
气候变化下栓皮栎潜在地理分布格局及其主导气候因子
2016-09-21 02:34:15高文强王小菲江泽平刘建锋
生态学报 2016年14期
高文强,王小菲,江泽平,2,刘建锋,2,*
1 中国林业科学研究院林业研究所,北京 100091 2 国家林业局林木培育重点实验室,北京 100091
气候变化下栓皮栎潜在地理分布格局及其主导气候因子
高文强1,王小菲1,江泽平1,2,刘建锋1,2,*
1 中国林业科学研究院林业研究所,北京1000912 国家林业局林木培育重点实验室,北京100091
栓皮栎(Quercusvariabilis)是东亚天然分布最广泛的树种之一。利用最大熵(Maxent)模型对现实气候条件下栓皮栎在东亚地区的潜在分布及其分布的主导气候因子进行分析,同时结合全新世中期(6000年前)和未来气候(2050年)来模拟和预测气候变化背景下栓皮栎潜在分布格局的变化。结果表明:现实气候条件下东亚栓皮栎适生区(适宜生境和低适宜生境)面积占总研究区面积的21.88%,主要集中在东亚南部区域,在我国北起陕西中部、山西和河北南部边缘、山东,西起甘肃东部边缘、四川中东部、云南、西藏东部边缘,一直到东部沿海区域,同时在朝鲜半岛南部和日本中南部也有分布,其中适宜生境面积占研究区总面积的5.69%,主要集中在秦岭山脉、大巴山脉、伏牛山、云南的云贵高原、罗霄山脉、南岭山脉、武夷山和台湾岛;气候变化情景下,栓皮栎的适生区分布面积变化较小,但其适宜生境的分布范围却发生了较大的变化,随着全球气候的波动性变化,适宜生境分布范围逐渐向西部秦岭山脉、大巴山脉、四川、重庆和云贵高原等区域集中,并使该分布中心的适宜生境面积逐渐扩大;影响栓皮栎分布的主要气候因子为最冷月的最低气温(Bio6)、最冷季平均气温(Bio11)和年降水量(Bio12),三者的贡献率分别为48.6%、21.4%和14.2%。
栓皮栎;气候变化;Maxent模型;分布格局
植被与气候之间相互关系的研究一直是植物学、生态学以及地理学等领域研究的热点问题。气候是决定地球上物种分布的最主要因素,而物种分布格局的变化最能直接和间接的反映气候变化[1]。目前全球气候变化已是无需争辩的事实,虽然众多科学家依据不同模型预测的结果不尽一致,但目前人们普遍认为,由于大气臭氧层的不断破坏以及温室气体的持续增加,全球的平均气温也将上升。IPCC[2-3]的研究表明,1906—2005年全球地表平均温度升高(0.74 ± 0.18)℃,预计到21世纪末地表平均温度将上升1.1—6.4℃。全新世是与人类关系最密切的一个时期,经历了全新世早期的升温阶段,进入全新世中期大暖期,平均温度高于现代1.5℃[4]。随着全球气候变化,生物的适宜分布区也将发生改变,因而预测气候变化对物种潜在分布格局的影响已成为生物地理学和全球变化研究的热点和重点领域[5-6]。
物种分布模型是基于物种的已知分布及其环境变量,来评估气候变化对物种潜在地理分布的影响[7-8]。目前得到广泛应用的模型,包括生态位模型(Bioclim、DIVA、Domain)、基于遗传算法的规则组合预测模型(genetic algorithm for ruleset production, GARP)、动态模拟模型(CLIMEX)、广义相加模型(GAM)、广义线性模型(GLM)以及最大熵模型(maxi-mum entropy,Maxent)等[8-11]。经许多研究证明,最大熵模型(Maxent)是目前对物种分布应用最广泛且具较好预测能力的模型[8-9,12-17]。最大熵模型是以最大熵理论为基础,通过物种的已知分布数据和环境数据找出物种概率分布的最大熵,从而对物种的地理分布进行估计和预测[18]。
栓皮栎(Quercusvariabilis)隶属于壳斗科栎属,是东亚天然分布最广泛的树种之一[19]。栓皮栎分布地理区域包括中国大陆、台湾岛和海南岛以及日本列岛和朝鲜半岛[20-23],其分布气候区横跨温带、暖温带、北亚热带和南亚热带,在各区以纯林和混交林的森林群落存在[24]。因此,栓皮栎是研究植物分布与气候、植物对气候变化响应的理想树种之一。另外,栓皮栎具有抗旱、耐贫瘠、对保持水土、涵养水源、增加土壤肥力等作用,不仅发挥着重要的生态效益,还具有潜在经济效益,因而,在林业生产和生态环境保护方面起着重要作用。有关栓皮栎的研究主要涉及其生态学习性、种群动态、种群生物学、群落特征、群落物种多样性和生态位[25-29];而关于地理分布以及气候特征,前人做过一些研究[24],但对于气候变化下栓皮栎的地理分布现状及其与气候的关系,以及在全新世中期、现实气候和未来气候变化情景下其适宜的分布范围和空间格局发生怎样的改变等问题尚未见过相关的研究报道。因此,本文基于大量文献和标本采集记录,结合GIS技术,利用最大熵(Maxent)模型模拟当前气候情形下东亚栓皮栎的潜在分布格局并综合分析影响其分布的主导因子;结合全新世中期和未来气候情景,来分析气候变化背景下栓皮栎潜在分布格局的变化,这对栓皮栎有效经营管理以及应对气候变化具有十分重要的理论与现实意义。
