
黄河下游平原非农植物多样性拆分研究
2016-09-21 02:13卢训令梁国付丁圣彦
生态学报 2016年14期
卢训令,汤 茜,梁国付,丁圣彦,*
1 教育部黄河中下游数字地理技术重点实验室,开封 475004 2 河南大学环境与规划学院,开封 475004 3 河南大学生态科学与技术研究所,开封 475004
黄河下游平原非农植物多样性拆分研究
卢训令1,2,3,汤茜1,2,梁国付1,2,丁圣彦1,2,3,*
1 教育部黄河中下游数字地理技术重点实验室,开封4750042 河南大学环境与规划学院,开封4750043 河南大学生态科学与技术研究所,开封475004
非农生物多样性的存在是农业景观生态系统健康持续发展的基础,对农业景观非农生境中植物群落物种多样性特征分析将有助于可持续农业景观构建措施的科学提出。在黄河下游平原典型农业景观中采用栅格分区的方式布设样点(共54个),采用典型样地法对各样点内的林地、树篱、田间道路和沟渠等主要非农生境的植物群落进行调查,采用生物多样性加性分配的方法探讨不同空间尺度上生物多样性的组成特征。结果显示:(1)各非农生境间植物群落物种多样性特征存在较大的差异。(2)偶见种从数量上构成了各非农生境中植物物种丰富度的主体,而常见种则行使着群落优势种和构建者的角色。(3)总体上,β多样性在各空间尺度中均对总物种丰富度具有重要贡献。(4)常见种和偶见种中物种组成格局存在显著差异:常见种的物种丰富度主要由α多样性贡献,而β多样性则贡献了偶见种的绝大部分。简言之,β多样性对区内植物多样性的保护和维持意义重大,农业景观中非农生境类别的出现对总物种丰富度的提高具有重要作用;各生境中较高的样点间β多样性(β样点)意味着在看似均质化的农业景观背景中依然具有较高的区域差异;景观组成和构型的变化将对农业景观中植物群落特征和物种多样性产生重要影响,且对偶见种的影响更甚。未来,应从景观和区域等更大尺度上,基于农业景观生态系统功能和服务的综合考虑及可持续农业景观的建立来探讨农业活动与生物多样性保护的权衡。
非农生境;β多样性;加性分配;常见种;偶见种;黄河下游平原
在全球人口激增,粮食需求日益增加的背景下,为维持粮食供给和人类福祉的提高,农业用地持续扩张。统计数据显示全球耕地面积已达15亿hm2,约占据了地球陆地面积的12%,可利用农业区中的自然草地已经有超过一半的面积被开垦为农田和永久性牧场,剩余的大部分地方也变为了季节性放牧草地[1],耕地的进一步扩展已很有限[2]。且这种态势在发展中国家表现的尤为严重[1]。随着农业用地的持续扩张、土地利用集约化的加剧、化肥与杀虫剂、除草剂等的过量施用,与之相伴的则是农业景观中非农生物多样性和绝大多数生态系统服务的持续下降[3-9]。而众多的研究认为生物多样性对生态系统功能的形成具有重要作用[10-12],生物多样性的存在对农业景观生态系统的持续发展意义重大[13-14]。同时生物多样性的保护也更多的依赖于农业景观,依赖于农业景观提供的生境、食源、栖息地和庇护所等[1,15-16]。β多样性是指沿环境梯度不同群落间物种组成相异性或沿环境梯度的物种更替和周转速率,它表述了物种组成在不同时空尺度上的变化,是生物多样性的重要组成部分,构成了α多样性与区域γ多样性的桥梁[17],成为当前生物多样性研究中的重要热点问题,越来越多的研究人员开始关注β多样性和生物多样性分配的研究,并取得了大量成果[18-21]。已有研究认为,在复杂农业景观中,受局地生境因子变化和人类干扰,景观要素内部样方尺度的α多样性的变化可能会远超景观要素间的多样性变化[22],而样方内α多样性的增加不一定与区域总物种丰富度的变化存在必然联系[23]。因此。因此借助β多样性来分析景观或更大尺度上生物多样性的变化会更有助于物种多样性变化与景观异质性特征、人类活动强度等因子间的关系[23],也更有助于理解区域群落物种多样性的形成机制。
黄河下游平原作为我国最重要的粮食主产区与核心区之一,区内耕作历史悠久,农业景观成为最主要的景观类型。人类活动对区内的景观异质性、生物多样性和生态系统服务产生了强烈的影响。研究发现区内农业景观不同非农生境中植物多样性存在较大的差异[24-25],但在农业景观中,植物物种多样性在空间尺度上有何特征?不同尺度上物种多样性组成对区域总物种多样性会有何贡献?这种空间格局在不同植物类别间会有何差异?目前尚不十分清楚,而这些又是探讨生物多样性保护措施的前提和基础。本研究拟在我国东部平原典型农业景观中,采用加性分配的方法,从不同空间尺度(样方、样点、生境、区域)对植物物种多样性进行拆分以探讨物种多样性组成特征及其形成机制,从而为区域生物多样性保护策略的提出奠定科学基础。
1 研究区概况
研究区位于河南省新乡市封丘县,属典型的黄河下游冲积平原,是我国重要的商品粮基地县之一,地理坐标为:34°53′—35°14′ N,114°14′—114°45′ E之间,总面积约1225.6 km2。暖温带大陆性季风气候,多年平均降水量约615mm,多年均温约13.9℃,地带性植被为落叶阔叶林,但目前自然植被已消失殆尽,代之以农田和人工林地。区内人类耕种活动悠久,形成了以农田为主的景观背景(图1)。

