
不同水深条件下刺参养殖池塘大型底栖动物群落结构季节性变化特征
2016-09-21 02:34高勤峰董双林宁鲁光
生态学报 2016年14期
温 彬, 高勤峰,*, 董双林, 宁鲁光
1 中国海洋大学海水养殖教育部重点实验室,青岛 266003 2 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室,青岛 266235 3 东营海跃水产养殖有限公司,东营 257500
不同水深条件下刺参养殖池塘大型底栖动物群落结构季节性变化特征
温彬1,2, 高勤峰1,2,*, 董双林1,2, 宁鲁光3
1 中国海洋大学海水养殖教育部重点实验室,青岛2660032 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室,青岛2662353 东营海跃水产养殖有限公司,东营257500
于2012年7月至2013年4月调查了荣成靖海湾3个不同水深的刺参(Apostichopusjaponicus)养殖池塘内大型底栖动物的构成,以了解不同水深对刺参养殖池塘环境条件的影响以及由此引起的大型底栖生物群落结构的改变。结果表明:3个不同水深梯度池塘(1#浅水位、2#正常水位和3#高水位)底部光照强度、叶绿素a(Chla)和总有机物(TOM)含量存在显著差异,各池塘水温差异不显著。光强、Chla和TOM含量在夏季、冬季和春季均表现为1#池塘显著高于3#池塘;秋季各池塘间光强和TOM含量差异不显著,Chla含量则表现为3#池塘显著高于1#池塘。各季节3个池塘间大型底栖动物在种类组成、丰度、生物量和多样性指数上均存在显著性差异。大型底栖动物丰度和生物量夏季均表现为1#池塘显著高于3#池塘,秋季和冬季则相反;春季1#池塘丰度显著高于3#池塘,生物量则差异不显著。这些差异主要与其各自优势种及其优势度指数大小有关。大型底栖动物多样性指数夏季和秋季均表现为1#池塘高于3#池塘,春季则相反,冬季各池塘间多样性指数差异不显著。单因子相似性分析(ANOSIM)表明,各季节3个池塘间大型底栖动物群落结构均存在显著差异,表明水深梯度对刺参养殖池塘大型底栖动物群落结构造成显著性影响。相似性百分比分析(SIMPER)显示,各季节对3个池塘间大型底栖动物群落差异起主要作用的物种为各个池塘的优势种。典范对应分析(CCA)表明,水深、Chla和TOM含量为影响大型底栖动物群落的主要环境因子。
水深;刺参;大型底栖动物;生物多样性
大型底栖动物在池塘生态系统食物网中发挥着重要作用,不仅是池塘生态系统中物质循环、能量流动过程的积极消费者和转移者,而且还可通过摄食、筑穴和建管等扰动活动间接或直接地影响池塘底质环境[1-2]。因此,大型底栖动物一方面可直接为池塘养殖刺参(Apostichopusjaponicus)提供食物来源[3],另一方面,大型底栖动物群落结构的稳定性对维持养殖池塘生态系统的平衡具有重要的作用。目前关于大型底栖动物和养殖动物关系的研究多集中于养殖活动对大型底栖动物群落的影响[4-10],例如,养殖活动产生的代谢废物、残饵以及营养物质再悬浮等造成水体污染,进而对大型底栖动物群落结构造成影响。而通过改变池塘水文条件来影响养殖池塘生物环境进而间接对养殖动物产生影响的报道较少。已有研究表明水深能够对大型底栖动物群落造成显著影响[11-17]。一方面,调节水深可以改变池塘底部温度和光照强度,从而影响底泥本身内源性初级生产力;同时,温度和光强的改变影响刺参的生长、发育和繁殖,水温升高是刺参夏眠的主要诱发因子[18],而水温过低时,刺参会出现活动停止、生长停止、代谢下降等现象[19]。