1 材料和方法
1.1数据收集
栓皮栎在东亚(中国、中国台湾、日本、韩国和朝鲜)的分布记录来自中国数字化标本馆(http://www.cvh.ac.cn/)、CVH分布式数据检索(http://pe.ibcas.ac.cn/sptest/syninvok.aspx)文献,共收集了297个物种分布样点。
环境数据来源于世界气候数据库(WORLDCLIM, http://www.Worldclim.org)。该数据库采用插值法将全球气象站点数据(1950—2000)生成世界气候栅格数据,并衍生19个生物气候变量(Bioclim variables)[30-33](表1)。本文利用该数据库中的3期气候数据,分别为全新世中期(Mid Holocene,6000年前),现实气候(1950—2000年)及未来气候情景(2050年),空间分辨率为30″。未来气候(2050年)选用了两个不同温室气体排放场景(rcp45和rcp85)[34],其中以rcp45和rcp85分别代表了2050年全球平均增温的最高(1.4℃)与最低值(2.0℃)。本研究对选取的19个生物气候变量利用ArcGIS软件将研究区数据裁剪出来,得到各气候环境图层数据。地图资料源自DIVA-GIS网站(http://www.diva-gis.org/)下载全球地理信息资料和国家基础地理信息中心(http://ngcc.sbsm.gov.cn/)下载1∶400万的中国行政区划图,提取东亚地图作为底图。
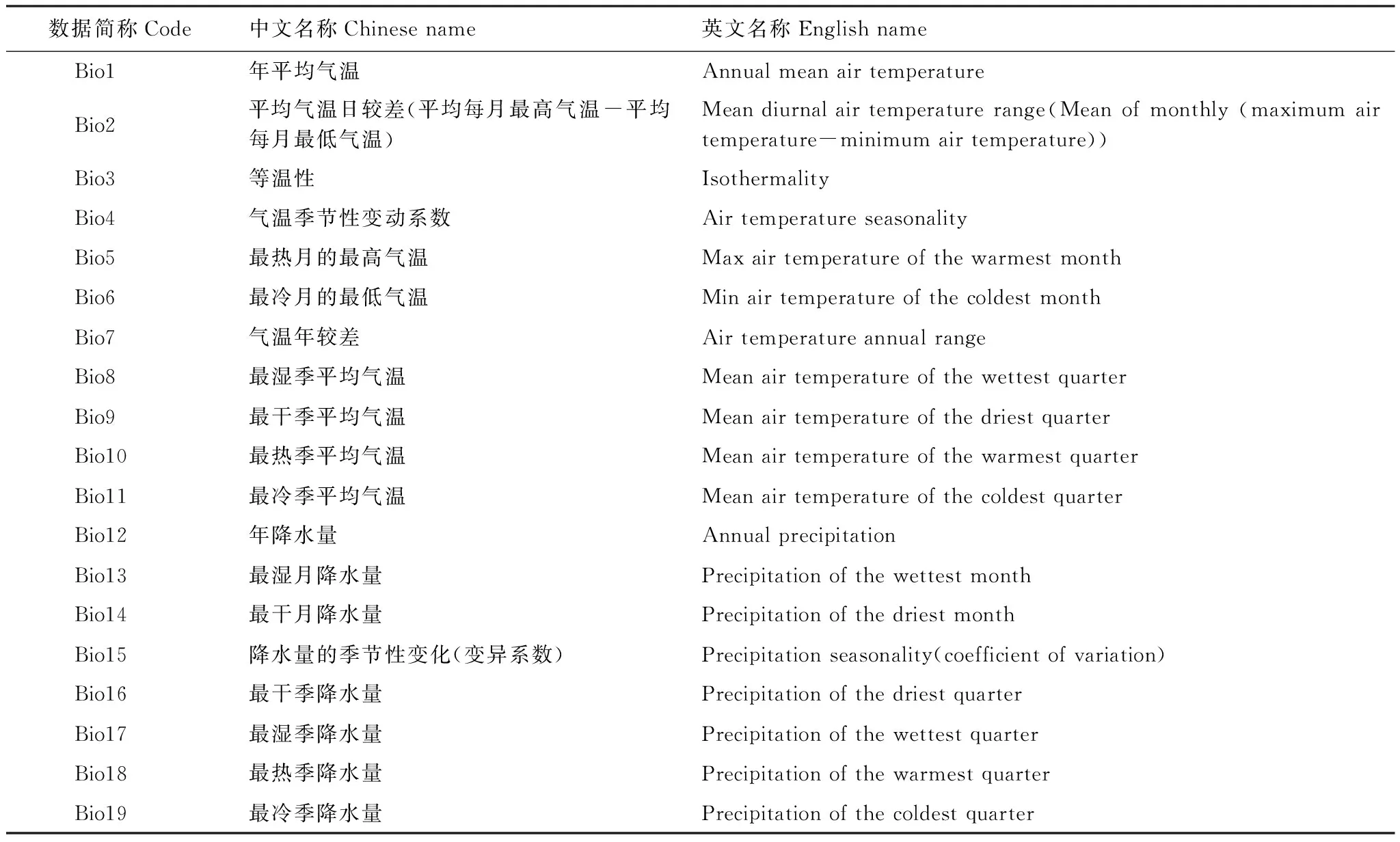
表1 19个生物气候变量
1.2模型精度验证
本研究选取Maxent模型(V3.3.3k)进行栓皮栎气候适应性研究。首先,确定物种特定空间分布区域,随机选取全部数据的75%作为训练集(training data)来训练模型,将剩余25%的数据用作评估子集(testing data)来验证模型。其次,寻找限制物种分布潜在气候因子构筑约束集合。第三,利用Maxent模型构建栓皮栎的地理分布与气候关系模型[35],并采用接受者操作特性曲线(receiver operating characteristic curve, ROC)分析法进行模型精度检验,ROC曲线基于非阈值依赖判断(threshold-independent evaluation)模型精度,即是以预测结果的每一个值作为可能的判断阈值,由此计算得到相应的灵敏度和特异度[35]。以特异度(1-specificity)即假阳性率为横坐标,以灵敏度(1-omission rate)即真阳性率为纵坐标绘制成ROC曲线,AUC值是ROC曲线与横坐标围成的面积值(the area under the ROC curve),其大小能很好地说明模型模拟值的准确性,取值范围为[0,1],AUC值越接近1,说明与随机分布相距越远,环境变量与预测的物种地理分布之间的相关性越大,模型预测结果越准确[36]。由于AUC不受诊断阈值的影响,且对物种发生率不敏感,因此目前被公认为是最佳的评价指标[37]。ROC曲线的评估标准[35, 38-40]为: AUC为0.5—0.6,预测结果不可接受(fail);AUC为0.6—0.7,预测结果勉强可以接受(poor);AUC为0.7—0.8,预测结果一般(fair),可以接受;AUC为0.8—0.9,预测结果较为满意(good);AUC为0.9—1.0,预测结果非常满意(excellent)。第四,基于各潜在气候因子对栓皮栎地理分布的贡献,采用模型的刀切法(Jackknife)模块来评价各环境因子的权重,筛选影响研究区栓皮栎地理分布的主导气候因子。第五,基于ArcGIS地理信息系统,得到Maxent模型模拟东亚栓皮栎的地理分布。
1.3适生区预测与划分
