. Anim Biol, 60: 259-271
Mohd-Padil H., Mohd-Adnan A, GabaldÓn T. 2012. Phylogenetic analyses uncover a novel clade of transferrin in nonmammalian vertebrates. Mol Biol Evol, doi:10.1093/molbev/ mss325
Moore L. G., Armaza F., Villena M., Vargas E. 2000. Comparative aspects of high-altitude adaptation in human populations. Adv Exp Med Biol, 475: 45-62
Morabito M. A., Moczydlowski E. 1994. Molecular cloning of bullfrog saxiphilin: a unique relative of the transferrin family that binds saxitoxin. Proc Natl Acad Sci USA, 91: 2478-2482
Moskaitis J. E., Pastori R. L., Schoenberg D. R. 1990. The nucleotide sequence of Xenopus laevis transferrin mRNA. Nucleic Acids Res, 18: 6135
Nor J. E., Christensen J., Mooney D. J., Polverini P. J. 1999. Vascular endothelial growth factor (VEGF)-mediated angiogenesis is associated with enhanced endothelial cell survival and induction of Bcl-2 expression. Am J Pathol, 154:375-384
Qiu Q., Zhang G., Ma T., Qian W., Wang J., Ye Z., Cao C.,Hu Q., Kim J., Larkin D. M., Auvil L., Capitanu B., Ma J.,Lewin H. A., Qian X., Lang Y., Zhou R., Wang L., Wang K.,Xia J., Liao S., Pan S., Lu X., Hou H., Wang Y., Zang X., Yin Y., Ma H., Zhang J., Wang Z., Zhang Y., Zhang D., Yonezawa T., Hasegawa M., Zhong Y., Liu W., Zhang Y., Huang Z.,Zhang S., Long R., Yang H., Wang J., Lenstra J. A., Cooper D. N., Wu Y., Wang J., Shi P., Wang J., Liu J. 2012. The yak genome and adaptation to life at high altitude. Nature Genet, 44:946-949
Rissane E., Tranberg H. K., Sollid J., Nilsson G. E., Nikinmaa M. 2006. Temperature regulates hypoxia-inducible factor-1(HIF-1) in a poikilothermic vertebrate, crucian carp (Carassius carassius). J Exp Biol, 209: 994-1003
Rose M. R. 2001. Adaptation. In Levin RA (Eds), Encyclopedia of Biodiversity. San Diego: Academic Press: 17-23
Sahoo P. K., Mohanty B. R., Kumari J., Barat A., Sarangi N. 2009. Cloning, nucleotide sequence and phylogenetic analyses,and tissue-specifi c expression of the transferrin gene in Cirrhinus mrigala infected with Aeromonas hydrophila. Comp Immunol Microb, 32: 527-537
Simonson T. S., Yang Y., Huff C. D., Yun H., Qin G.,Witherspoon D. J., Bai Z., Lorenzo F. R., Xing J., Jorde L. B., Prchal J. T., Ge R. L. 2010. Genetic evidence for highaltitude adaptation in Tibet. Science, 329: 72-75
Soitamo A. J., Rabergh C. M., Gassmann M., Sistonen L.,Nikinmaa M. 2001. Characterization of a hypoxia-induciblefactor (HIF-1α) from rainbow trout. Accumulation of protein occurs at normal venous oxygen tension. J Biol Chem, 276:19699-19705
Stafford J. L., Belosevic M. 2003. Transferrin and innate immune response of fish: identification of a novel mechanism of macrophage activation. Dev Comp Immunol, 27: 539-554
Stewart E. R., Reese S. A., Ultsh G. R. 2004. The physiology of hibernation in Canadian leopard frogs (Rana pipiens) and bullfrogs (Rana catesbeiana). Physio Biochem Zool, 77: 65-73
Sun Y. B., Xiong Z. J., Xiang X. Y., Liu S. P., Zhou W. W., Tu X. L., Zhong L., Wang L., Wu D. D., Zhang B. L., Zhu C. L., Yang M. M., Chen H. M., Li F., Zhou L., Feng S. H.,Huang C., Zhang G. J., Irwin D., Hillis D. M., Murphy R. W., Yang H. M., Che J., Wang J., Zhang Y. P. 2014. Wholegenome sequence of the Tibetan frog Nanorana parkeri and the comparative evolution of tetrapod genomes. Proc Natl Acad Sci USA, 112: E1257-E1262
Tacchini L., Bianchi L., Bernelli-Zazzera A., Cairo G. 1999. Transferrin Receptor Induction by Hypoxia: HIF-1-Mediated transcriptional activation and cell-specific post-transcriptional regulation. J Biol Chem, 274: 24142-24146
Weber R. E., Ostojic H., Fago A., Dewilde S., Van Hauwaert M. L., Moens L., Monge C. 2002. Novel mechanism for high-altitude adaptation in hemoglobin of the Andean frog Telmatobius peruvianus. Am J of Physiol-Regul Integr Comp Physiol, 283: 1052-1060
Wenger R. H., Gassmann M. 1997. Oxygen (es) and the hypoxiainducible factor-1. Biol Chem, 378: 609-616
William G. K., Peter J. R. 2008. Oxygen sensing by metazoans:the central role of the HIF hydroxylase pathway. Mol Cell, 30:393-402
Wolf J. B. W. 2013. Principles of transcriptome analysis and gene expression quantifi cation: an RNA-seq tutorial. Mol Ecol Resour, 13: 559-572
Woodhams D. C., Alford R. A. 2005. Ecology of chytridiomycosis in rainforest stream frog assemblages of tropical Queensland. Conserv Biol, 19: 1449-1459
Wu T., Kayser B. 2006. High altitude adaptation in Tibetans. High Alt Med Biol, 7: 193-208
Xue Y., Petrovic N., Cao R., Larsson O., Lim S., Chen S.,Feldmann H. M., Liang Z. Zhu Z., Nedergaard J., Cannon B., Cao Y. 2009. Hypoxia-independent angiogenesis in adipose tissues during cold acclimation. Cell Metabol, 9: 99-109
Zhang L. X., Ma X. Y., Jiang J. P., Lu X. 2012. Stronger condition dependence in female size explains altitudinal variation in sexual size dimorphism of a Tibetan frog. Biol J Linnean Soc, 107:558-565
*
Dr. Qiong Zhang, from Institute of Zoology,Chinese Academy of Sciences, Beijing, China, with her research focusing on the molecular ecology and adaptive evolution of amphibians.
E-mail: zhangqiong@ioz.ac.cn
29 June 2015 Accepted: 1 December 2015
 Asian Herpetological Research2016年1期
Asian Herpetological Research2016年1期
- Asian Herpetological Research的其它文章
- Allelic Polymorphism, Gene Duplication and Balancing Selection of MHC Class IIB Genes in the Omei Treefrog (Rhacophorus omeimontis)
- The Expression Plasticity of Hypoxia Related Genes in High-Altitude and Plains Nanorana parkeri Populations
- Soundscape Dynamics at Anuran Reproductive Sites in Pannonian Biogeographical Region: Effects of Road Noise on Vocal Activity
- Effects of Pesticide Exposure on Embryonic Development and Hatchling Traits of Turtles
- No Evidence for Signifi cant Effect of Body Size and Age on Male Mating Success in the Spot-legged Treefrog
- Effects of Predation by Invasive Western Mosquitofi sh (Gambusia affi nis) on Survival of Eggs, Embryos and Tadpoles of Pelophylax nigromaculatus and Duttaphrynus melanostictus in South China