刺参具避光行为,在光强较弱时活动频繁,而在光强较高时,刺参会栖息在人工礁阴影区或附着在参礁上[20]。另一方面,水深的改变影响水体-底泥界面的营养盐通量,以及由水体沉积到池塘底部浮游植物、有机碎屑等颗粒有机物,从而影响大型底栖动物群落结构[21]。因此,在不同季节不同水深条件下根据刺参养殖池塘非生物环境和生物环境变化特征确定合理的水深,对刺参的养殖产量会产生直接的影响。
本实验的目的为研究不同水深对靖海湾刺参养殖池塘大型底栖动物的物种组成、优势种以及群落结构的影响,旨在提供不同水深条件下刺参养殖池塘底部生物环境的季节性变化规律,为不同季节确定刺参养殖池塘合理的水深,发展高效健康的刺参养殖技术提供科学依据。
1 材料和方法
1.1实验地点
本实验于2012年7月至2013年4月在山东荣成好当家海洋发展股份有限公司靖海湾(36°86′N, 122°13′E)刺参养殖三场进行。选取3口条件类似的池塘,面积约为2 hm2(200 m×100 m×2 m)。池塘采用自然纳潮换水,实验开始后,通过控制进水闸门的纳水时间,调节池塘的水深,即3口池塘分别按照池塘最低水位(1.5 m)、正常水位(2.3 m)和最高水位(3.1 m)纳水,标记为1#,2#和3#池塘。高潮期每次换水量约为池塘水量的40%—60%。刺参投放密度为5 个/m2,刺参个体平均体重约为20 g。池塘不投饵。
在实验开始前一年对池塘底部进行人工修整且铺有聚丙烯薄膜,以保证池塘底质条件一致,池塘底泥主要为水层沉降下来的泥沙。每个池塘选取3个站位作为取样点,并用浮漂做标记,以确保每次取样位点相同。
1.2样品采集及处理
2012年7月(夏季)、2012年10月(秋季)、2013年1月(冬季)、2013年4月(春季)于3个刺参养殖池塘进行采样,利用面积为0.05 m2的抓斗式采泥器在每站位采集3个重复样,然后合并为1个样品,使用0.5 mm孔径的网筛分选底栖动物,并用75%酒精现场固定[22-23]。室内进行种类鉴定、个体计数及称重。样品的处理、保存、计数、称重均按照(海洋调查规范)(GB/T12763.6—2007)进行。
1.3环境因子的测定
在采样现场测定池塘水深、底部水温、光照强度;取表层5 cm泥样一部分于50 ℃低温烘干研磨,经丙酮萃取后于665 nm和750 nm波长下测定叶绿素a含量(Chla);另一部分在马弗炉550 ℃下灼烧6 h测定总有机物含量(TOM)。
1.4数据处理与分析
采用以下指数分析各月份各池塘大型底栖动物群落的多样性及优势种:
Shannon-Wiener多样性指数:
H′ = -∑Pilog2Pi
Margalef物种丰富度指数:
d= (S-1)1og2N
Pielou物种均匀度指数:
J=H′/1og2S
优势度指数:
Y=Pi×fi
式中,N为各样点采集的大型底栖动物总个数;S为种类总数;Pi为第i种的个数与总个数的比值;fi为物种i出现的频率;当物种优势度Y≥0.02时,该种为优势种。
大型底栖动物丰度经平方根转换后,计算Bray-Curtis相似性系数构建相似性矩阵进行等级聚类分析(Cluster)。运用单因子相似性分析(ANOSIM)进行3个池塘间群落结构差异显著性检验。相似性百分比分析(SIMPER)造成各池塘间群落结构差异的分歧种。运用典范对应分析(CCA)进行大型底栖动物与环境因子之间关系。分析数据分析由SPSS 20.0,PRIMER 6.0 和CANOCA 4.5软件完成。