本文利用Maxent软件结合栓皮栎在东亚的分布数据和环境数据,对全新世中期(6000年前)、现实气候(1950—2000)以及21世纪50年代两个未来情景气候(CCSM4-rcp85和CCSM4-rcp45)的影响下东亚栓皮栎的分布进行了模拟,在ArcGIS 10.0中对生成的ASCII格式数据转换为栅格数据文件,并进行重分类,按照适宜生境评价指数[1, 41-42],将研究区栓皮栎生境划分为3类;即不适宜生境、低适宜生境和适宜生境,其中不适宜生境的评价指数小于0.25,低适宜生境和适宜生境的评价指数分别为0.25—0.50和0.50—1.00。同时经ArcGIS 10.0空间分析模块叠加显示,并计算全新世中期、现实气候和未来两种情景气候条件下各生境分布区占研究区总面积的比率,得出研究区栓皮栎在气候变化下的潜在空间分布结果。
2 结果与分析
2.1模型模拟结果的准确性评价
本文基于Maxent模型模拟栓皮栎全新世中期、现实和未来情景的潜在分布,采用ROC曲线作为模型预测准确性的衡量指标(图1),其训练集和验证集AUC值均大于0.88,表2。这表明所构建模型的预测准确性较好,可以用于东亚栓皮栎地理分布与气候关系的研究。
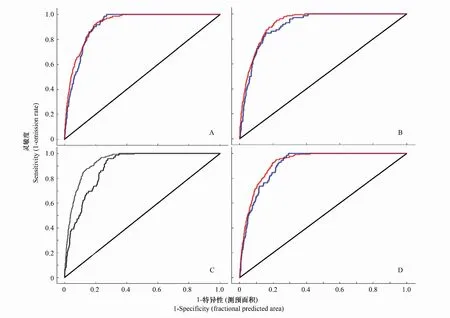
图1 栓皮栎模型预测ROC曲线Fig.1 The predicted ROC curve by Quercus variabilis model红线表示训练集,蓝线表示验证集,黑线表随机预测; A:全新世中期; B: 现实气候; C和D: 未来气候情景CCSM4-rcp45和CCSM4-rcp85
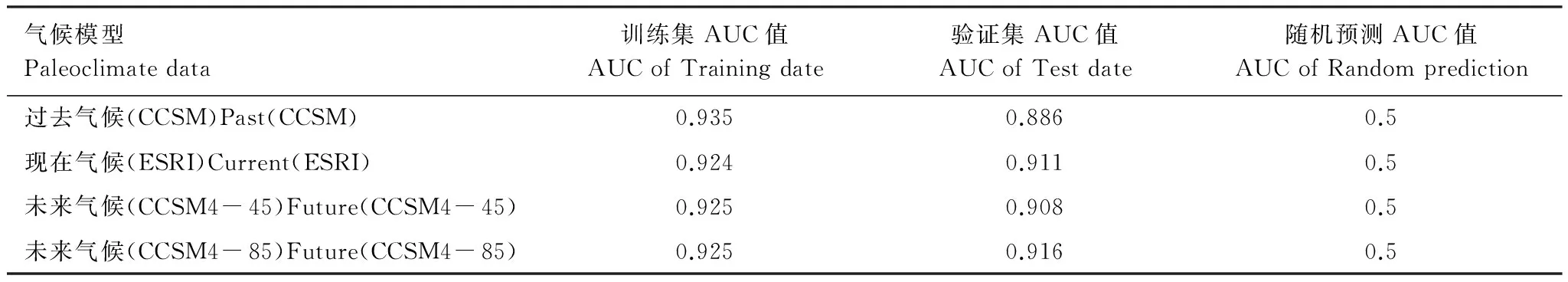
气候模型Paleoclimatedata训练集AUC值AUCofTrainingdate验证集AUC值AUCofTestdate随机预测AUC值AUCofRandomprediction过去气候(CCSM)Past(CCSM)0.9350.8860.5现在气候(ESRI)Current(ESRI)0.9240.9110.5未来气候(CCSM4-45)Future(CCSM4-45)0.9250.9080.5未来气候(CCSM4-85)Future(CCSM4-85)0.9250.9160.5
2.2气候变化下栓皮栎的潜在分布格局
如表3所示,随着时间的变化,由全新世中期到现实气候,东亚栓皮栎的总适生区(低适宜生境和适宜生境)面积呈减小的趋势。然而,未来气候情景CCSM4-rcp45(2050)栓皮栎总适生区面积与CCSM4-rcp85模拟的结果稍有不同,其适生区面积比全新世中期、现实气候条件下略有增大,而后者表现为缩小。在历史气候条件下,东亚栓皮栎适宜生境和低适宜生境的分布面积分别占研究区总面积的6.22%和16.62%;与历史气候条件相比,当前气候条件下,不论是低适宜生境,还是适宜生境,栓皮栎的分布面积均略有减小,分别占总面积的5.69%和16.19%。未来气候情景(CCSM4-rcp45和CCSM4-rcp85)下适宜生境面积均比过去和当前气候条件下大,分别占总面积的6.77%和6.42%;但在低适宜生境方面,CCSM4-rcp85情景拟合的面积比过去和当前气候条件下的小,占研究区总面积的15.31%,而CCSM4-rcp45情景下则比过去和当前气候条件下分布面积大,占研究区总面积的16.46%。就未来气候情景而言,与较小的增温幅度相比,增温幅度大(rcp85)会导致总适生区面积(包括低适宜生境面积和适宜生境面积)减小。

表3 气候变化情景下栓皮栎适生区分布面积百分比
虽然在全新世中期、现实气候和未来气候情景下栓皮栎总适生区(适宜生境和低适宜生境)面积变化较小,但适宜生境的分布范围却发生了较大的变化,如图2。模拟的全新世中期东亚栓皮栎适生区主要位于东亚南部,在我国北起陕西中部、山西和河北南部边缘、山东,西起甘肃东部边缘、四川中东部、云南、西藏东部边缘,一直到达东部沿海区域,另外在河北和北京交界也有少量适生区的存在;在朝鲜半岛和日本主要分布于南部和中南部。其中适宜生境主要位于陕西中南部、山西南部边缘、河南中西部、甘肃东南部、四川东北和东南部、西藏东南边缘、重庆、湖北、湖南、云南、贵州、广西西部、江西、安徽中部、江苏西南缘、浙江东北部、福建和台湾中部,以及朝鲜半岛东南缘和日本中南部也有少量适宜生境的存在。