图1 研究区景观概况和野外样点布设示意图Fig.1 Landscape classification map and the schematic diagram of plots distribution
2 研究方法
2.1样地设置和数据的收集
在研究区卫星影像和土地利用现状图的基础上,采用方格分区的方式进行布点,采用法瑞学派的典型样地法对各样点中主要非农生境(人工林地、树篱、田间道路和沟渠等)中的植物群落进行调查,记录所有调查物种的名称、多优度、群集度、物候期和各样方的地理坐标等。野外调查在2011—2013年进行,其中2011年共设置26个样点,2012年以后将取样点加密到54个,每个生境类别5个样方,全区共计调查880个样方(其中人工林地255个,树篱260个,田间道路260个,沟渠105个),在春、夏、秋三季重复取样。
2.2数据分析方法
(1)物种丰富度
以样地中实际出现的物种数目S表示,是最简单和使用最为广泛与久远的物种多样性测度方法,传统计算物种丰富度的方法不考虑物种优势度的差异,即忽略了常见种和偶见种间的差异,将二者在群落中的贡献一致化。物种丰富度稀疏化法是依据调查样方中的物种分布状态来估计物种丰富度的一种统计方法,该方法考虑了偶见种对总体物种丰富度的影响。采用EstimateS 9.1.0对数据进行分析,绘制稀疏化曲线,并基于抽样统计的方法将各生境物种丰富度数据外推至100个样点,绘制物种-样点数量累积曲线图,估算各生境中的物种丰富度和多样性特征[26-27]。
(2)常见种与偶见种的划分
参照前人研究方法[28-29],将研究区内调查的物种划分为常见种(在所有样方(880个)中出现频率超过25%的物种)和偶见种(仅在不超过5%的调查样方中出现的物种)。
(3)物种丰富度的加性分配
对植物多样性数据进行加性分配分析时,在生境尺度上将数据定义为两个水平:样方尺度和样点尺度;全区的数据定义为3个水平:样方尺度(880个),样方组合尺度(176个)和生境类别尺度(4个),随机化计算次数为100;计算结果中的α样方为level 1(样方)的物种丰富度,β1为样点尺度上样方间物种多样性差异,α+β1为样点水平上的总物种丰富度(α样点或γ样点);β2为样点间物种多样性差异,α2+β2为区域中该生境类别的总物种丰富度(α生境或γ生境);β3是生境间β多样性,即γ区域=α样方+β1+β2+β3或γ区域=γ生境+β3,所有计算过程在软件PARTITION 3.0中进行[30]。将分解的α、β和γ多样性数据导入到Microsoft Office Excel 2007 中进行作图。
3 结果分析
3.1物种多样性总体特征
物种丰富度在林地和树篱生境中最高(实调数据分别为155和154种,EstimateS估计值分别为167.5和165.4),其次是沟渠生境,共调查记录到138种(估计值为156.2),田间道路生境中最少,实调数据为136种(估计值为148.3),方差分析显示林地和树篱生境中的物种丰富度要显著高于沟渠与田间道路生境(图2)。物种丰富度-样点数量累积曲线显示,随着调查样方的增加,物种丰富度迅速增加,在增加到10个样方的时候,沟渠生境中已涵盖了总物种丰富度的74.60%,最少的田间道路生境中也达到了66.70%,增加到20个样方时,沟渠生境中的比例已达到了87.52%,其它3类生境中也均接近80%。这表明,研究区内非农生境中植物群落以大量的常见种组成了群落的主体,具有较高的优势度,而偶见种则具有较高的空间差异性,并随着调查样方数量的增加而缓慢的加入进来。
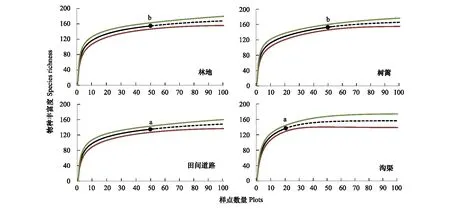
图2 不同生境中物种稀疏化和外推曲线图Fig.2 The rarefaction and extrapolation curves of plant species richness in different habitats图中黑色实心圆为实际调查物种丰富度值,实心曲线为稀疏化后的结果,虚线为外推结果曲线,上下两条彩色线之间为95%置信区间; 图中字母a、b表示方差分析的结果,P<0.05

图3 不同生境中Shannon多样性指数的稀疏化结果 Fig.3 The rarefaction curves of Shannon diversity index in different habitats图中字母a、b、c表示方差分析的结果,P<0.05
Shannon多样性指数显示,林地生境中物种多样性最高(3.70),其次是沟渠(3.59)和树篱生境(3.47),田间道路生境中最低(2.88),方差分析表明,林地、沟渠和树篱生境中Shannon多样性显著高于田间道路生境(图3)。通过稀疏化法对取样点Shannon多样性指数重取样绘制多样性指数-样点数量图发现,其变化态势与物种丰富度-样方数量累积曲线相似:随着样方数量的增加,物种多样性指数迅速升高,且升高速率要高于前者,多在10个左右的样方数量时即已达到总多样性的96%以上,表明在该区域内本研究中的样方调查数量能够满足对区内植物群落物种多样性特征的分析。
3.2物种丰富度加性分配