2 结果与分析
2.1环境参数
刺参养殖池塘环境参数见表1。4个采样月3个池塘水深、光强、叶绿素a(Chla)和总有机物(TOM)含量均存在显著差异。水深各季节均表现3#池塘水深显著大于1#和#2#池塘,2#池塘均显著大于1#池塘。光强和TOM含量在夏、冬和春季均表现为1#池塘显著大于3#池塘,秋季各池塘间差异不显著。Chla含量在夏、冬和春季均表现为1#池塘显著大于3#池塘,秋季则表现为3#池塘显著大于1#池塘;3个池塘水温在各季节差异均不显著。
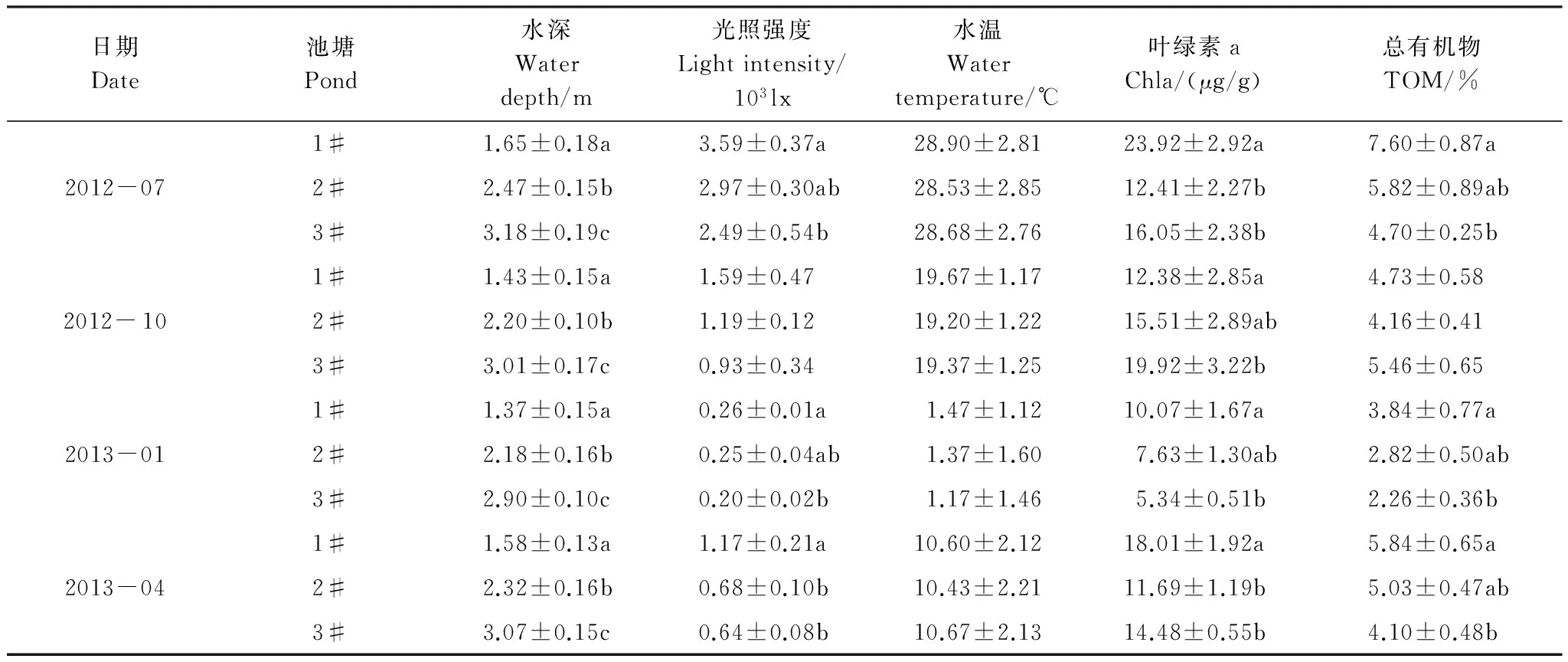
表1 刺参养殖池塘环境参数
数据采用平均值±标准差(n=3),各季节不同字母代表数据间有显著性差异(P<0.05)
2.2物种组成和优势种
4个采样月共鉴定出大型底栖动物41种,其中多毛纲环节动物21种,占51.2%,软体动物16种,占39.0%,甲壳动物4种,占9.8%。多毛类和软体动物是该区域刺参养殖池塘的主要类群。各季节3个池塘间大型底栖动物优势种差异较大,主要表现在优势种数多少以及优势度指数大小(表2)。
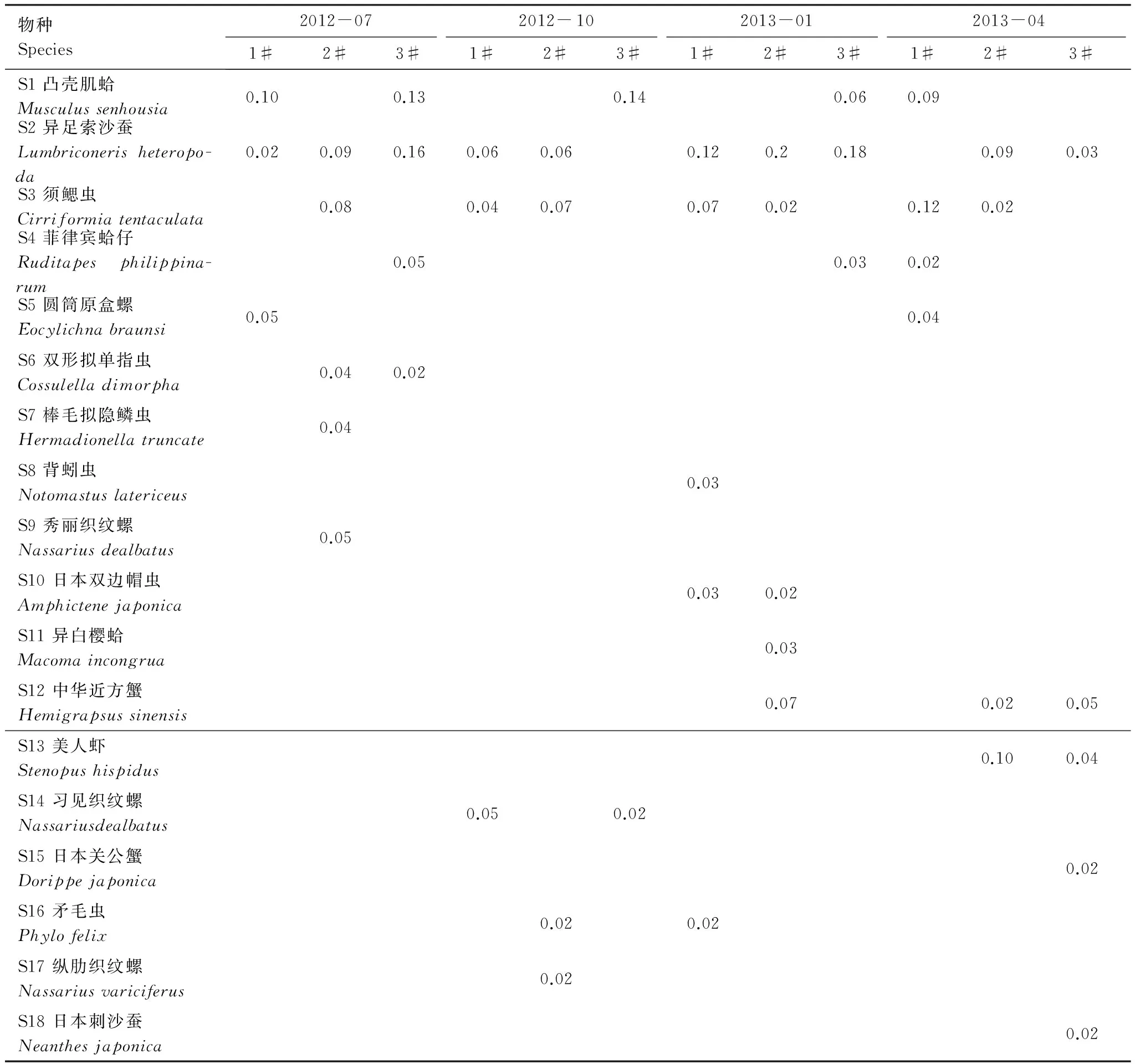
表2 刺参养殖池塘大型底栖动物优势种及其优势度指数
2.3丰度和生物量
4个采样月3个池塘大型底栖动物丰度和生物量如图1所示。夏季丰度和生物量均表现为1#池塘显著大于3#池塘,2#和3#池塘差异不显著;秋季和冬季则表现为3#池塘显著大于1#和2#池塘,1#和2#池塘差异不显著;春季丰度表现为1#池塘显著大于3#池塘,2#和3#池塘差异不显著,生物量则表现为1#池塘显著大于2#池塘,1#和3#池塘差异不显著。
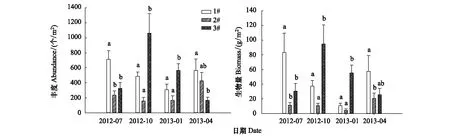
图1 刺参养殖池塘大型底栖动物丰度和生物量Fig.1 Macrobenthic abundance and biomass of each pond for each season数据采用平均值±标准差(n=3),各季节不同字母代表数据间有显著性差异(P<0.05)
2.4物种多样性
4个采样月3个池塘大型底栖动物的Shannon-Wiener多样性指数(H′)、Margalef物种丰富度指数(d)和Pielou物种均匀度指数(J)见表3。夏季1#池塘d显著大于3#池塘;秋季1#池塘H′和d显著大于3#池塘;冬季各池塘间H′、d和J差异均不显著;春季3#池塘d和J显著大于1#池塘。
2.5群落结构分析
大型底栖动物聚类分析结果如图2所示。4个采样月3个池塘大型底栖动物群落结构总体相似性为26%。夏季9个样点间的相似性为43%,ANOSIM 分析表明3个池塘间群落组成差异极显著(r=0.85,P<0.01)。秋季9个样点间的相似性较夏季降低,总体相似性为37%,ANOSIM 分析表明3个池塘间群落组成差异极显著(r=0.77,P<0.01)。冬季9个样点间的相似性较秋季有所增大,总体相似性43%,ANOSIM 分析表明3个池塘间群落组成差异极显著(r=0.88,P<0.01)。春季9个样点间的相似性继续增大,总体相似性为48%,ANOSIM 分析表明3个池塘间群落组成差异极显著(r=0.70,P<0.01)。SIMPER分析显示各季节造成3个池塘间大型底栖动物群落差异的主要物种为各个池塘的优势种(表4)。