如图2,与全新世中期相比,现实气候条件下,在朝鲜半岛的适宜生境分布区消失了,同时位于我国东部包括台湾岛、江苏、浙江、江西、河南和湖南以及日本的适宜生境区面积也有不同程度的减小;在全新世中期,北京与河北的交界处以及海南岛均没有适宜生境的存在,而现实气候条件下出现了适宜生境,同时位于云南、四川、重庆以及陕西地区的适宜生境的分布面积均略有增大。在未来情景下(21世纪50年代),CCSM4-rcp85模型预测我国河南和湖北以及日本的栓皮栎适宜生境分布面积将比现实气候下进一步减少,而云南、贵州、四川和陕西南部等区域有进一步增加的趋势。与前者稍有不同的是,CCSM4-rcp45模型预测的日本中部区域其适宜生境分布面积在扩大。对于低适宜生境,两个模型预测的朝鲜半岛的分布区面积均略有减少,而广东和广西南部边缘面积均略增大。从总体来看,东亚栓皮栎的适宜生境的核心区位于我国云南、贵州、四川、陕西南部等区域,在未来情境下该核心区适宜生境分布面积有进一步扩大的趋势,而位于东南沿海地区的适宜生境分布面积有减少的趋势;在低适宜生境区,未来气候情景的两个模型预测的朝鲜半岛分布面积均稍有减少,而广东和广西南部边缘的面积均稍有增大,其余的分布范围变化较小。
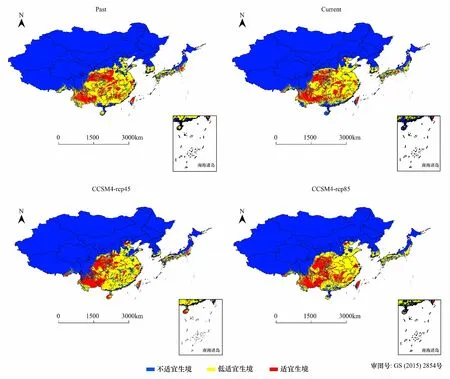
图2 不同气候变化情景下栓皮栎生境适应性分布Fig 2 Distribution of habitat suitability for Quercus variabilis under different climate change scenariosPast表示全新世中期气候情景(6000年前);Current表示现实气候条件(1950—2000);CCSM4-rcp85和CCSM4-rcp45表示21世纪50年代两个未来气候情景; Past: Last Glacial Maximum climate scenario(6000 years previous); Current: current climate scenario; CCSM4-rcp85 and CCSM4-rcp45 represent the climate scenario of 2050s, respectively
2.3影响栓皮栎分布区的主导气候因子
刀切法(Jackknife)常被用来分析各环境因子对预测结果的影响程度,或用来确定影响物种分布的关键性环境因素[43]。根据刀切法分析得到的环境因子对最大熵模型的贡献率,如表4,可以判断影响物种分布的主要环境因子。结果表明:最冷月的最低气温贡献率(bio6)最高为48.6%,最冷季平均气温(bio11)、年降水量(bio12)贡献率分别为21.4%和14.2%,最湿季降水量(bio17)、平均气温日较差(平均每月最高气温-平均每月最低气温)(bio2)和年平均气温(bio1)分别为3.2%、2.7%和2.3%。这6个因子的累积贡献率高达92.4%。综合分析表明,最冷月的最低气温和最冷季平均气温是影响栓皮栎分布最主要的环境因子,其次是年降雨量,最湿季降水量、平均气温日较差和年平均气温对栓皮栎的分布也一定的影响,但较最冷月气温和年降水量小。
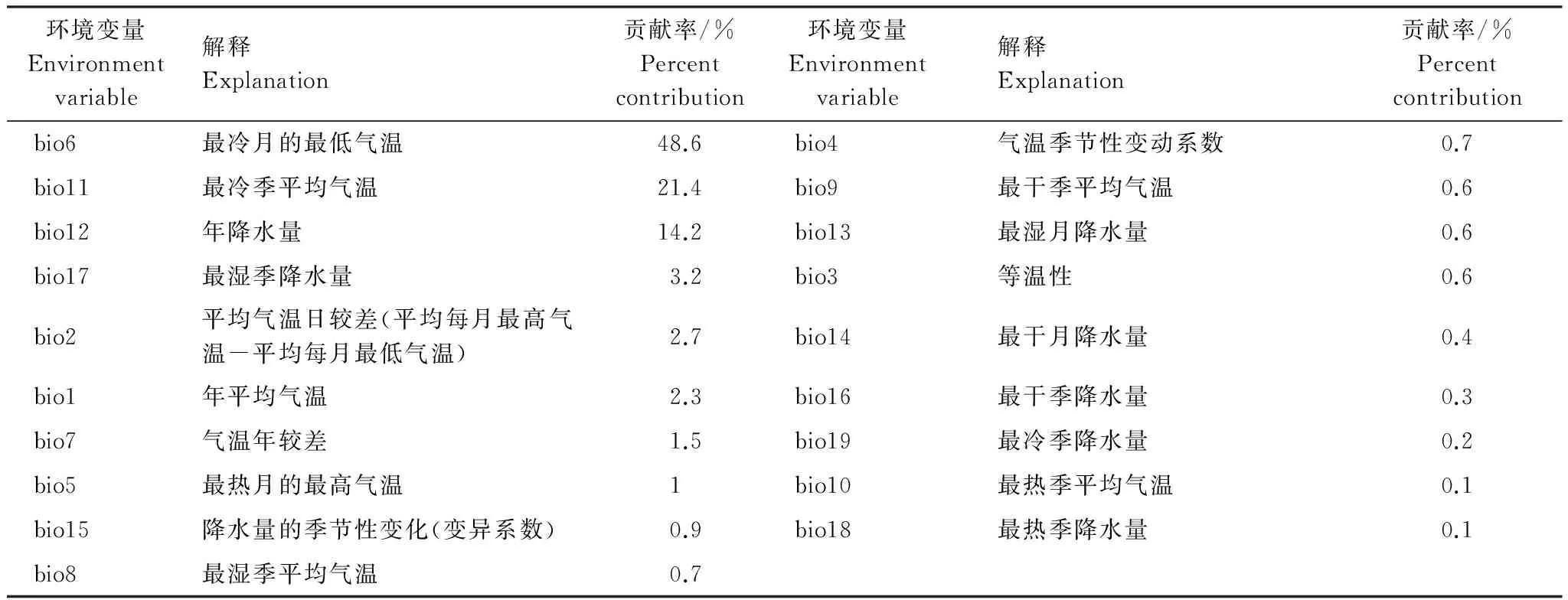
表4 最大熵模型中各环境变量贡献率
3 讨论与结论
应用Maxent模型对气候变化下物种潜在分布预测的研究已有报道,主要针对能源植物[44]、药用植物[45]、入侵物种[35, 40]、濒危物种[45]以及物种的迁移路线[12, 46]等所做的相关研究,本文首次利用Maxent生态位模型与地理信息系统相结合,对广布于东亚且对林业生产和生态环境保护具有重要价值的栓皮栎在气候变化下的潜在分布进行了预测,获得了不同气候条件下栓皮栎在东亚的潜在适生分布图。经检验,Maxent模型的受试者工作特征曲线(ROC曲线)下面积AUC值均接近于1,表明预测结果较好。