黄河下游平原农业景观非农生境中植物物种丰富度加性分配结果表明(图4):样方尺度上物种丰富度(α)对各生境总物种丰富度的贡献均较小,从高倒低依次是:林地(28.9种,18.63%)、树篱(26.8种,17.38%)、沟渠(20.4种,14.75%)和田间道路生境(20.0种,14.70%);从全区来看,α多样性的贡献度则仅占区域总物种丰富度的12.69%。样方间物种多样性差异(β1)对各生境类别物种丰富度的贡献也相对较低,均不超过该生境总物种丰富度的17%。其中林地和树篱生境中β1的贡献度要稍低于α多样性,田间道路和沟渠中β1的贡献度却高于α,说明这两类生境中各样点上不同样方间的差异要高于林地和树篱生境。从全区来看,β1的贡献也很低,与α的贡献基本持平。样点间物种多样性差异(β2)在各生境总物种丰富度中意义重大(均>60%),最高的田间道路和沟渠生境中贡献度均达69%以上;从全区来看,β2的贡献也最高,贡献了区域总物种丰富度的50.23%。从区域尺度看,生境间的物种多样性差异(β3)在总物种丰富度中也占有较大比重(48.3种,24.87%),基本等同于样点水平上的α多样性(α+β1=48.31种),再次表明研究区内植物物种组成在各生境间存在较大差异(图2,图3)。

图4 研究区不同生境物种丰富度加性分配图Fig.4 Additive partitioning of plant species diversity in different habitats*前的数据为物种丰富度值,()中数据为该水平物种丰富度占总丰富度的百分比(%)
3.3不同尺度β多样性对γ多样性的贡献
将区域内的总物种丰富度(γ区域)拆分为不同尺度,探讨β多样性对各级γ多样性的贡献性(表1)。在样点尺度上(γ样点),α多样性(α样方)与β多样性(β1)的贡献在各生境间差异不大,但林地和树篱生境中α样方>β1,而田间道路和沟渠生境则相反。在生境尺度上(γ生境),样点间物种丰富度差异(β2)的贡献率明显大于样点内的物种丰富度(γ样点或α样点)。从区域总物种丰富度(γ区域)来看,生境内物种丰富度(γ生境或α生境)的贡献达到了75.13%,生境间物种多样性差异(β3)依然贡献了总物种丰富度近四分之一。结合图4可以发现,β多样性对于区内生物多样性的保护和维持意义重大。样点间较高的β多样性(β2)意味着在看似更均质化的农业景观背景中依然具有较高的区域差异,景观异质性变化将会对生物多样性产生重要影响。

表1 不同生境中β多样性对各级γ多样性的贡献率
3.4常见种与偶见种比较分析
在黄河下游平原农业景观非农生境中根据物种出现频率划分结果表明(图5),常见种数量远低于偶见种(35∶105);从生境类别来看,35个常见种在4个生境中均有分布,但常见种在4个生境中的分布仍存在显著差异;偶见种在不同生境间差异显著,林地和树篱生境数量最多,分别达66种和65种,田间道路中最少,仅有47种。
物种丰富度代表了物种组成名录的变化,但无法表征各物种在群落中的地位,为此对各生境中不同功能群的累积重要值进行分析(图5)。结果表明:在个生境中,常见种对群落的贡献均远高于偶见种;偶见种在各生境中的贡献则与常见种相反,二者差值高达5—20倍以上。结果表明,田间道路生境中无论从物种组成还是从物种重要值来看,都是以常见种为主,偶见种的贡献显著低于其它生境;而沟渠的情况与之相反,该生境中的偶见种以生境特有种为主,故而数量不多的偶见种在沟渠生境中具有显著优于其它生境(累积重要值为0.21)的优势。

图5 不同生境中常见种与偶见种物种丰富度与累积重要值特征Fig.5 Species richness and accumulate importance value of common and rare species in different habitats图中字母表示各生境间物种丰富度方差分析的结果,小写字母表示偶见种,大写字母表示常见种,P<0.05
常见种与偶见种物种多样性加性分配结果表明(图6):样方水平的物种丰富度(α多样性)在两类物种中的贡献存在显著差异。常见种中,α在不同生境总物种丰富度中的贡献均较高(33%—53%左右),但各生境间有一定差异,从全区来看,α贡献度接近半数(47.17%);偶见种的情况与常见种大致相反,各生境中α的贡献度均很低(均<6%),从全区来看,α多样性的贡献度仅为1.86%。
样方间物种多样性差异(β1)在两类物种中的贡献亦存在较大差异。常见种中,β1在各生境中的贡献度差异不大(24.80% — 27.86%间),在全区中的贡献为27.77%;偶见种中,β1的贡献度亦很低,且在各生境间存在较大差异:沟渠中最高(6.48%),其次是林地(4.03%)和树篱生境(2.20%),田间道路中最低,约为0,全区的贡献也仅为2.73%。
样点间物种多样性差异(β2)在两类物种间差异巨大。常见种中,β2的贡献在各生境间差异较大,沟渠生境中高达41.94%,而林地和树篱生境中则仅为20%强,在全区中的贡献为15.94%。偶见种中,β2在各生境中均有极高的贡献(高达87.94%—97.21%),同时贡献了全区的约半数(50.17%)。
生境间物种多样性差异(β3)在常见种中的贡献约为9.11%,远低于偶见种中的比例(45.24%)。