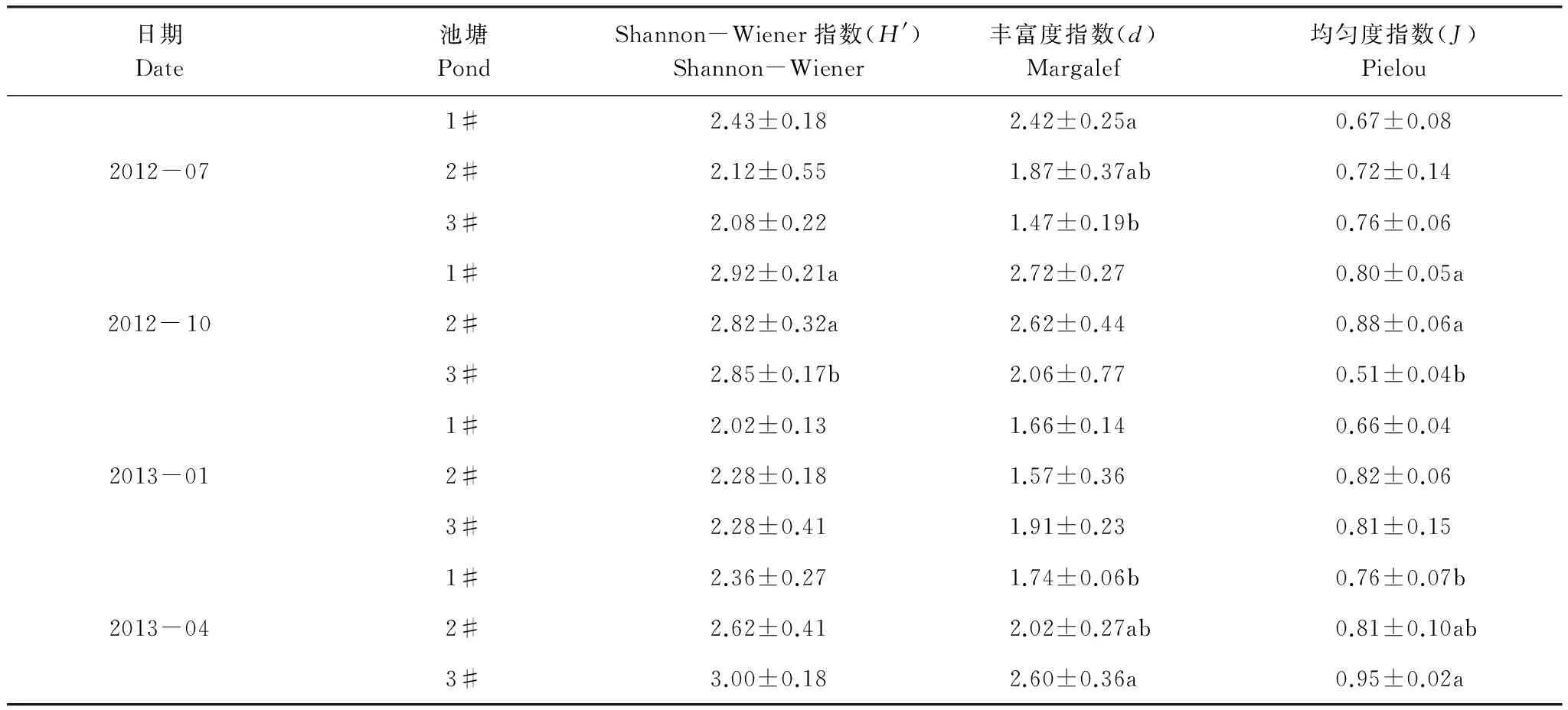
表3 刺参养殖池塘大型底栖动物多样性指数
数据采用平均值±标准差(n=3),各季节不同字母代表数据间有显著性差异(P<0.05)

图2 刺参养殖池塘不同样点大型底栖动物聚类分析Fig.2 Cluster analysis for each sample of each season
2.6大型底栖动物群落与环境因子的关系
大型底栖动物群落优势种与环境因子的CCA分析结果如图3。夏季排序图前两轴的特征值分别为0.437和0.255,种类与环境因子排序轴相关系数高达0.924和0.953,且物种-环境关系的累积百分率为77.1,说明排序轴与环境因子间线性结合的程度较好的反映了物种与环境间的关系。第一轴与Chla含量负相关(r=-0.76);第二轴与水深正相关(r=0.80),与TOM含量负相关(r=-0.47)。秋季排序图前两轴的特征值分别为0.558和0.095,种类与环境因子排序轴的相关系数为0.948和0.919,物种-环境关系的累积百分率为91.1,说明排序轴与环境因子间线性结合的程度较好的反映了物种与环境间的关系。第一轴与水深(r=-0.93)、TOM含量(r=-0.57)和Chla含量(r=-0.80)负相关,与光强正相关(r=0.62);第二轴与TOM含量正相关(r=0.53)。冬季排序图前两轴的特征值为0.372和0.124,种类与环境因子排序轴的相关系数为0.992和0.956,物种-环境关系的累积百分率为70.5,说明排序轴与环境因子间线性结合的程度较好的反映了物种与环境间的关系。第一轴与水深(r=-0.96)、TOM含量(r=0.86)和Chla含量(r=0.71)负相关;第二轴与Chla含量正相关(r=0.64)。春季排序图前两轴的特征值分别为0.482和0.161,种类与环境因子的相关系数为0.957和0.949,物种-环境关系的累积百分率为83.8,说明排序轴与环境因子间线性结合的程度较好的反映了物种与环境间的关系。第一轴与水深正相关(r=0.83),与光强(r=-0.81)、Chla含量(r=-0.78)和TOM含量(r=-0.70)负相关;第二轴与水深(r=0.38)和Chla含量(r=0.36)正相关。综合而言,水深、Chla和TOM含量是影响刺参养殖池塘大型底栖动物的关键环境因子。
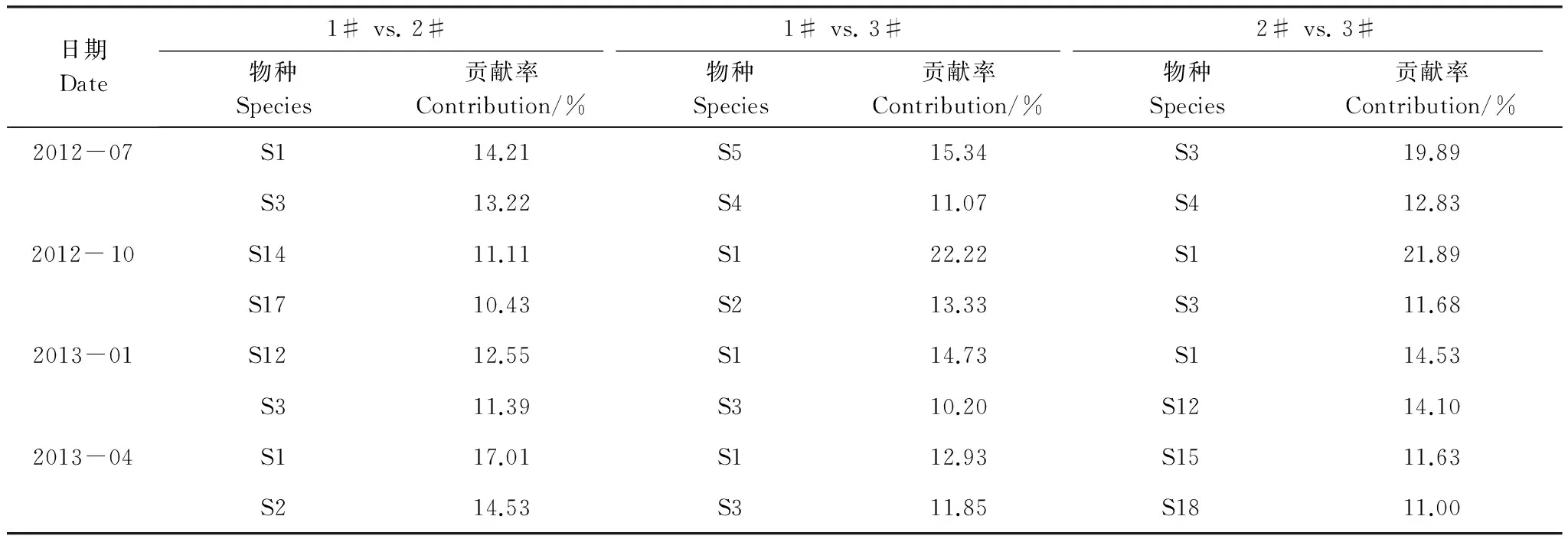
表4 刺参养殖池塘间大型底栖动物群落分歧种及贡献率
S: 物种代码见表1
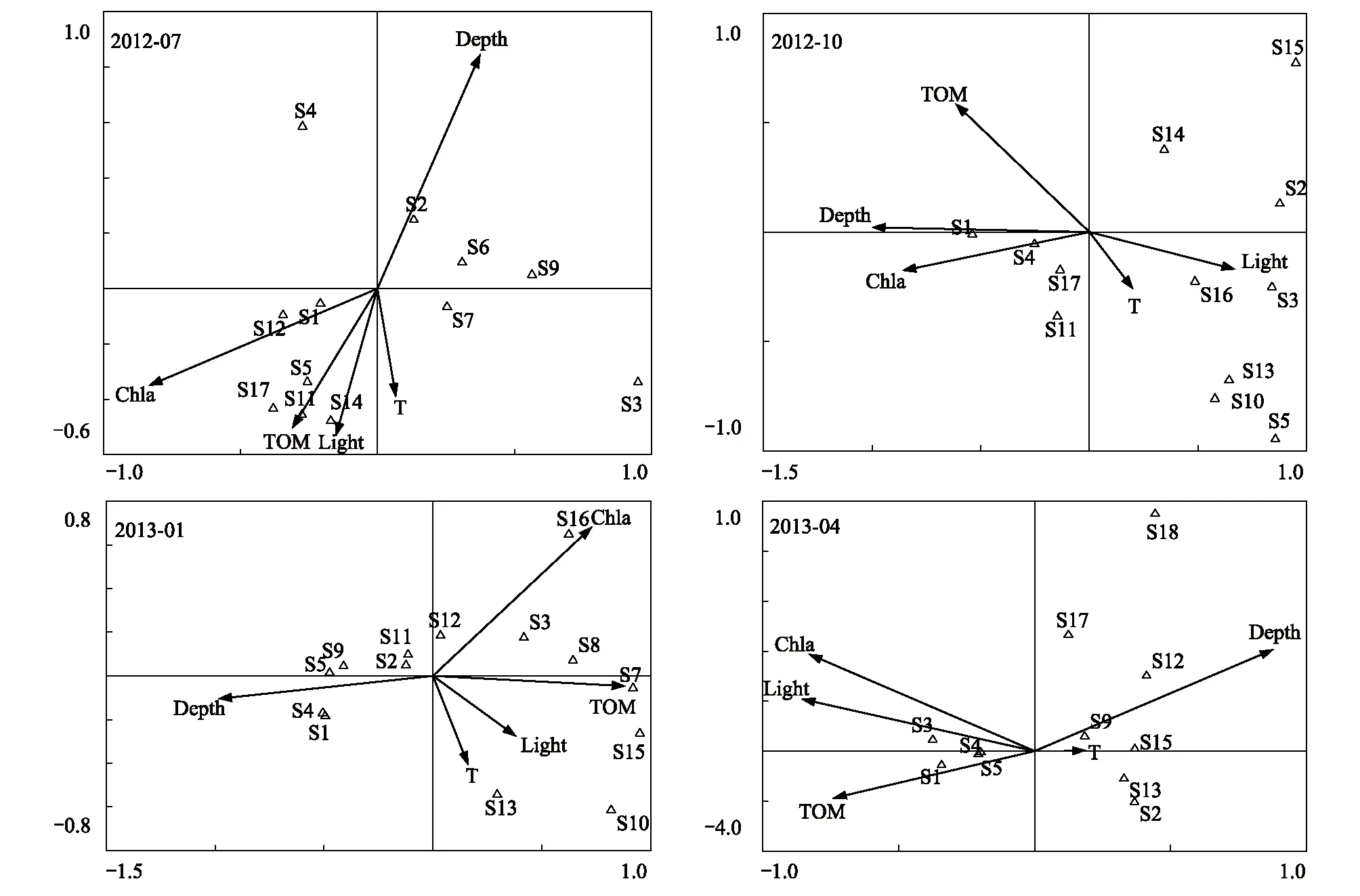
图3 大型底栖动物与环境因子CCA排序图Fig.3 CCA ordination diagram of macroinvertebrates-environmental factorsS: 物种代码见表1
3 讨论
3.1不同水深的刺参养殖池塘大型底栖动物群落结构的季节性变化
CCA分析表明,水深为影响刺参养殖池塘大型底栖动物群落的主要环境因子,这与水深是影响秋季莱州湾以及渤海中、南部大型底栖动物群落结构的重要因子一致[24-25]。各季节3个池塘间群落结构相似性表现为春季(48%)>夏季(43%)>冬季(43%)>秋季(37%)。ANOSIM分析显示,各季节3个池塘间群落结构均存在显著差异,表明水深对刺参养殖池塘大型底栖动物群落结构造成显著影响。SIMPER分析表明,各季节对3个池塘间大型底栖动物群落差异起主要作用的物种为各个池塘的优势物种。