Maxent模型模拟结果表明,在当前气候条件下,栓皮栎适生区(低适宜生境和适宜生境)主要位于东亚东南区域,包括我国辽宁南部边缘、北京和河北交界的燕山山脉、山东、山西和河北南部边缘、陕西中南部、甘肃东部、河南、安徽、江苏、上海、浙江、福建、湖北、湖南、广东、广西、云南、贵州、重庆、四川中东部、西藏南部边缘以及海南岛和台湾岛,朝鲜半岛南部和日本中南部。这与前人所研究调查的地理分布大致相同[19,24,47-48],表明经Maxent模型模拟结果与实际分布相符。其中,适宜生境主要位于我国陕西中南部、甘肃东部、四川东北部、重庆、河南和湖北东部的秦岭山脉和大巴山脉、云贵高原、以及位于江西和湖南交界的罗霄山脉、福建的武夷山、湖南、江西和广东的南岭山脉以及海南岛东部边缘和台湾岛中部,另外日本中部与南部也有较小的适宜生境存在。关于栓皮栎的分布中心,目前存在较多的争论,吴征镒[49]认为大别山、桐柏山和秦岭;郑万钧[50]认为在鄂西、秦岭和大别山区;傅焕光认为在秦岭、伏牛山、桐柏山、大别山、鄂西和川东一带。而本研究显示秦岭、桐柏山、伏牛山、大巴山、鄂西和云南的云贵高原是其最适宜分布区,而安徽的大别山不属于其最适宜分布区[51-52]。
在气候变化情景下,全新世中期栓皮栎的总适生区(低适宜生境和适宜生境)分布范围基本与现实和未来气候情景下的分布范围一致;而其适宜生境面积却发生了较大的变化。随着全球气候变化,适宜生境范围逐渐向西部秦岭山脉、大巴山脉、四川、重庆和云贵高原等区域集中,使该分布中心的适宜生境面积在逐渐地扩大。与当前气候相比,全新世中期分布区最北的北京、天津和河北等地没有适宜生境,而当前气候条件出现了栓皮栎的分布,这可能是由于全新世中期我国北方气温波动较剧烈,不断经历温度的冷暖交替[53],难以形成其适宜分布区,同时该区域又属于栓皮栎分布北界,对温度变化响应较敏感。而到当前气候条件下,气候波动较小,存在栓皮栎的适宜生境,甚至到未来气温升高的情景下,该区域的适宜生境面积进一步扩大。另外,海南岛在全新世中期没有适宜生境,而到当前气候条件下出现了小面积的适宜生境,这可能与随着全球气温的降低(该时期在我国南方温度波动较小[54]),物种分布范围向南迁移有关。然而经调查发现,目前海南岛没有栓皮栎的分布,仅在中科院华南植物园标本馆资料中有记录[24],这可能是由于人类活动的影响所致。但在其他研究区域未见到明显的纬向迁移,这可能是由于栓皮栎主要生长在山区,且研究区山地较多,地形复杂多变,而气候本身也随地区的不同而变化很大,使得该区域栓皮栎分布范围无法纬向迁移。另外,极端气候也是物种分布范围变化的重要助推剂,因为极端气候事件的不连续性,导致物种分布范围的变化通常也是非连续性的[41],而本研究表明极端气候(bio6和bio11)是栓皮栎分布的主导因子,因此未来气候变化对栓皮栎适宜生境的影响也较为复杂。根据本文气候因子贡献率的分析结果,Bio6(最冷月的最低气温)和Bio11(最冷季平均气温)评价因子贡献率之和为70%,说明冬季极端低温是影响栓皮栎适生区分布的决定性因子。然而,不同地理区域存在一定差异。王婧等[24]对东亚栓皮栎的气候特征研究结果表明,在大陆地区栓皮栎分布区的最冷季均温为-9.8—18℃,而台湾岛为-0.2—21.2℃,日本则为-3.8—6.2℃。同时评价因子年降水量(Bio12)的贡献率达到14.2%,说明降水也会在一定程度上影响栓皮栎适生区的分布。这与王婧[24]等的研究结果一致,即栓皮栎的分布范围受降水量影响,也与本文关于栓皮栎适生区止于西北甘肃和四川东部区域等降水线相吻合。
本文基于气候因素预测了在过去(6000年前)、当前(1950—2000)以及未来气候情景下(2050)东亚栓皮栎适生区(适宜生境和低适宜生境)分布的变化趋势,并识别了影响栓皮栎分布的主导气候因子。在未来气候情景下,适宜生境范围逐渐向西部秦岭山脉、大巴山脉、四川、重庆和云贵高原等区域集中,并使该分布中心的适宜生境面积在逐渐地扩大。同时极端低温(最冷月的最低气温和最冷季平均气温)和年降水量作为栓皮栎分布的决定性因子,这对于栓皮栎在林业生产以及有效的经营管理方面具有十分重要参考价值。该研究结果支持Petit等[55]和Désamoré等[56]的假设:弄清楚物种在过去气候变化条件下的分布格局的变化,能够预测它们将会如何应对不断发生的未来气候的变化。除气候因素外,影响物种分布的因素还很多,包括生物因素和非生物因子,其中生物因素包括因物种进化而带来的其对环境因子适应性的改变,以及物种种间竞争对物种分布区的影响等,非生物因素包括地形和土壤等,由于目前的认识水平以及技术条件的不足,因此本文未做考虑。
[1]郭彦龙, 卫海燕, 路春燕, 张海龙, 顾蔚. 气候变化下桃儿七潜在地理分布的预测. 植物生态学报, 2014, 38(3): 249-261.