图6 研究区常见种与偶见种物种丰富度加性分配结果Fig.6 Additive partitioning of common and rare species in different habitats*前的数据为物种丰富度值,()中数据为该水平丰富度占总物种丰富度的比例(%); **因偶见种在各生境中的α和β1数值太小,图上未显示数据标志
4 讨论
4.1物种多样性的拆分研究
生物多样性是群落生态学研究中最核心的内容,也是贯穿几乎所有生态学科的主要内容,关于生物多样性的研究数不胜数。但众多研究所关注的多是不同尺度上生物物种的编目多样性,即Whittake提出的生物多样性组成中的α多样性和γ多样性(二者的区别仅是尺度不同而已)。而对二者间桥梁——β多样性的研究则相对较弱,但β多样性的分析更有助于对生物群落结构、物种多样性动态和变化机制的理解,因而近年来愈来愈受到人们的关注[31-33],研究表明β多样性在区域景观生态系统中发挥着重要作用,在生物多样性保护中扮演着重要角色,是区域物种丰富度的重要组成部分。研究人员提出了β多样性的多种度量和计算方法[34]。加性分配方法,可以在多个不同尺度上将γ多样性拆分为β多样性和α多样性,且度量单位一致,其生态学意义更易于理解和解释,便于比较不同时空尺度和景观结构间的多样性差异,从而为众多研究人员所接受和运用[19, 29,34-41]。当前关于农业景观背景中生物多样性组成及其尺度变化也越来越受到重视,研究人员通过生物多样性拆分进行了大量的研究,对农业景观中景观异质性特征、耕作模式、管理方式等对生物多样性的影响效应和机制有了更深入理解[28, 31,42-47]。本研究通过物种多样性拆分也发现,在不同尺度上、不同物种类别中,植物群落物种丰富度组成存在显著的空间差异,而β多样性在不同尺度上均对γ多样性具有重要贡献。如Poggio等对阿根廷潘帕斯(Pampas)草原人工管理农业景观中农业植物多样性研究也发现,β多样性在各景观类型下均解释了γ多样性绝大部分的变化[48]。
4.2常见种与偶见种的划分研究
植物调查个体在调查样方中的频度是表征其在群落中地位(重要值)的重要组成部分,而常用的物种丰富度指数不考虑常见种与偶见种对群落多样性结构贡献的差异。依据物种出现频率划分为常见种与偶见种,则利于更清晰的认识群落物种丰富度组成及其空间格局特征。研究结果表明,偶见种对区域总物种丰富度组成具有极高的贡献,明显高于常见种,且两类物种的多样性组成特征亦存在显著差异:总体上,各生境不同尺度总物种丰富度组成中α多样性的贡献度多在30%—50%左右,且随着尺度增大而有降低;常见种中,α多样性的贡献则均高达60%以上,并随着尺度增大而上升;偶见种中,β多样性的贡献则极为重要,β2贡献了各生境中偶见种物种丰富度的绝大部分,表明区域内偶见种物种丰富度主要源自于大尺度上空间样点间群落组成间的差异。这与其他研究人员的结论较一致。如Klimek等在德国下萨克森州农业景观中的研究显示β多样性对全区所有物种和偶见种多样性的具有极高的贡献(分别为81%和96%),而对常见种的贡献仅为51%,且研究也表明在不同的空间尺度上β多样性的贡献存在一定的差异性[29];Gabriel等在欧洲农业景观中对植物群落中β多样性特征的研究结果表明,β多样性在中尺度和大尺度上对植物群落物种丰富度具有重要作用,且在常见种和偶见种中的作用有一定的差异,偶见种的物种丰富度主要源自于中、大尺度上群落组成的差异,而常见种的物种多样性则主要源自于小尺度和中尺度(样方和样点)[28]。
4.3不同生境中植物群落物种多样性组成差异
林地、树篱、田间道路和沟渠是黄河下游平原农业景观中最主要的非农景观要素和生境类别。各生境类别的形状、面积、环境要素特征、空间格局和人类活动干扰强度均存在差异。单从物种丰富度指数来看,林地和树篱,沟渠与田间道路间的差异很小,但其群落结构特征却存在显著差异[25]。不同生境类别中物种多样性组成尺度特征分析有助于对区内物种多样性特征的理解及未来生物多样性保护措施的提出与制定。同时,非农生境的存在是区域农业景观生态系统健康持续发展的重要保证,研究表明各非农自然、半自然景观要素对农业景观中不同生物类群和降低农业活动对环境的负效应均具有重要作用[49-54],而关于各生境中植物群落结构和物种多样性格局的探讨则稍显不足[55-57],而各非农生境及其上的植物群落扮演着食源地、栖息地、庇护所、缓冲带、水土保持者等不可或缺的重要角色[1,16]。
5 结论和建议
在黄河下游平原典型农业景观中,各非农生境植物群落中物种多样性和组成格局存在显著差异。各非农生境中物种组成均以众多的偶见种为主体,而常见种则以优势种的身份塑造了群落的外貌和结构。空间差异引起的β多样性变化是偶见种和总体物种丰富度的主体,而常见种则主要由α多样性来解释,其空间差异性很低。