夏季3个池塘共有优势种如凸壳肌蛤(Musculussenhousia)优势度差异较大,加之各池塘特有优势种如1#池塘的圆筒原盒螺(Eocylichnabraunsi),2#池塘的须鳃虫(Cirriformiatentaculata)和3#池塘的菲律宾蛤仔(Ruditapesphilippinarum),造成各池塘间大型底栖动物群落结构相似性较低。秋季凸壳肌蛤(M.senhousia)在1#和2#池塘已不再是优势种,但在3#池塘仍保持较高优势度,同时1#和3#池塘出现新的优势种如习见织纹螺(Nassariusdealbatus)以及2#池塘独有的优势种纵肋织纹螺(Nassariusvariciferus),导致3个池塘间群落结构相似性进一步降低。冬季虽然2#池塘出现特有的优势种中华近方蟹(Hemigrapsussinensis),但其优势度指数较小,同时3#池塘凸壳肌蛤(M.senhousia)优势度变小,造成各池塘间群落结构相似性变大。春季3#池塘出现特有优势种日本关公蟹(Dorippejaponica)和日本刺沙蚕(Neanthesjaponica),但其优势度指数均较小,同时凸壳肌蛤(M.senhousia)在各池塘间差异继续变小,导致3个池塘群落结构相似性进一步变大。
3.2环境因素对刺参养殖池塘大型底栖动物多样性的影响
大型底栖动物同刺参一样,作为沉积食性动物,沉积有机物是其重要的食物来源[26],底栖微藻可为其提供更多的食物资源[27]。在不投饵的刺参养殖池塘,底部有机质主要为水层沉降的颗粒有机物主要为浮游植物和底泥内源自生底栖微藻等。水深的改变,一方面会影响由水体沉降的颗粒有机物的量;另一方面会改变池塘底部温度和光照强度,影响底泥本身内源性初级生产力,进而影响大型底栖动物群落和刺参养殖池塘环境。
CCA分析表明,叶绿素a(Chla)和总有机物(TOM)含量为影响刺参养殖池塘大型底栖动物群落的关键环境因子,且其对大型底栖动物丰度、生物量和多样性的影响是随季节变化而变化的。夏季1#池塘大型底栖动物丰度、生物量和多样性指数均高于3#池塘,这与葡萄牙大陆架较浅海域[28],以及地中海和美国南加州大陆架海域具更高的大型底栖动物丰度和多样性指数一致[29-30]。随着水深的增加,底泥中Chla的质和量都会下降,进而影响大型底栖动物群落结构导致丰度和多样性降低[21]。这可能是因为光强是影响底栖微藻生物量主要因子,而光通量主要受水体深度和浊度的影响[31],1#池塘水位较浅,底部光强大,更有利于底栖微藻的生长和繁殖,且其对底泥Chla贡献作用大于水体中浮游植物的沉降作用。1#池塘具更高底泥Chla和TOM含量,从而能够支持更多的大型底栖动物。但另一方面,池塘水深影响到底部温度和光照的调节,刺参生长适宜水温范围为10—20 ℃,水温超过20 ℃时,刺参就会发生夏眠[32]。本研究中,3个池塘间水温差异不显著,但1#池塘光强显著大于3#池塘,高水位能够降低高温和强光照对刺参的刺激。此外,夏季池塘水位过低会造成大型藻类如束生刚毛藻(Cladophorafascicularis)的发生和大量繁殖,与浮游植物发生资源竞争以及克生效应,从而抑制浮游植物的生长[33-35]。这不仅会影响刺参在底部活动,也会导致池塘初级生产力下降。综合考虑,夏季刺参养殖池塘宜保持较高水位。