[2]IPCC(Intergovernmental Panel on Climate Change). The Physical Science Basis Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007.
[3]IPCC(Intergovernmental Panel on Climate Change). Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007.
[4]姚檀栋, Tompson L G, 施雅风, 秦大河, 焦克勤, 杨志红, 田立德, L G Thompson, E Mosley-Thompson. 古里雅冰芯中末次间冰期以来气候变化记录研究. 中国科学(D), 1997, 27(5): 447-452.
[5]Céline Bellard, Cleo Bertelsmeier, Paul Leadley, Wilfried Thuiller, Franck Courchamp. Impacts of climate change on the future of biodiversity. Ecology Letters, 2012, 15(4): 365-377.
[6]Anahí Espíndola, Loïc Pellissier, Luigi Maiorano, Wim Hordijk, Antoine Guisan, Nadir Alvarez. Predicting present and future intra-specific genetic structure through niche hindcasting across 24 millennia. Ecology Letters, 2012, 15(7): 649-657.
[7]Antoine Guisan, Niklaus E, Zimmermann. Predictive habitat distribution models in ecology. Ecological Modelling, 2000, 135(2/3): 147-186.
[8]王娟, 倪健. 植物种分布的模拟研究进展. 植物生态学报, 2006, 30(6): 1040-1053.
[9]Steven J Phillips, Miroslav Dudik. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175.
[10]Jane Elith, Catherine H Graham, Robert P Anderson, Miroslav Dudík, Simon Ferrier, Antoine Guisan, Robert J Hijmans, Falk Huettmann, John R Leathwick, Anthony Lehmann, Jin Li, Lucia G Lohmann, Bette A Loiselle, Glenn Manion, Craig Moritz, Miguel Nakamura, Yoshinori Nakazawa, Jacob McC M Overton, A Townsend Peterson, Steven J Phillips, Karen Richardson, Ricardo Scachetti-Pereira, Robert E Schapire, Jorge Soberón, Stephen Williams, Mary S Wisz, Niklaus E Zimmermann. Novel methods improve prediction of species′ distributions from occurrence data. Ecography, 2006, 29(2): 129-151.
[11]Antoine Guisan, Wilfried Thuiller. Predicting species distribution: offering more than simple habitat models. Ecology Letters, 2005, 8(9): 993-1009.
[12]张雷, 刘世荣, 孙鹏森, 王同立. 不同立气候变化对马尾松潜在分布影响预估的多模型比较. 植物生态学报, 2011, 35(11): 1091-1105.