众多的研究证实,农业景观中非农生境的存在对区域农业景观生态系统结构、功能和服务具有重要作用。而黄河下游平原农业景观属于人类耕作活动极其悠久和强烈、各种人工辅助能大量使用、自然生境几乎消失殆尽、仅余的非农生境多呈斑块状和条带状镶嵌分布,面积很小。而已有研究表明农业景观中自然、半自然生境的比例对生物多样性保护和保护措施的效应存在重要影响[58]。区域内农业景观各非农生境中尽管留存有较高的物种丰富度,但以偶见种为主,而常见种则在绝大多数的调查样方中形成优势种群,并具有较高的空间相似性。结合区域内的相关研究,认为,区域内农业景观中非农生境中植物群落物种组成以广布种为主,在较大尺度上,偶见种的随机性分布成为物种丰富度的重要组成。因此,在区域内未来生物多样性保护策略中,建议不应以物种丰富度高低为目标,而更应关注非农植物群落存在在整个景观生态系统功能中的作用,如:水土保持、碳氮循环、污染物截留降解、调节区域小气候以及其他生物类群的食源地、栖息地、掩蔽物等。并进一步从提高生态系统功能和生态系统服务维持的角度探讨农业活动与生物多样性保护的权衡。
致谢:感谢中国科学院封丘农业生态实验站对野外实验提供的支持。
[1]Green R E, Cornell S J, Scharlemann J P W, Balmford A. Farming and the fate of wild nature. Science, 2005, 307(5709): 550-555.
[2]FAO STAT. FAO Statistical Yearbook 2013 World Food and Agriculture. United Nations, Rome, 2013.
[3]Sala O E, Chapin F S III, Armesto J J, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke L F, Jackson R B, Kinzig A, Leemans R, Lodge D M, Mooney H A, Oesterheld M, Poff N L, Sykes M T, Walker B H, Walker M, Wall D H. Global biodiversity scenarios for the year 2100. Science, 2000, 287(5459): 1770-1774.
[4]Robinson R A, Sutherland W J. Post-war changes in arable farming and biodiversity in Great Britain. Journal of Applied Ecology, 2002, 39(1): 157-176.
[5]Foley J A, DeFries R, Asner G P, Barford C, Bonan G, Carpenter S R, Chapin F S, Coe M T, Daily G C, Gibbs H K, Helkowski J H, Holloway T, Howard E A, Kucharik J, Monfreda C, Patz J A, Prentice I C, Ramankutty N, Snyder P K. Global consequences of land use. Science, 2005, 309(5734): 570-574
[6]Billeter R, Liira J, Bailey D, Bailey D, Bugter R, Arens P, Augenstein I, Aviron S, Baudry J, Bukacek R, Burel F, Cerny M, De Blust G, De Cock R, Diekötter T, Dietz H, Dirksen J, Dormann C, Durka W, Frenzel M, Hamersky R, Hendrickx F, Herzog F, Klotz S, koolstra B, Lausch A, Le Coeur D, Maelfait J P, Opdam P, Roubalova M, Schermann, A, Schermann N, Schmidt T, Schweiger O, Smulders M J M, Speelmans M, Simova P, Verboom J, Van Wingerden W K R E, Zobel M, Edwards P J. Indicators for biodiversity in agricultural landscapes: a pan-European study. Journal of Applied Ecology, 2008, 45(1): 141-150.
[7]Carpenter S R, Mooney H A, Agard J A, Capistranod D, DeFriese R S, Díazf S, Dietzg T, Duraiappahh A K, Oteng-Yeboahi A, Pereiraj H M, Rerringsk C, Reidl W V, Sarukhanm J, Scholesn R J, Whyteo A. Science for managing ecosystem services: Beyond the millennium ecosystem assessment. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(5): 1305-1312.