秋季1#池塘大型底栖动物多样性指数高于3#池塘,但3#池塘丰度和生物量显著大于1#池塘,这与渤海湾大型底栖动物生物量[36]以及渤海中、南部大型底栖动物丰度[25]均和水深呈显著正相关一致。在水位较深水域,由于有较高的初级生产量到达底部,从而支持着较高的大型底栖动物生物量[37]。3#池塘Chla含量显著高于1#池塘,这可能是因为秋季光强减弱,底栖微藻生长受到影响,同时3个池塘底部光强差异不显著,导致内源性底栖微藻生物量差异不大;池塘底部的Chla主要由水体中浮游植物的沉降作用贡献,换水带来的颗粒有机物沉降到底部为大型底栖动物及刺参主要的食物来源。前期研究也表明秋季刺参养殖池塘参礁表面Chla主要为水体沉降的浮游植物,少部分来源于自生的底栖微藻[38]。虽然1#池塘生物多样性高,但3#池塘单细藻丰富,能够为刺参结束夏眠开始摄食提供更多的食物资源。
冬季3个池塘间大型底栖动物多样性指数差异不显著,但3#池塘丰度和生物量显著大于1#池塘,可能是由秋季3#池塘大型底栖动物高丰度和高生物量所致。1#池塘底部Chla和TOM含量显著高于3#池塘,表明池塘底部有机质主要来自内源自生底栖微藻而非水体中浮游植物。这可能是因为冬季温度低光照弱,外海中浮游植物生物量较低,导致池塘换水带来的颗粒有机物较少,而1#池塘水深较低,底部能够获得较强光照,更有利于底栖微藻生长和繁殖,能够为冬眠刺参春季摄食提供更多的食物储备。
春季同冬季情况类似,1#池塘底部Chla和TOM含量显著高于3#池塘,表明水位越低,池塘底部有机质更为丰富。这可能是由于春季光照增大,池塘越浅,底部获得的光辐射越强,更有利于底栖微藻的生长,与此同时水体中浮游植物量也逐渐增大,但其对池塘底部有机质贡献作用小于内源性底栖微藻。前期研究也发现春季刺参养殖池塘参礁表面以底栖微藻为主,且其Chla含量显著高于水体中的Chla含量[38]。丰富的有机质导致1#池塘大型底栖动物丰度和生物量大于3#池塘,但3#池塘生物多样性高,更能维持池塘生态系统的平衡。
4 结论
本研究表明,水深是影响刺参养殖池塘大型底栖动物群落结构的重要环境因子。3个不同水深池塘光照强度、叶绿素a(Chla)和总有机物(TOM)含量相差较大,导致3个池塘大型底栖动物在种类组成、丰度、生物量和多样性指数上均存在显著差异。夏季浅水位池塘有机质丰富,具更高生物多样性,但深水位池塘利于刺参夏眠;秋季深水位池塘能够为刺参开始摄食提供更多的食物资源;冬季浅水位池塘可以为刺参春季开始摄食提供更多食物储备;春季深水位池塘生物多样性较高,能够为刺参提供稳定生态环境。
[1]Herman P M J, Middelburg J J, van de Koppel J, Heip C H R. Ecology of Estuarine Macrobenthos. Advances in Ecological Research, 1999, 29: 195-240.
[2]张志南. 水层—底栖耦合生态动力学研究的某些进展. 青岛海洋大学学报: 自然科学版, 2000, 30(1): 115-122.
[3]Sun Z L, Gao Q F, Dong S L, Shin P K S, Wang F. Seasonal changes in food uptake by the sea cucumberApostichopusjaponicusin a farm pond: evidence from C and N stable isotopes. Journal of Ocean University of China, 2013, 12(1): 160-168.