[13]Steven J Phillips, Robert P Anderson, Robert E Schapire. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3/4): 231-259.
[14]雷军成, 徐海根. 基于MaxEnt的加拿大一枝黄花在中国的潜在分布区预测. 生态与农村环境学报, 2010, 26(2): 137-141.
[15]王运生, 谢丙炎, 万方浩,肖启明,戴良英. 相似穿孔线虫在中国的适生区预测. 中国农业科学, 2007, 40(11):2502-2506.
[16]João G R Giovanelli, Célio F B Haddad, João Alexandrino. Predicting the potential distribution of the alien invasive American bullfrog (Lithobatescatesbeianus) in Brazil. Biological Invasions, 2008, 10(5): 585-590.
[17]杨波, 薛跃规, 唐小飞,王媛,张达敏. 外来入侵植物飞机草在中国的适生区预测. 植物保护, 2009, 35(4):70-73.
[18]Steven J Phillips, Miroslav Dudík, Robert E Schapire. A maximum entropy approach to species distribution modeling // Proceedings of the Twenty-First International Conference on Machine Learning. Banff Alberta, 2004: 655-662.
[19]魏林. 栓皮栎分布的初步调查. 林业科学, 1960, 6(1): 70-71.
[20]雷静品, 肖文发, 刘建锋. 我国栓皮栎分布及其生态学研究. 世界林业研究, 2013, 26(4): 57-62.
[21]Miyazaki Masahisa, Teramoto Noriyuki. Morphology and bionomics of the Japanese oak dwarflousePhylloxerakunugi(Homoptera, Aphidinea, Phylloxeridae). Entomologia Generalis, 1991, 16(3): 201-206.
[22]Mi Yoon Chung, Myong Gi Chung. Fine-scale genetic structure in populations ofQuercusvariabilisfrom southern Korea. Canadian Journal of Botany, 2002, 80(10): 1034-1041.
[23]Yeonsook Choung, Byung-Chun Lee, Jae-Hyoung Cho, Kyu-Song Lee, In-Soo Jang, Sun-Hee Kim, Sun-Kee Hong, Hui-Cheul Jung, Heung-Lak Choung. Forest responses to the large-scale east coast fires in Korea. Ecological Research, 2004, 19(1): 43-54.
[24]王婧, 王少波, 康宏樟, 辛在军, 钱振华, 刘春江. 东亚地区栓皮栎的地理分布格局及其气候特征. 上海交通大学学报: 农业科学版, 2009, 27(3): 235-241.
[25]吴明作, 刘玉萃. 栓皮栎种群数量动态的谱分析与稳定性. 生态学杂志, 2000, 19(4): 23-26, 37.
[26]吴明作, 姜志林, 刘玉萃. 栓皮栎种群的年龄动态与稳定性研究. 河南科学, 1999, 17(1): 69-73.
[27]吴明作, 杨玉珍, 杨玉珍, 李战军. 河南省栓皮栎林主要种群的生态位研究. 西北植物学报, 1999, 19(3): 511-518.
[28]程瑞梅, 肖文发. 河南宝天曼栓皮栎林群落特征及物种多样性. 植物资源与环境, 1998, 7(4): 8-13.
[29]张文辉, 卢志军. 栓皮栎种群的生物学生态学特性和地理分布研究. 西北植物学报, 2002, 22(5): 1093-1101.
[30]Nix H A. A biogeographic analysis of Australian elapid snakes // Longmore R, ed. Atlas of Australian Elapid Snakes. Canberra: Australian Government Publishing Service, 1986: 4-15.
[31]Pradeep Kumar. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using MaxEnt modelling: limitations and challenges. Biodiversity and Conservation, 2012, 21(5): 1251-1266.
[32]Robert J Hijmans, Susan E Cameron, Juan L Parra, Peter G Jones, Andy Jarvis. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 2005, 25(15): 1965-1978.
[30]William D Collins, Cecilia M Bitz, Maurice L Blackmon, Gordon B Bonan, Christopher S Bretherton, James A Carton, Ping Chang, Scott C Doney, James J Hack, Thomas B Henderson, Jeffrey T Kiehl, William G Large, Daniel S McKenna, Benjamin D Santer, Richard D Smith. The community climate system model version3 (CCSM3). Journal of Climate, 2006, 19(11): 2122-2143.
[34]James J McCarthy, Osvaldo F Canziani, Neil A Leary, David J Dokken, Kasey S White. Climate Change 2001: Impacts, Adaptation, and Vulnerability: Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change. New York: Cambridge University Press, 2001.
[35]王运生, 谢丙炎, 万方浩, 肖启明, 戴良英. ROC曲线分析在评价入侵物种分布模型中的应用. 生物多样性, 2007, 15(4): 365-372.
[36]Hanley J A, Mcneil B J. The meaning and use of the area under a Receiver Operating Characteristic (ROC) curve. Radiology, 1982, 143(1): 29-36.
[37]Alan H Fielding, John F Bell. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation, 1997, 24(1): 38-49.
[38]Swets J A. Measuring the accuracy of diagnostic systems. Science, 1988, 4857(240): 1285-1293.
[39]Giedirus Vanagas. Receiver operating characteristic curves and comparison of cardiac surgery risk stratification systems. Interactive Cardio Vascular and Thoracic Surgery, 2004, 3(2): 319-322.
[40]张颖, 李君, 林蔚, 强胜. 基于最大熵生态位元模型的入侵杂草春飞蓬在中国潜在分布区的预测. 应用生态学报, 2011, 22(11): 2970-2976.
[41]胡理乐, 张海英, 秦岭, 闫伯前. 中国五味子分布范围及气候变化影响预测. 应用生态学报, 2012, 23(9): 2445-2450.