[8]Butchart S H M, Walpole M, Collen B, Van Strien A, Scharlemann J P W, Almond R E A, Baillie J E M, Bomhard B, Brown C, Bruno J, Carpenter K E, Carr G M, Chanson J, Chenery A M, Csirke J, Davidson N C, Dentener F, Foster M, Galli A, Galloway J N, Genovesi P, Gregory R D, Hockings M, Kapos V, Lamarque J F, Leverington F, Loh J, McGeoch M A, McRae L, Minasyan A, Morcillo M H, Oldfield T E E, Pauly D, Quader S, Revenga C, Sauer J R, Skolnik B, Spear D, Stanwell-Smith D, Stuart S N, Symes A, Tierney M, Tyrrell, T D, Vié J C, Watson R. Global biodiversity: indicators of recent declines. Science, 2010, 328(5982): 1164-1168.
[9]Meehan T D, Werling B P, Landis D A, Gratton C. Agricultural landscape simplification and insecticide use in the Midwestern United States. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(28): 11500-11505.
[10]Hooper D U, Chapin F S, Ewel J J, Hector A, Inchausti P, Lavorel S, Lawton J H, Lodge D M, Loreau M, Naeem S, Schmid B, Setälä H, Symastad A J, Vandermeer J, Wardle D A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs, 2005, 75(1): 3-35.
[11]Cardinale B J, Duffy E, Gonzalez A, Hooper D U, Perrings C, Venail P, Narwani A, Mace G M, Tilman D, Wardle D A, Kinzig A P, Daily G C, Loreau M, Grace J B, Larigauderie A, Srivastava D S, Naeem S. Biodiversity loss and its impact on humanity. Nature, 2012, 486(7401): 59-67.
[12]Midgley G F. Biodiversity and ecosystem function. Science, 2012, 335(6065): 174-175.
[13]Altieri M A. The ecological role of biodiversity in agroecosystems. Agriculture, Ecosystems & Environment, 1999, 74(1/3): 19-31.
[14]Marshall E J P, Brown V K, Boatman N D, Lutman P J W, Squire G R, Ward L K. The role of weeds in supporting biological diversity within crop fields. Weed Research, 2003, 43(2): 77-89.
[15]Rodrigues A S L, Andelman S J, Bakarr M I, Boitani L, Brooks T M, Cowling R M, Fishpool L D C, da Fonseca G A B, Gaston K J, Hoffmann M, Long J S, Marquet P A, Pilgrim J D, Pressey R L, Schipper J, Sechrest W, Stuart S N, Underhill L G, Waller R W, Watts M E J, Yan X. Effectiveness of the global protected area network in representing species diversity. Nature, 2004, 428(6983): 640-643.
[16]Fahrig L, Baudry J, Brotons L, Burel F G, Crist T O, Fuller R J, Sirami C, Siriwardena G M, Martin J L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecology Letters, 2011, 14(2): 101-112.
[17]Whittaker R H. Vegetation of the Siskiyou mountains, Oregon and California. Ecological Monographs, 1960, 30(3): 279-338.
[18]Crist T O, Veech J A, Gering J C, Summerville K S. Partitioning species diversity across landscapes and regions: a hierarchical analysis of α, β, and γ diversity. The American Naturalist, 2003, 162(6): 734-743.
[19]Crist T O, Veech J A. Additive partitioning of rarefaction curves and species-area relationships: unifying α-, β-and γ-diversity with sample size and habitat area. Ecology Letters, 2006, 9(8): 923-932.
[20]Legendre P, De Cáceres M. Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecology Letters, 2013, 16(8): 951-963.
[21]López-Martínez J O, Hernández-Stefanoni J L, Dupuy J M, Meave J A. Partitioning the variation of woody plant β-diversity in a landscape of secondary tropical dry forests across spatial scales. Journal of Vegetation Science, 2013, 24(1): 33-45.
[22]Tscharntke T, Tylianakis J M, Rand T A, Didham R K, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist T O, Dormann C F, Ewers R M, Fründ J, Holt R D, Holzschuh A, Kleijn D, Kremen C, Landis D A, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, van der Putten W H, Westphal C. Landscape moderation of biodiversity pattern and processes-eight hypotheses. Biological Reviews, 2012, 87(3): 661-685.
[23]Egan J F, Mortensen D A. A comparison of land-sharing and land-sparing strategies for plant richness conservation in agricultural landscapes. Ecological Applications, 2012, 22(2): 459-471.
[24]卢训令, 梁国付, 汤茜, 丁圣彦, 李乾玺, 张晓青. 黄河下游平原农业景观中非农生境植物多样性. 生态学报, 2014, 34(4): 789-797.
[25]卢训令, 汤茜, 梁国付, 丁圣彦. 黄河下游平原不同非农生境中植物多样性. 生态学报, 2015, 35(5): 1527-1536.
[26]Colwell R K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. 2013. http://viceroy.eeb.uconn.edu/estimates/.
[27]Colwell R K, Chao A, Gotelli N J, Lin S Y, Mao C X, Chazdon R L, Longino J T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. Journal of Plant Ecology, 2012, 5(1): 3-21.
[28]Gabriel D, Roschewitz I, Tscharntke T, Thies C. Beta diversity at different spatial scales: plant communities in organic and conventional agriculture. Ecological Applications, 2006, 16(5): 2011-2021.