[4]Stenton-Dozey J M E, Jackson L F, Busby A J. Impact of mussel culture on macrobenthic community structure in Saldanha Bay, South Africa. Marine Pollution Bulletin, 1999, 39(1/12): 357-366.
[5]Gao Q F, Cheung K L, Cheung S G, Shin P K S. Effects of nutrient enrichment derived from fish farming activities on macroinvertebrate assemblages in a subtropical region of Hong Kong. Marine Pollution Bulletin, 2005, 51(8/12): 994-1002.
[6]Da Costa K G, Nalesso R C. Effects of mussel farming on macrobenthic community structure in Southeastern Brazil. Aquaculture, 2006, 258(1/4): 655-663.
[7]Apostolaki E T, Tsagaraki T, Tsapakis M, Karakassis I. Fish farming impact on sediments and macrofauna associated with seagrass meadows in the Mediterranean. Estuarine, Coastal and Shelf Science, 2007, 75(3): 408-416.
[8]Jelbart J E, Schreider M, MacFarlane G R. An investigation of benthic sediments and macrofauna within pearl farms of Western Australia. Aquaculture, 2011, 319(3/4): 466-478.
[9]杨俊毅, 高爱根, 宁修仁, 张东声. 乐清湾大型底栖生物群落特征及其对水产养殖的响应. 生态学报, 2007, 27(1): 34-41.
[10]冯建祥, 董双林, 高勤峰, 孙侦龙, 王芳, 张凯. 海蜇养殖对池塘底泥营养盐和大型底栖动物群落结构的影响. 生态学报, 2011, 31(4): 964-971.
[11]Ellingsen K E. Soft-sediment benthic biodiversity on the continental shelf in relation to environmental variability. Marine Ecology Progress Series, 2002, 232: 15-27.
[12]Dolbeth M, Ferreira, Teixeira H, Marques J C, Dias J A, Pardal M A. Beach morphodynamic impact on a macrobenthic community along a subtidal depth gradient. Marine Ecology Progress Series, 2007, 352: 113-124.
[13]Freitas R, Ricardo F, Pereira F, Sampaio L, Carvalho S, Gaspar M, Quintino V, Rodrigues A M. Benthic habitat mapping: concerns using a combined approach (acoustic, sediment and biological data). Estuarine, Coastal and Shelf Science, 2011, 92(4): 598-606.
[14]Carvalho S, Cunha M R, Pereira F, Pousão-Ferreira P, Santos M N, Gaspar M B. The effect of depth and sediment type on the spatial distribution of shallow soft-bottom amphipods along the southern Portuguese coast. Helgoland Marine Research, 2012, 66(4): 489-501.
[15]Martins R, Sampaio L, Rodrigues A M, Quintino V. Soft-bottom Portuguese continental shelf polychaetes: Diversity and distribution. Journal of Marine Systems, 2013, 123-124: 41-54.
[16]Mutlu E, Ergev M B. Depth-related gradient of soft-bottom crustacean distribution along the Cilician shelf. Turkish Journal of Zoology, 2013, 37(3): 262-276.
[17]刘录三, 孟伟, 李新正, 李子成, 郑丙辉, 雷坤, 李中宇. 辽东湾北部海域大型底栖动物研究: Ⅱ. 生物多样性与群落结构. 环境科学研究, 2009, 22(2): 155-161.
[18]袁秀堂, 杨红生, 陈慕雁, 高菲. 刺参夏眠的研究进展. 海洋科学, 2007, 31(8): 88-90.
[19]董云伟, 董双林. 刺参对温度适应的生理生态学研究进展. 中国海洋大学学报: 自然科学版, 2009, 39(5): 908-912.
[20]张硕, 陈勇, 孙满昌. 光强对刺参行为特性和人工礁模型集参效果的影响. 中国水产科学, 2006, 13(1): 20-27.
[21]Grebmeier J M, McRoy C P, Feder H M. Pelagic-benthic coupling on the shelf of the northern Bering and Chukchi Seas. I. Food supply source and benthic biomass. Marine Ecology Progress Series, 1988, 48: 57-67.
[22]蔡永久, 姜加虎, 张路, 陈宇炜, 龚志军. 长江中下游湖群大型底栖动物群落结构及影响因素. 生态学报, 2013, 33(16): 4985-4999.
[23]陆强, 陈慧丽, 邵晓阳, 王莹莹, 陶敏, 何京, 唐龙. 杭州西溪湿地大型底栖动物群落特征及与环境因子的关系. 生态学报, 2013, 33(9): 2803-2815.
[24]周红, 华尔, 张志南. 秋季莱州湾及邻近海域大型底栖动物群落结构的研究. 中国海洋大学学报: 自然科学版, 2010, 40(8): 80-87.
[25]韩洁, 张志南, 于子山. 渤海中、南部大型底栖动物的群落结构. 生态学报, 2004, 24(3): 531-537.
[26]余婕, 刘敏, 侯立军, 许世远, 欧冬妮, 程书波. 崇明东滩大型底栖动物食源的稳定同位素示踪. 自然资源学报, 2008, 23(2): 319-326.
[27]Hansen J L S, Josefson A B. Ingestion by deposit-feeding macro-zoobenthos in the aphotic zone does not affect the pool of live pelagic diatoms in the sediment. Journal of Experimental Marine Biology and Ecology, 2004, 308(1): 59-84.
[28]Martins R, Quintino V, Rodrigues A M. Diversity and spatial distribution patterns of the soft-bottom macrofauna communities on the Portuguese continental shelf. Journal of Sea Research, 2013, 83: 173-186.
[29]Coll M, Piroddi C, Steenbeek J, Kaschner K, Ben Rais Lasram F, Aguzzi J, Ballesteros E, Bianchi C N, Corbera J, Dailianis T, Danovaro R, Estrada M, Froglia C, Galil B S, Gasol J M, Gertwagen R, Gil J, Guilhaumon F, Kesner-Reyes K, Kitsos M S, Koukouras A, Lampadariou N, Laxamana E, López-Fé de la Cuadra C M, Lotze H K, Martin D, Mouillot D, Oro D, Raicevich S, Rius-Barile J, Saiz-Salinas J I, Vicente C S, Somot S, Templado J, Turon X, Vafidis D, Villanueva R, Voultsiadou E. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS One, 2010, 5(8): e11842.