[42]Lu C Y, Gu W, Dai A H, Wei H Y. Assessing habitat suitability based on geographic information system (GIS) and fuzzy: a case study ofSchisandrasphenantheraRehd. et Wils. in Qinling Mountains, China. Ecological Modelling, 2012, 242: 105-115.
[43]A Townsend Peterson, Kevin P Cohoon. Sensitivity of distributional prediction algorithms to geographic data completeness. Ecological Modelling, 1999, 117(1): 159-164.
[44]周婧, 李巧云, 肖亮, 蒋建雄, 易自力. 芒和五节芒在中国的潜在分布. 植物生态学报, 2012, 36(6): 504-510.
[45]车乐, 曹博, 白成科, 王娟娟, 张琳琳. 基于MaxEnt和ArcGIS对太白米的潜在分布预测及适宜性评价. 生态学杂志, 2014, 33(6): 1623-1628.
[46]于海彬, 张镱锂, 李士成, 祁威, 胡忠俊. 基于GIS和物种分布模型的高山植物长花马先蒿迁移路线模拟. 应用生态学报, 2014, 25(6): 1669-1673.
[47]江泽平. 麻栎-栓皮栎及小叶栎的生态地理学[D]. 北京: 中国林业科学研究院, 1991.
[48]李迎超, 厉月桥, 王利兵, 于海燕, 张兴锐, 刘鹏飞. 木本淀粉能源植物栓皮栎与麻栎的资源调查以及分布规律. 林业资源管理, 2013, 4(2): 94-101.
[49]吴征镒. 中国植被. 北京: 科学出版社, 1980: 261-262.
[50]郑万钧. 中国树木志(2卷). 北京: 中国林业出版社, 1985: 23-30.
[51]傅焕光, 于光明. 栓皮栎栽培与利用. 北京: 中国林业出版社, 1986: 7-10.
[52]周建云, 林军, 何景峰, 张文辉. 栓皮栎研究进展与未来展望. 西北林学院学报, 2010, 25(3): 43-49.
[53]王琫瑜, 孙湘君. 内蒙古察素齐泥炭剖面全新世古环境变迁的初步研究. 科学通报, 1997, 42(5): 514-518.
[54]李平日, 黄镇国, 张仲英, 宗永强. 广东东部晚更新世以来的海平面变化. 海洋学报, 1987, 9(2): 216-222.
[55]Rémy J Petit, Hu F S, Christopher W Dick. Forests of the past: a window to future changes. Science, 2008, 320(5882): 1450-1452.
[56]Désamoré A, Laenen B, Stech M, Papp B, Hedenäs L, Mateo R G, Vanderpoorten A. How do temperate bryophytes face the challenge of a changing environment? Lessons from the past and predictions for the future. Global Change Biology, 2012, 18(9): 2915-2924.
Impact of climate change on the potential geographical distribution pattern and dominant climatic factors ofQuercusvariabilis
GAO Wenqiang1, WANG Xiaofei1,JIANG Zeping1,2, LIU Jianfeng1,2,*
1ResearchInstituteofForestry,ChineseAcademyofForestry,Beijing100091,China2KeyLaboratoryofTreeBreedingandCultivation,StateForestryAdministration,Beijing100091,China
In order to predict the impact of climate change on the distribution ofQuercusvariabilis, one of the most widely distributed species in East Asia, the maximum entropy model (Maxent) was used to analyze the potential distribution under past (Mid Holocene, 6000 years ago), current (1950—2000) and future (2050) climate conditions. The jackknife method was used to reveal the contribution of the dominant climatic factors ofQ.variabilis. The results showed that the potential distribution ofQ.variabilisunder the current climate conditions centered on the south of East Asia, comprising up to 21.88% of the total studied area. The distribution range was mostly located in central and south of China, as well as south of the Korean peninsula, and central to south of Japan. Suitable habitats, comprising 5.69% of the studied area, were mainly centered on Qinling, Daba, Funiu, Luoxiao, Nanling, and Wuyi Mountains, the Yunnan-Guizhou Plateau, and Taiwan. The climate change scenarios showed that the areas of the suitable habitats have changed slightly, while significantly for the distribution range. With changes in global climate volatility, the distribution centers ofQ.variabilisare gradually concentrating in the regions of Qinling and Daba Mountains and Yunnan-Guizhou Plateau, and the areas of suitable habitats are gradually expanding. Minimum air temperature of the coldest month (Bio6), mean air temperature of the coldest quarter (Bio11), and annual precipitation (Bio12) were found to be dominant in influencing the geographic distribution ofQ.variabilis, with contribution rates of 48.6%, 21.4%, and 14.2%, respectively.
Quercusvariabilis; climate change; Maxent model; distribution pattern
国家自然科学基金项目(41371075);北京市科技计划课题(Z141100002314009)
2014-12-01;
2015-12-08
Corresponding author.E-mail: liujf@caf.ac.cn
10.5846/stxb201412012387
高文强,王小菲,江泽平,刘建锋.气候变化下栓皮栎潜在地理分布格局及其主导气候因子.生态学报,2016,36(14):4475-4484.
Gao W Q, Wang X F,Jiang Z P, Liu J F.Impact of climate change on the potential geographical distribution pattern and dominant climatic factors ofQuercusvariabilis.Acta Ecologica Sinica,2016,36(14):4475-4484.
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:34
中国中医药信息杂志(2023年6期)2023-06-13 02:05:36
落叶果树(2021年6期)2021-02-12 01:29:10
南方农业·下旬(2020年1期)2020-06-01 07:47:32
植物资源与环境学报(2020年2期)2020-04-20 07:18:02
现代园艺(2019年24期)2019-01-07 01:19:36
竹子学报(2018年2期)2018-10-25 02:02:12
现代农业科技(2009年21期)2018-09-03 00:00:00
农村百事通(2017年18期)2017-02-08 11:23:54
体育科技(2016年2期)2016-02-28 17:06:07