[29]Klimek S, Marini L, Hofmann M, Isslstein J. Additive partitioning of plant diversity with respect to grassland management regime, fertilisation and abiotic factors. Basic and Applied Ecology, 2008, 9(6): 626-634.
[30]Veech J A, Crist T O. PARTITION: software for hierarchical partitioning of species diversity, version 3.0 2009. http://www.users.miamioh.edu/cristto/partition.htm.
[31]Wagner H H, Wildi O, Ewald K C. Additive partitioning of plant species diversity in an agricultural mosaic landscape. Landscape Ecology, 2000, 15(3): 219-227.
[32]Anderson M J, Crist T O, Chase J M, Vellend M, Inouye B D, Freestone A L, Sanders N J, Cornell H V, Comita L S, Davies K F, Harrison S P, Kraft N J B, Stegen J C, Swenson N G. Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecology Letters, 2011, 14(1): 19-28.
[33]Carvalho J C, Cardoso P. Drivers of beta diversity in Macaronesian spiders in relation to dispersal ability. Journal of Biogeography, 2014, 41(10): 1859-1870.
[34]陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323-335.
[35]Concepción E D, Fernández-González F, Díaz M. Plant diversity partitioning in Mediterranean croplands: effects of farming intensity, field edge, and landscape context. Ecological Applications, 2012, 22(3): 972-981.
[36]Chávez V, Macdonald S E. Partitioning vascular understory diversity in mixedwood boreal forests: the importance of mixed canopies for diversity conservation. Forest Ecology and Management, 2012, 271: 19-26.
[37]Golodets C, Kigel J, Sternberg M. Plant diversity partitioning in grazed Mediterranean grassland at multiple spatial and temporal scales. Journal of Applied Ecology, 2011, 48(5): 1260-1268.
[38]Chiarucci A, Bacaro G, Ramón Arévalo J, Delgado J D, Fernández-Palacios J M. Additive partitioning as a tool for investigating the flora diversity in oceanic archipelagos. Perspectives in Plant Ecology, Evolution and Systematics, 2010, 12(2): 83-91.
[39]Jost L. Partitioning diversity into independent alpha and beta components. Ecology, 2007, 88(10): 2427-2439.
[40]Chao A, Chiu C H, Hsieh T C. Proposing a resolution to debates on diversity partitioning. Ecology, 2012, 93(9): 2037-2051.
[41]Zhang Q, Hou X Y, Li F Y, Niu J M, Zhou Y L, Ding Y, Zhao L Q, Li X, Ma W J, Kang S. Alpha, Beta and Gamma diversity differ in response to precipitation in the Inner Mongolia grassland. PLoS One, 2014, 9(3): e93518.
[42]Hendrickx F, Maelfait J P, Van Wingerden W, Schweiger O, Speelmans M, Aviron S, Laugenstein I, Billeter R, Bailey D, Bukacek R, Burel F, diekötter, T, Dirksen J, Herzog F, Liira J, Roubalova M, Vandomme V, Bugter R. How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. Journal of Applied Ecology, 2007, 44(2): 340-351.
[43]Diekötter T, Billeter R, Crist T O. Effects of landscape connectivity on the spatial distribution of insect diversity in agricultural mosaic landscapes. Basic and Applied Ecology, 2008, 9(3): 298-307.
[44]Flohre A, Fischer C, Aavik T, Bengtsson J, Berendse F, Bommarco R, Ceryngier P, Clement L W, Dennis C, Eggers S, Emmerson M, Geiger F, Guerrero I, Hawro V, Inchausti P, Liira J, Morales M B, Oate J J, Pärt T, Weisser W W, Winqvist C, Thies C, Tscharntke T. Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids, and birds. Ecological Applications, 2011, 21(5): 1772-1781.
[45]Ernoult A, Alard D. Species richness of hedgerow habitats in changing agricultural landscapes: are α andγ diversity shaped by the same factors?. Landscape Ecology, 2011, 26(5): 683-696.
[46]Schultz N L, Reid N, Lodge G, Hunter J T. Broad-scale patterns in plant diversity vary between land uses in a variegated temperate Australian agricultural landscape. Austral Ecology, 2014, 39(7): 855-863.
[47]Loos J, Turtureanu P D, von Wehrden H, Hanspach J, Dorresteijn I, Frink J P, Fischer J. Plant diversity in a changing agricultural landscape mosaic in Southern Transylvania (Romania). Agriculture, Ecosystems & Environment, 2015, 199: 350-357.
[48]Poggio S L, Chaneton E J, Ghersa C M. The arable plant diversity of intensively managed farmland: effects of field position and crop type at local and landscape scales. Agriculture, Ecosystems & Environment, 2013, 166: 55-64.
[49]Stockan J A, Baird J, Langan S J, Young M R, Iason G R. Effects of riparian buffer strips on ground beetles (Coleoptera, Carabidae) within an agricultural landscape. Insect Conservation and Diversity, 2014, 7(2): 172-184.
[50]Morandin L A, Long R F, Krenmen C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agriculture, Ecosystems & Environment, 2014, 189: 164-170.