[30]Bergen M, Weisberg S B, Smith R W, Cadien D B, Dalkey A, Montagne D E, Stull J K, Velarde R G, Ranasinghe J A. Relationship between depth, sediment, latitude, and the structure of benthic infaunal assemblages on the mainland shelf of southern California. Marine Biology, 2001, 138(3): 637-647.
[31]MacIntyre H L, Cullen J J. Primary production by suspended and benthic microalgae in a turbid estuary: time-scales of variability in San Antonio Bay, Texas. Marine Ecology Progress Series, 1996, 145: 245-268.
[32]陈勇, 高峰, 刘国山, 邵丽萍, 石国锋. 温度、盐度和光照周期对刺参生长及行为的影响. 水产学报, 2007, 31(5): 687-691.
[33]Jin Q, Dong S L. Comparative studies on the allelopathic effects of two different strains ofUlvapertusaonHeterosigmaakashiwoandAlexandriumtamarense. Journal of Experimental Marine Biology and Ecology, 2003, 293(1): 41-55.
[34]许妍, 董双林, 于晓明. 缘管浒苔对赤潮异弯藻的克生效应. 生态学报, 2005, 25(10): 2681-2685.
[35]张善东, 俞志明, 宋秀贤, 宋飞, 王悠. 大型海藻龙须菜与东海原甲藻间的营养竞争. 生态学报, 2005, 25(10): 2676-2680.
[36]蔡文倩, 孟伟, 刘录三, 朱延忠, 周娟. 春季渤海湾大型底栖动物群落结构特征研究. 环境科学学报, 2013, 33(5): 1458-1466.
[37]韩洁, 张志南, 于子山. 渤海大型底栖动物丰度和生物量的研究. 中国海洋大学学报: 自然科学版, 2001, 31(6): 889-896.
[38]任贻超, 董双林, 王修滨, 王芳, 田相利, 高勤峰. 刺参养殖池塘一种塑料人工参礁表面生态学特征初步研究. 海洋科学, 2013, 37(1): 29-34.
Seasonal changes in macrobenthic community structures of sea cucumberApostichopusjaponicus(Selenka) farming ponds at different water depths
WEN Bin1,2, GAO Qinfeng1,2,*, DONG Shuanglin1,2, NING Luguang3
1KeyLaboratoryofMariculture,OceanUniversityofChina,Qingdao266003,China2FunctionLaboratoryforMarineFisheriesScienceandFoodProductionProcesses,QingdaoNationalLaboratoryforMarineScienceandTechnology,Qingdao266235,China3DongyingHaiyueAquacultureCo.Limited,Dongying257500,China
Sediment samples were collected from three sea cucumber (Apostichopusjaponicus) farming ponds of varying water depths, i.e., low (1#), normal (2#), and high (3#) water level depths, in the mariculture zone of Jinghai Bay from July 2012 to April 2013. Macrobenthic community structures were analyzed to investigate the effects of the changes in water depth. Light intensity, Chlorophyll a (Chla) and total organic matter (TOM) contents differed significantly among the three ponds. Light intensity and TOM content of pond 1# were significantly higher than those of pond 3# in summer, winter, and spring; whereas no significant differences were observed among the three ponds in autumn. Chla content of pond 1# was significantly higher than that of pond 3# in summer, winter, and spring; but significantly lower than that of pond 3# in autumn. No significant differences in water temperature were observed among the three ponds for any of the four seasons. Throughout the investigation, 41 macrobenthic species were identified, of which 21 species were annelids (51.2%), 16 species were mollusks (39.0%) and four species were crustaceans (9.8%). Significant differences in species composition, dominant species, abundance, biomass, and diversity index were observed among the three ponds during each season. Macrobenthic abundance and biomass of pond 1# were significantly higher than those of pond 3# in summer, but significantly lower than those of pond 3# in autumn and winter. In spring, macrobenthic abundance of pond 1# was significantly higher than that of pond 3#, with no significant difference in biomass between the two ponds. These results could be attributed to the remarkable fluctuations in the abundance of the dominant species such asMusculussenhousia,Lumbriconerisheteropoda,Cirriformiatentaculata,RuditapesphilippinarumandEocylichnabraunsi. The Margalef′s richness index of pond 1# was significantly higher than that of pond 3# in summer; and the Shannon-Wiener diversity and Pielou′s evenness indices of pond 1# were significantly higher than those of pond 3# in autumn. The Margalef′s richness index and Pielou′s evenness index of pond 3# however, were significantly higher than those of pond 1# in spring. No significant differences in diversity index were observed among the three ponds in winter. Hierarchical cluster analysis based on the Bray-Curtis similarity coefficients revealed that the community structure similarities among the three ponds were 43% in July, 37% in October, 43% in January and 48% in April. The low similarities among the three ponds suggested that the macrobenthic community structures were significantly influenced by differences in water depth. The analysis of similarities (ANOSIM) indicated significant differences in species compositions among the three ponds for each season. The similarity percentages analysis (SIMPER) showed that the species that comprised the macrobenthic communities of different ponds, were also the dominant species in each pond for each season. Canonical correspondence ordination analysis (CCA) showed that water depth, Chla, and TOM contents were the key factors influencing the macrobenthic community structures in summer and winter; whereas water depth, light intensity, Chla, and TOM contents were the key factors affecting macrobenthic community structures in spring and autumn.
water depth;Apostichopusjaponicus; macrobenthic community; biodiversity
国家自然科学基金项目(31172426,31372549);国家科技支撑计划项目(2011BAD13B03)
2015-06-29;
2015-10-13
Corresponding author.E-mail: qfgao@ouc.edu.cn
10.5846/stxb201506291335
温彬, 高勤峰, 董双林, 宁鲁光.不同水深条件下刺参养殖池塘大型底栖动物群落结构季节性变化特征.生态学报,2016,36(14):4327-4336.
Wen B, Gao Q F, Dong S L, Ning L G.Seasonal changes in macrobenthic community structures of sea cucumberApostichopusjaponicus(Selenka) farming ponds at different water depths.Acta Ecologica Sinica,2016,36(14):4327-4336.
猜你喜欢
河北水利(2022年10期)2022-12-29
海洋通报(2022年4期)2022-10-10
海洋信息技术与应用(2022年1期)2022-06-05
红蜻蜓·低年级(2021年12期)2022-01-19
红蜻蜓·低年级(2021年12期)2021-12-19
今日农业(2020年19期)2020-12-14
大连海洋大学学报(2020年2期)2020-05-06
中国科技信息(2015年2期)2015-11-16
浙江国土资源(2015年4期)2015-05-09
植物营养与肥料学报(2014年1期)2014-03-11