[51]Lindborg R, Plue J, Andersson K, Cousins S A O. Function of small habitat elements for enhancing plant diversity in different agricultural landscapes. Biological Conservation, 2014, 169: 206-213.
[52]Fahrig L, Girard J, Duro D, Pasher J, Smith A, Javorek S, King D, Lindsay K F, Mitchell S, Tischendorf L. Farmlands with smaller crop fields have higher within-field biodiversity. Agriculture, Ecosystems & Environment, 2015, 200: 219-234.
[53]Power A G. Ecosystem services and agriculture: tradeoffs and synergies. Philosophical Transactions of the Royal Society B: Biological Sciences, 2010, 365(1554): 2959-2971.
[54]赵紫华, 欧阳芳, 贺达汉. 农业景观中不同生境界面麦蚜天敌的边缘效应与溢出效应. 中国科学: 生命科学, 2012, 42(10): 825-840.
[55]Gabriel D, Sait S M, Hodgson J A, Schmutz U, Kunin W E, Benton T G. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecology Letters, 2010, 13(7): 858-869.
[56]Jonason D, Andersson G K S, Öckinger E, Rundlöf M, Smith H G, Bengtsson J. Assessing the effect of the time since transition to organic farming on plants and butterflies. Journal of Applied Ecology, 2011, 48(3): 543-550.
[57]马守臣, 原东方, 杨慎骄, 王和洲. 豫北低山丘陵区农田边界系统植物多样性的研究. 中国生态农业学报, 2010, 18(4): 815-819.
[58]Tscharntke T, Klein A M, Kruess A, Steffan-Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecology Letters, 2005, 8(8): 857-874.
Additive partitioning of non-agricultural plant diversity in the lower reaches of the Yellow River plain
LU Xunling1, 2, 3, TANG Qian1, 2, LIANG Guofu1, 2, DING Shengyan1, 2, 3,*
1KeyLaboratoryofGeospatialTechnologyfortheMiddleandLowerYellowRiverRegions,MinistryofEducation,Kaifeng475004,China2CollegeofEnvironmentandPlanning,HenanUniversity,Kaifeng475004,China3InstituteofEcologicalScienceandTechnology,HenanUniversity,Kaifeng475004,China
Biodiversity plays a crucial role in producing and maintaining ecosystem functions and services, so biodiversity conservation and preservation are prerequisites for the sustainability of regional landscape ecosystems. The mechanisms determining spatial patterns of plant communities in non-agricultural habitats are the basis for the construction of sustainable agro-landscapes. To understand the composition of plant communities in non-agricultural habitats, 54 plots were established in Fengqiu County, Henan Province. Plant communities in non-agricultural habitats (planted forests, treelines, field roads, and ditches) were investigated using the Braun-Blanquet method, and the spatial pattern of plant species diversity was analyzed by additive partitioning. We found that species richness and diversity were significantly higher in planted forests and treelines than in field roads and ditches, and field roads had the lowest plant diversity. A comparative analysis of common and rare species showed that rare species were the main component of species richness in non-agricultural habitats, whereas common species were dominant in agricultural habitats. Additive partitioning of species richness showed thatβdiversity made an important contribution to total regional species richness across multiple spatial scales. At the quadrat scale,αdiversity contributed about 50% of the total species richness of common species, and the contribution ofα+β1was more than 75% of the total common species richness. For rare species, the contribution ofαwas less than 6% of total species richness in each habitat. In contrast,β2contributed up to 97% of total rare species richness, especially in field roads. At the regional scale, the contribution ofαwas only 1.86%, whileβ2+β3contributed more than 95% of total rare species richness. According to the results, the emergence of non-agricultural habitat could play an important role in improving total species richness in agro-landscapes. In addition, changes in landscape composition and configuration could have a profound impact on plant diversity, and the effects would be stronger for rare species. In future, more attention should be paid to the tradeoffs between agricultural production and sustainability of ecosystem services on landscape and regional scales, and improvement of the ecological processes and ecosystem services controlled by biodiversity should be emphasized.
non-agricultural habitats;βdiversity; additive partitioning; common species; rare species; lower reaches of the Yellow River plain
国家自然科学基金项目(41371195, 41071118)
2014-12-24; 网络出版日期:2015-10-30
Corresponding author.E-mail: syding@henu.edu.cn
10.5846/stxb201412242560
卢训令,汤茜,梁国付,丁圣彦.黄河下游平原非农植物多样性拆分研究.生态学报,2016,36(14):4395-4405.
Lu X L, Tang Q, Liang G F, Ding S Y.Additive partitioning of non-agricultural plant diversity in the lower reaches of the Yellow River plain.Acta Ecologica Sinica,2016,36(14):4395-4405.
猜你喜欢
湖北植保(2022年4期)2022-08-23
林业勘查设计(2022年1期)2022-02-15
世界博览(2021年23期)2021-12-16
中国果业信息(2021年1期)2021-12-01
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
土壤(2021年1期)2021-03-23
优雅(2020年8期)2020-09-16
林业调查规划(2020年3期)2020-06-03
土壤学报(2017年5期)2017-11-01
